VEGF及其受体基因多态性与有氧运动能力表型的关联研究
2010-09-14何子红
聂 晶,胡 扬,席 翼,文 立,何子红
VEGF及其受体基因多态性与有氧运动能力表型的关联研究
聂 晶1,胡 扬2,席 翼3,文 立3,何子红4
目的:探讨血管内皮生长因子(VEGF)及其受体基因多态性与有氧运动能力表型的关联。方法:102名中国北方汉族男子进行18周长跑训练,测试训练前后的O2max、12 km/h跑速下的RE和心室结构功能指标。用PCR-RFLP分析VEGF基因SNP/C-2578A、VEGFR1基因SNP/A+193019G以及VEGFR2基因SNP/A+18487T。结果:3个多态位点与O2max、RE的初始值及训练敏感性均不关联,但与部分心室结构和/或功能指标的初始值及训练敏感性有关联。其中SNP/C-2578A的携A等位基因者、SNP/A+193019G的AA基因型者和SNP/A+18487T的AT、AA基因型者均表现出较好的适应性变化。结论: VEGF基因SNP/C-2578A、VEGFR1基因SNP/A+193019G以及VEGFR2基因SNP/A+ 18487T可以作为预测18周长跑训练后部分心室结构和/或功能指标敏感性差异的分子遗传学标记。但还需加大样本量进一步验证。
血管内皮生长因子;受体;基因多态性;最大耗氧量;跑节省化;心功能;有氧运动
遗传学已证实,有氧运动能力受多个基因与环境的共同作用,为多因子的复杂表型,且尚未发现有主效基因存在。近几年来,由SNP研究委员会与美国国立人类基因组研究院发起的对单核苷酸多态性(single nucleotial polymorphisms,SNPS)的大规模研究表明,单核苷酸变异能够为解释个体间的表型差异提供新方法。因此,利用人类基因组大量的SNP信息来探究运动能力与训练敏感性的遗传机制已成为国内外体育科学界的研究热点。
血管内皮生长因子(VEGF),又称血管通透性因子(VPN),是一种特异作用于血管内皮细胞的强有力的多功能细胞因子。研究表明,组织缺氧时,细胞通过分泌VEGF与血管内皮细胞表面的VEGF受体(VEGFR1, VEGFR2)结合,启动一系列细胞内信号传递途径使血管生成增加[14]。这在生理性和病理性的血管发生和生长中具有重要作用。毛细血管增生是机体组织对有氧运动训练的一种关键适应性变化。这种变化可以缩短氧气从毛细血管向组织弥散的距离、扩大弥散面积、延长弥散时间,从而增强氧的传递与利用,是提高有氧运动能力的重要机制之一。现已有不少关于急性有氧运动和长期耐力训练影响VEGF及其受体表达的报道[8-10,12,15,24],结果提示VEGF及其受体可能是与氧运动能力相关的重要候选基因。
因此,本研究拟通过VEGF及其受体基因多态性与有氧运动训练前生理指标的初始值和训练敏感性的相关分析,来揭示个体间有氧运动能力的不同生理表型,以期为制定个体化的训练方案提供新的指标和依据。
1 研究对象和方法
1.1 研究对象
102名汉族受试者来自中国武装警察8631部队的新兵,均为2003年1月入伍,3月参加本实验,籍贯为东北三省、河北、山东等淮河以北平原地区。所有士兵入伍前均无训练史,也无由于生活环境而导致的长期耐力性活动,且无家族疾病史。受试者平均年龄为18.82±0.88岁,身高为171.67±5.83 cm,体重为60.27±6.54 kg。
1.2 长跑训练计划
训练时间共18周,其中,前2周为适应性训练。第3周开始进行每周3次5 000 m匀速跑,分别在每周1、3、5下午4:00~5:00进行。训练强度为95%个体通气VT时的HR(HRVT)±3次/min。从第10周开始以105%HRVT±3次/min强度运动。心率的控制由芬兰产的Polar心率表实施。如有气候和军事行动等不可抗拒的原因而停训,择日补训。参加训练的士兵组作息时间一致,饮食条件一致,训练安排一致。同时,参加训练的士兵不再进行额外的2 000 m以上的耐力训练,只进行一般军事和力量训练。
1.3 生理学指标的测试方法
1.3.1 身高、体重
训练前后各测定一次,两次条件一致,均参照国民体质测量标准的要求实施,由同一实验师完成。
1.3.2 最大耗氧量和跑节省化
1.先让所有受试者依次在跑台上试跑3 min,以适应跑台的运动方式。
2.第1 min:坡度为0%,速度逐渐增加到7.5 km/h,受试者从自然走步移行为跑步,保持2 min。
3.第3 min:坡度为0%,速度增至10.0 km/h,保持2 min。
4.第5 min:坡度为0%,速度增至12.0 km/h(“适宜的极限下负荷速度”),保持5 min,其间测定RE。
5.第10 min:坡度增加2%,速度增至12.5 km/h。6.第11 min后:坡度每分钟增加2%,速度每分钟增加0.5 km/h,视个人能力逐级递增,直至力竭。
1.3.3 心室结构和功能
采用Doppler超声心动图仪分别在训练的18周前后各测定1次。每次测试前先静坐10 min后平卧,测定安静时心室结构和功能指标,然后以起始负荷50 W和60 rpm的频率开始蹬功率自行车,每3 min增加负荷50 W,直至150 W蹬满3 min后停止。平卧恢复3 min。分别测定50 W、100 W、150 W负荷的最后30 s和恢复的最后30 s的左心室结构和功能指标。每名受试者两次测试中的时相(上午、下午、晚上)保持一致。全部测试由同一名实验师完成。
测试指标包括直接测试指标和衍生指标,其中直接测试的指标有左室舒张末内径(EDD)、左室收缩末内径(ESD)、左室舒张末后壁厚度(PWD)、左室收缩末后壁厚度(PWS)、室间隔舒张末厚度(IVSD)和室间隔收缩末厚度(IVSS);衍生的指标有每搏输出量(SV)、每搏输出量指数(SI)、心输出量(CO)、心输出量指数(CI)、射血分数(EF)、左室重量(LVM)、左室重量指数(LVM I)和心动周期(T)。计算公式如下:SV(m l)=EDD3-ESD3;CO(L/min)=SV ×HR;EF=(EDD3-ESD3)/EDD3×100%;LVM(g)= 1.04×[(EDD+PWD+IVSD)3-EDD3]-14;T=1/HR;体表面积(BSA)=0.006×身高(cm)+0.0128×体重(kg)-0.1529;CI=CO/BSA;SI=SV/BSA;LVM I=LVM/BSA。
1.4 基因多态性分析
用Promaga公司试剂盒提取全血DNA,并在紫外分光光度计上测定OD260、OD 280。若OD260/OD280>1.7,说明DNA纯度合格。人类VEGF基因位于染色体6p 21.3,由8个外显子和7个内含子组成。VEGFR1、VEGFR2基因分别位于染色体13q12、4q12,由30个外显子和29个内含子组成。本研究分别在这3个基因上选取SNP/ C-2578A、SNP/A+193019G、SNP/A+18487T,应用primer 5.0自行设计引物,采用PCR-RFLP方法进行基因分型(表1)。

表1 本研究引物序列和基因多态性分析条件一览表Table 1 Primersand PCR-RFLPConditions for these Genes Polymorphisms
1.5 统计学分析
所有数据处理采用SPSS 13.0软件统计包完成。采用卡方检验和精确率检验受试者基因型频率是否符合H-W平衡定律;训练前后生理指标数据先用K-S检验是否符合正态分布。若符合正态分布,则基因型之间各生理指标的初始值指标采用独立样本t检验或单因素方差分析,基因型之间训练敏感性采用协方差处理。若不符合正态分布采用非参数检验处理。显著性水平设为P<0.05,非常显著性水平设为P<0.01。变化量(△)=训练后-训练前。
2 结果
2.1 VEGF及其受体基因多态性的分布特征
受试者VEGF基因SNP/C-2578A、VEGFR1基因SNP/A+193019G以及VEGFR2基因SNP/A+18487T基因型的分布结果见表2。经卡方检验和精确率检验后,各基因型频率均符合H-W平衡,所以,具有群体代表性。由于SNP/C-2578A的AA基因型和SNP/A+193019G的GG基因型人数较少,所以,在以下分析中将其与杂合型进行合并。

表2 本研究VEGF及其受体基因多态性基因型频率一览表Table 2 Genotypic frequencies of VEGF and its receptors genes polymorphisms
长跑训练前,SNP/C-2578A、SNP/A+193019G以及SNP/A+18487T各基因型之间的O2max绝对值(L/ min)和相对值(m l/kg/min)的初始值均没有显著差异;长跑训练后,不同基因型O2max(L/min、ml/kg/min)的变化也无显著差异(表3)。
2.3 VEGF及其受体基因多态性与RE的关联性
长跑训练前,SNP/C-2578A、SNP/A+193019G以及SNP/A+18487T各基因型之间跑节省化(12 km/h)的HR、VE、O2(L/min,ml/kg/min)的初始值均没有显著差异;经过18周训练后,所有受试者的跑节省化能力均增强,但是,不同基因型间的变化没有显著差异(表4)。
表3 本研究各基因型O2max的变化一览表Table 3 Changs ofO2max in diffrenet Genotypes

表3 本研究各基因型O2max的变化一览表Table 3 Changs ofO2max in diffrenet Genotypes
O2max (L/min) rO2max (m l/kg/min)△O2max (L/min)△rO2max (m l/kg/min) C-2578A CC CA+AA 3.42±0.41 3.47±0.34 56.84±4.21 57.79±4.03 A+193019G AA AG+GG 3.44±0.40 3.44±0.36 0.06±0.25 0.03±0.23 -0.29±3.91 -0.49±3.88 57.47±4.57 56.87±3.49 A+18487T AA A T TT 3.41±0.28 3.42±0.40 3.49±0.42 0.06±0.23 0.03±0.25 -0.51±3.95 -0.17±3.83 56.56±3.18 57.58±4.46 57.06±4.21 0.08±0.23 0.02±0.24 0.07±0.23 0.25±4.20 -1.01±3.87 0.13±3.70
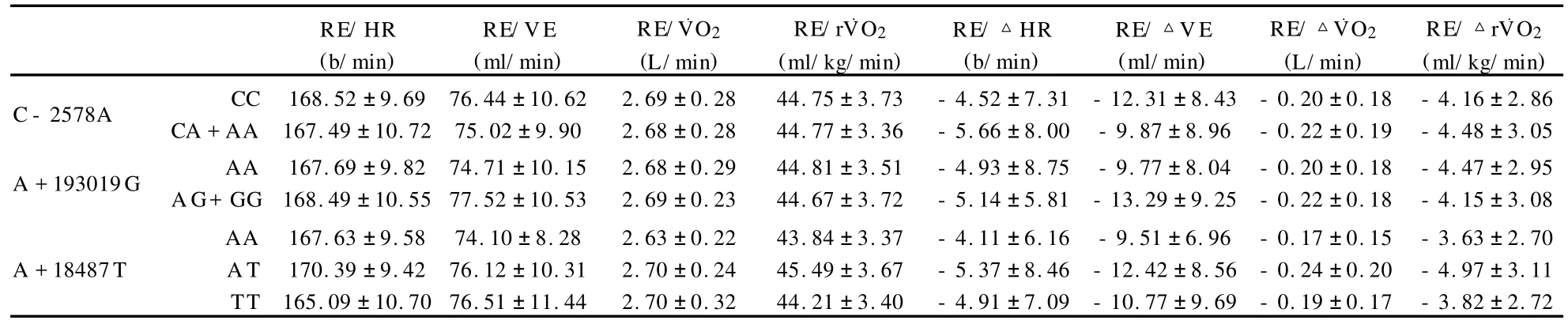
表4 本研究各基因型RE的变化一览表Table 4 Changs of RE in diffrenet Genotypes
2.4 VEGF及其受体基因多态性与心室结构的关联性
长跑训练前,SNP/C-2578A、SNP/A+193019G以及SNP/A+18487T各基因型之间心室结构指标的初始值均没有显著差异。经过18周训练后,SNP/C-2578A的CC基因型者ESD增加量明显高于携A等位基因者; SNP/A+193019G的AA基因型者ESD、IVSS、PWD、LVM以及LVM I的增加量均明显高于携G等位基因者;SNP/A +18487T的AT和AA基因型者EDD的增加量均显著高于TT基因型者,且AT基因型者PWD较其他基因型者显著下降。不同基因型者其他指标的变化量未见显著差异(表5)。

表5 本研究各基因型心室结构的变化一览表Table 5 Changs of Left Ventricular Structure in diffrenet Genotypes
2.5 VEGF及其受体基因多态性与心功能的关联性
2.5.1 VEGF及其受体基因多态性与安静状态下心功能的关联性
长跑训练前,SNP/C-2578A、SNP/A+193019G以及SNP/A+18487T各基因型之间心功能指标的初始值均没有显著差异。经过18周训练后,SNP/C-2578A的CC基因型者T增加量明显低于携A等位基因者;SNP/A +18487T的AA、AT基因型者SV和EF的增加量均明显高于TT基因型者。而其他指标的变化量在各基因型间未出现显著差异(表6)。
2.5.2 VEGF及其受体基因多态性与50 W负荷下心功能的关联性
长跑训练前,SNP/C-2578A、SNP/A+193019G以及SNP/A+18487T各基因型50W负荷下心功能指标的初始值均没有显著差异;经过18周训练后,SNP/A+ 193019G的AA基因型者SV和T的增加量明显高于携G等位基因者。而其他指标的变化量在各基因型间未见显著性差异(表7)。
2.5.3 VEGF及其受体基因多态性与100 W负荷下心功能的关联性
长跑训练前,SNP/C-2578A的CC基因型者CO和CI的初始值均高于携A等位基因者,而其余各指标的初始值在不同基因型间未见显著差异。经18周训练后, SNP/C-2578A的CC基因型者EF较携A等位基因者显著下降。其他指标的变化量在SNP/C-2578A、SNP/A+ 193019G以及SNP/A+18487T各基因型间未见显著差异(表8)。
2.5.4 VEGF及其受体基因多态性与150 W负荷下心功能的关联性
长跑训练前,SNP/C-2578A、SNP/A+193019G以及SNP/A+18487T各基因型之间心功能指标的初始值均没有显著差异;经过18周训练后,SNP/C-2578A的CC基因型者EF的变化量较携A等位基因者明显降低,其余各指标的变化量在不同基因型间未出现显著差异(表9)。
2.5.5 VEGF及其受体基因多态性与恢复期心功能的关联性
长跑训练前,SNP/C-2578A、SNP/A+193019G以及SNP/A+18487T各基因型之间心功能指标的初始值均没有显著差异;经过18周训练后,SNP/C-2578A的CC基因型者T增加量明显低于携A等位基因者,而其他各指标的变化量在不同基因型间未见显著差异(表10)。

表6 本研究VEGF及其受体基因多态性各基因型安静状态下心功能的变化一览表Table 6 Changs of Left Ventricular Function Under Static Status in Diffrenet Genotypes
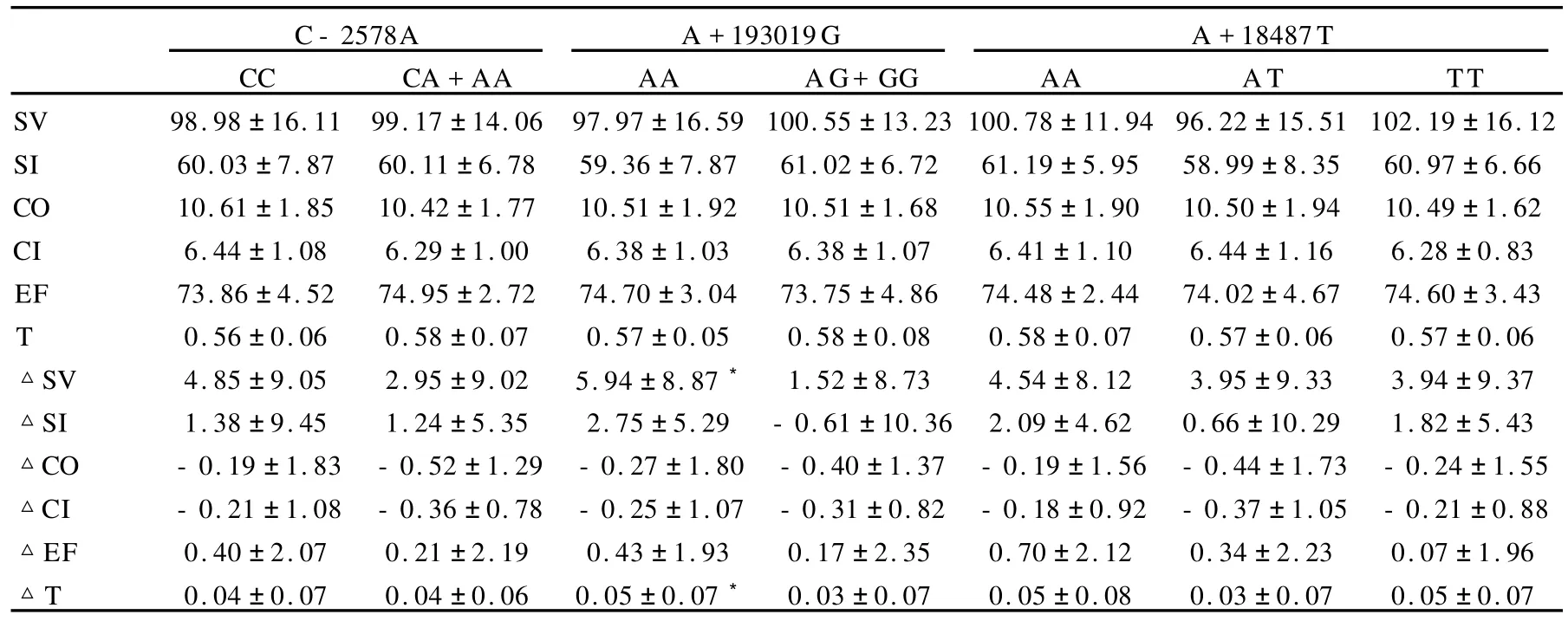
表7 本研究VEGF及其受体基因多态性各基因型50 W负荷下心功能的变化一览表Table 7 Changs of Left Ventricular Function Under 50 W Load Exercise in Diffrenet Genotypes

表8 本研究VEGF及其受体基因多态性各基因型100 W负荷下心功能的变化一览表Table 8 Changs of Left Ventricular Function Under 100 W Load Exercise in Diffrenet Genotypes
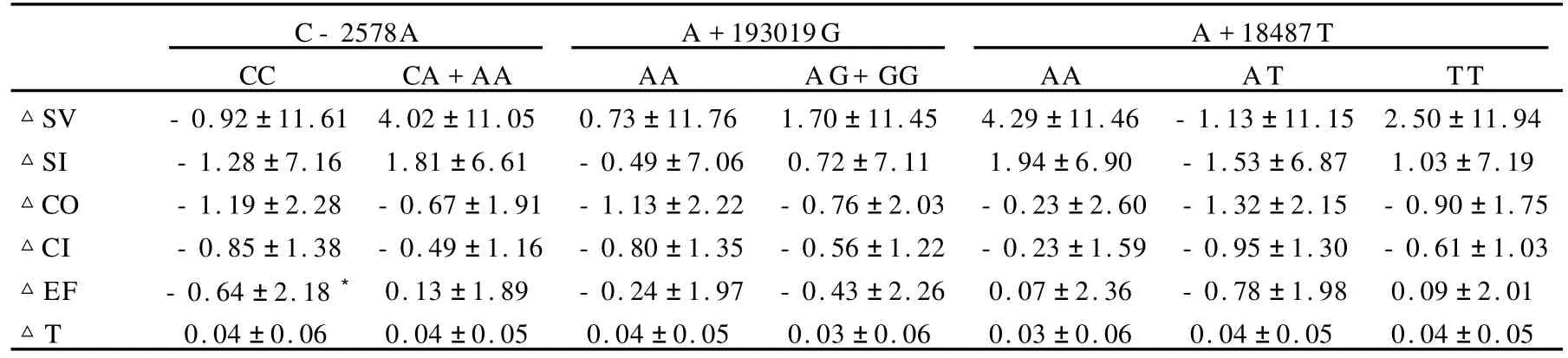
续表8

表9 VEGF及其受体基因多态性各基因型150 W负荷下心功能的变化一览表Table 9 Changs of Left Ventricular Function Under 150W Load Exercise in Diffrenet Genotypes
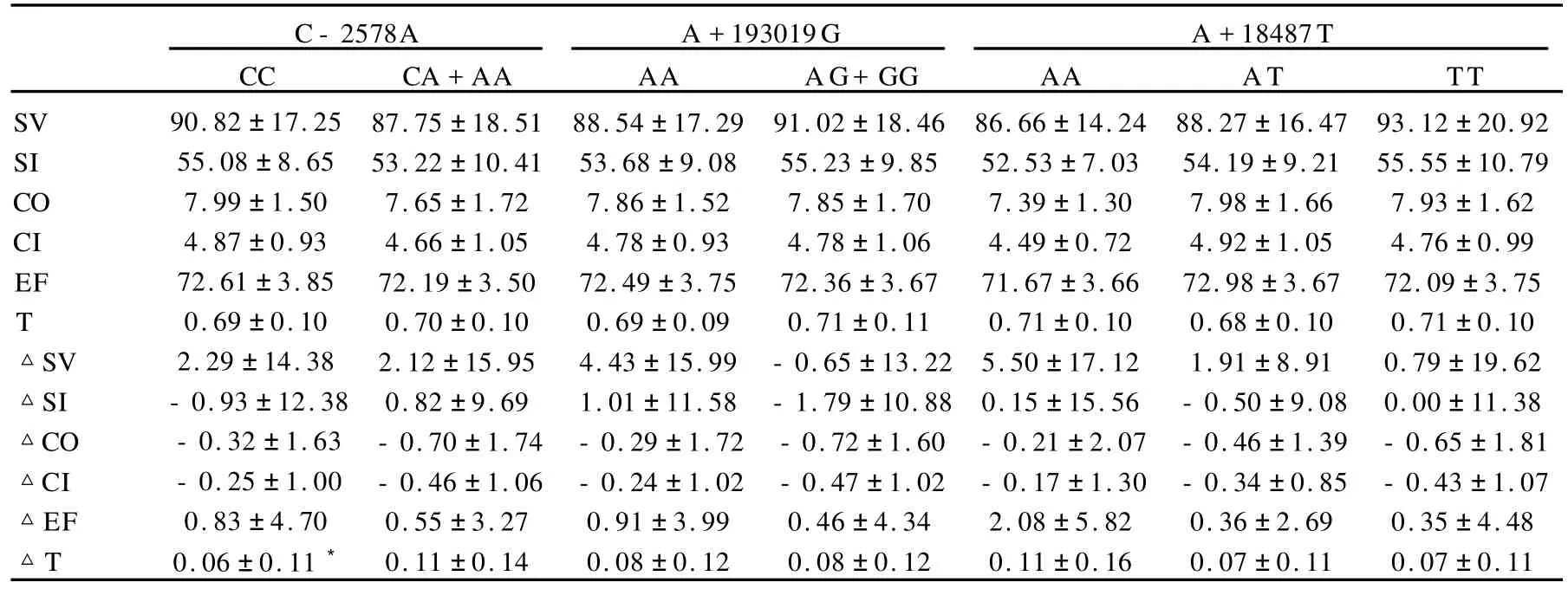
表10 VEGF及其受体基因多态性各基因型恢复期心功能的变化一览表Table 10 Changs of Left Ventricular Function on Recovery Process in Diffrenet Genotypes
3 讨论
VO2max和RE都是反映人体有氧运动能力的指标,其中,O2max代表机体整体利用氧的最大能力,并能综合评价呼吸和循环机能水平。同时,O2max与心血管疾病的死亡率有关[6]。而RE是评价次最大负荷跑速下氧利用能力的指标,通常通过测试次最大强度下的稳态的耗氧量来表示。有研究表明,在O2max相似的情况下,RE能更好的预测耐力成绩[21,26]。长期的耐力训练能增加骨骼肌和心肌毛细血管密度,提高机体运氧和耗氧能力,从而影响O2max和RE。VEGF及其受体在正常人体中广泛分布,其表达在许多组织中可检测到,包括骨骼肌和心肌组织。研究证实,VEGF及其受体确实参与了有氧运动的血管增生[11,12,19],且Timmon等学者发现,6周耐力训练后O2max训练敏感组和不敏感组中,VEGF及其受体mRNA表达存在差异[24]。这些差异提示可能与基因多态性有关。
但本研究结果显示,VEGF基因SNP/C-2578A、VEGFR1基因SNP/A+193019G以及VEGFR2基因SNP/A+ 18487T与O2max、RE的初始值和训练敏感性均不关联。这与Prior等学者的研究结果不一致。目前关于VEGF及其受体基因多态性与有氧运动能力表型的研究仅发现两篇。Prior等学者在2006年首次报道VEGF基因启动子上SNP/C-2578A,SNP/G-1154A和SNP/C-634G(即SNP/C +405G)单体型与VEGF的基因表达和有氧运动能力表型(O2max)有关联[18]。在146名老年黑种人和白种人中,携AAG和CGC单体型者VEGF基因表达显著高于携AGG和CGG单体型者;携AAG和/或CGC单体型者在有氧运动前O2max的初始值均明显高于携AGG和/或CGG单体型者,且O2max的训练敏感性也有类似的趋势(P=0.06)。
Ahmetov等学者通过case-control研究发现,VEGFR2基因His472Gln多态性与有氧运动能力有关联[3]。携472Gln等位基因频率在男性耐力运动员中显著高于男性对照组。其中,Gln/Gln基因型在23名杰出有氧耐力运动员中分布频率显著高于其他水平耐力运动员及对照组。并进一步通过genotype-phenotype关联研究发现,VEGFR2基因His472Gln多态性与有氧运动能力表型有关联。在26名杰出男运动员中,携472Gln等位基因者O2max(L/ min)显著高于His/His基因型者;在12名较杰出女运动员中,携472Gln等位基因者O2max(ml/min/kg)也显著高于His/His基因型者。另外,肌肉活检发现,携472Gln等位基因者慢肌百分比含量在运动员及对照组中均显著高于His/His基因型者。
因此,推测以上结果的不同可能与种族、年龄、样本量、训练方案、统计方法等多种因素有关。且本研究是进行单个位点与表型的相关分析,而Prior等学者进行的单体型研究包含了多个SNPs的遗传信息。单体型信息是包括相关基因定位的大规模分子遗传学的重要组成部分[13]。许多研究表明,在与复杂性状的相关分析中,采用单体型比单个SNP具有更好的统计分析效果[23]。同时,已有研究发现,位于6q21染色体区域存在与O2max关联的遗传标记[20]。笔者认为,在今后的研究中可进一步加大样本量,进行多位点的单体型分析,从而继续寻求与有氧运动能力表型(O2max和RE)关联的功能性位点。
3.2 VEGF及其受体基因多态性与左心室结构功能的关联分析
心血管功能的改善是有氧运动最显著的效益,而此效益可能与心肌组织学的改变有关。研究发现,耐力训练后,左室心肌组织中毛细血管数密度显著增加,且表现在毛细血管的绝对数量和相对数量均增加[2]。这是机体对训练的一种适应性变化。心肌是对氧特别敏感的组织,也是耗氧量最高的组织。当机体进行中等以上强度运动时,心脏做功加强引起耗氧量增加,因而对氧需求也相应增加,但是冠状动脉不能满足心肌代谢需要,进而刺激毛细血管以及侧支循环的增加以代偿组织的需要。这样,心肌血液供应增多,有利于心脏的氧化代谢和能量的产生。Amaral等发现,大鼠经过8周的耐力训练后心肌VEGF、VEGFR1、VEGFR2的mRNA及蛋白表达均显著上调,且心肌毛细血管密度明显增加[4]。国内也有学者研究发现, 3~5周的中等负荷运动,心肌毛细血管分布增加,VEGF基因表达也有所增加[1]。这些结果提示,耐力训练会增强心肌VEGF血管生成的信号途径。而心脏主要通过冠状血管扩张、红细胞增多和毛细血管增生来增加心肌细胞氧的供应,有利于心脏的射血,从而增强有氧运动能力。因此,VEGF及其受体的生物学活性与运动心脏的形成密切相关。
本研究结果显示:1)VEGF基因SNP/C-2578A的CC基因型者ESD训练敏感性明显高于携A等位基因者;安静状态下以及训练后恢复期携A等位基因者T训练敏感性明显高于CC基因型者;在100 W运动负荷下,CC基因型者CO和CI的初始值均高于携A等位基因者,而携A等位基因者EF训练敏感性显著高于CC基因型者;在150 W运动负荷下携A等位基因者EF训练敏感性也明显高于CC基因型者。2)VEGFR1基因SNP/A+193019G的AA基因型者ESD、IVSS、PWD、LVM以及LVM I的训练敏感性均明显高于携G等位基因者;在50 W运动负荷下AA基因型者SV和T的训练敏感性也明显高于携G等位基因者。3)VEGFR2基因SNP/A+18487T的AT和AA基因型者EDD的训练敏感性均显著高于TT基因型者,且AT基因型者PWD较其他基因型者显著下降;安静状态下AA、AT基因型者SV和EF的训练敏感性亦均明显高于TT基因型者。
以上结果表明,VEGF基因SNP/C-2578A与100 W运动负荷下CO和CI的初始值有关联,且该多态性以及VEGFR1基因SNP/A+193019G、VEGFR2基因SNP/A+ 18487T均与部分心室结构功能指标的训练敏感性有关联。其中SNP/C-2578A的携A等位基因者、SNP/A+ 193019G的AA基因型者和SNP/A+18487T的AT、AA基因型者均表现出较好的适应性变化。其关联机制可能与这些多态位点在基因中所处位置以及蛋白功能有关。SNP/C-2578A位于VEGF基因的启动子,该区域虽与基因所编码的氨基酸序列无关,但在DNA→蛋白质合成过程中发挥重要的调控作用。Shahbazi等研究发现,30名健康人VEGF基因SNP/C-2578A和VEGF表达量有关联, CC纯合子VEGF表达量明显高于AA纯合子,CA杂合子VEGF表达量居中[22]。这可能是由于启动子上碱基的改变影响了VEGF基因5’侧翼序列的一些反应元件对VEGF转录的调节。例如,VEGF基因启动子上-2012到-2005区存在低氧反应元件(HRE),HRE上有与HIF-1结合的位点,且HRE受到上游激活蛋白1以及下游激活蛋白2α的相互作用影响[17]。因此,推测该碱基的改变影响了转录因子结合位点,进而影响转录活性以及表达生成。关于SNP/A+193019G的研究目前尚未见报道,推测碱基的改变影响了mRNA的稳定性和蛋白的生成量。SNP/ A+18487T位于VEGFR2基因第11外显子上,编码胞外第5个免疫球蛋白样结构域,该位点碱基的改变引起了相应密码子的改变,由谷氨酰胺转变为组氨酸(Gln→His),进而影响其蛋白质结构,改变了VEGFR2的活性。早有研究报道,第4到第7个免疫球蛋白结构域包含了与R2激活有关的结构特征[7,16]。有研究表明,VEGFR2基因SNP/ A+18487T具有功能性,与一些病理表型有关联,且该多态还会影响R2与VEGF的结合活性[5,25,27],且已有学者发现,在23名杰出有氧耐力运动员中Gln/Gln(即AA)基因型分布频率显著高于其他水平耐力运动员及对照组;携472Gln(即A)等位基因者O2max(L/min)和慢肌百分比含量均显著高于His/His(即TT)基因型者[3],提示携A等位基因者有氧运动能力更强,这与本结果较相似。但由于样本量大小不同,且本研究采用的是共显性基因模型,因而导致了统计方法上的差异。
4 结论
VEGF基因SNP/C-2578A、VEGFR1基因SNP/A+ 193019G以及VEGFR2基因SNP/A+18487T与O2max、RE的初始值、训练敏感性均不关联。但与部分心室结构和/或功能指标的初始值及训练敏感性有关联,其中,SNP/C-2578A的携A等位基因者、SNP/A+ 193019G的AA基因型者和SNP/A+18487T的AT、AA基因型者均表现出较好的适应性变化,可以作为预测心室结构和/或功能指标训练敏感性差异的遗传学标记。但由于SNP/C-2578A的AA基因型和SNP/A+193019G的GG基因型人数较少,还需加大样本量进一步验证。
[1]步斌.不同负荷运动对大鼠心肌促血管内皮生长因子表达的影响[J].中国运动医学杂志,2005,24(2):165-169.
[2]田野.运动生理学高级教程[M].北京:高等教育出版社,2003:234.
[3]AHM ETOV I I,HA KIMULL INA A M,POPOV D V,et al.Association of the VEGFR2 gene His472Gln polymorphism with endurance-related phenotypes[J].Eur J Appl Physiol,2009,107 (1):95-103.
[4]AMARAL SL,PAPANEK P E,GREENE A S.AngiotensinⅡand VEGF are involved in angiogenesis induced by sho rt-term exercise training[J].Am J Physiol Heart Circ Physiol,2001,281 (3):H1163-1169.
[5]ELL IS SG,CHEN M S,JIA G,et al.Relation of polymorphisms in five genes to long-term ao rtocoronary saphenous vein graft patency[J].Am J Cardiol,2007,99(8):1087-1089.
[6]ERIKSSEN G,L IESTOL K,BJORNHOLT J,et al.Changes in physical fitness and changes in mortality[J].Lancet,1998,352 (9130):759-762.
[7]FUH G,L IB,CROWLEY C,et al.Requirements for binding and signaling of the kinase domain recep tor for vascular endothelial grow th factor[J].JBiol Chem,1998,273(18):11197-11204.
[8]GAV IN T P,DREW J L,KUBIK CJ,et al.Acute resistance exercise increases skeletalmuscle angiogenic grow th facto r exp ression[J].Acta Physiol(Oxf),2007,191(2):139-146.
[9]GAV IN T P,ROBINSON CB,YEAGER R C,et al.Angiogenic grow th facto r response to acute systemic exercise in human skeletal muscle[J].J App l Physiol,2004,96(1):19-24.
[10]GUSTAFSSON T,AM ELN H,FISCHER H,et al.VEGF-A splice variants and related receptor exp ression in human skeletalmuscle following submaximal exercise[J].J Appl Physiol, 2005,98(6):2137-2146.
[11]GUSTAFSSON T,PUN TSCHART A,KA IJSER L,et al.Exercise-induced exp ression of angiogenesis-related transcription and grow th factors in human skeletalmuscle[J].Am J Physiol Heart Circ Physiol,1999,276(2):H 679-H685.
[12]GUSTAFSSON T,RUNDQV IST H,NORRBOM J,et al.The influence of physical training on the angiopoietin and VEGF-A system s in human skeletalmuscle[J].J App l Physiol,2007;103 (3):1012-1020.
[13]LEE S J,LEE S Y,JEON H S,et al.Vascular endothelial grow th factor gene polymorphisms and risk of p rimary lung cancer[J].Cancer Epidemiol Biomarkers Prev,2005,14(3): 571-575.
[14]LEV Y A P,LEV Y N S,WEHNER S,et al.Transcriptional regulartion of the rat vascular endothelial grow th factor gene by hypoxia[J].JBiol Chem,1995,270(22):13333-13340.
[15]LLOYD P G,PRIOR B M,YANG H T,et al.Angiogenic grow th facto r exp ression in rat skeletal muscle in response to exercise training[J].Am J Physiol Heart Circ Physiol,2003, 284(5):H1668-1678.
[16]LU D,KUSSIE P,PYTOWSKIB,et al.Identification of the residues in the extracellular region of KDR important for interaction with vascular endothelial grow th factor and neutralizing anti-KDR antibodies[J].J Biol Chem,2000,275(19):14321-14330.
[17]PAGES G,POU YSSEGUR J.Transcrip tional regulation of the vascular endothelial grow th factor gene-a concert of activating factors[J].Cardiovasc Res,2005,65(3):564-573.
[18]PRIOR S J,HAGBERG J M,PA TON C M,et al.DNA sequence variation in the p romoter region of the VEGF gene impacts VEGF gene exp ression and maximal oxygen consumption [J].Am J Physiol Heart Circ Physiol,2006,290(5):H1848-H1855.
[19]RICHARDSON R S,WAGNER H,MUDAL IAR S R,et al. Exercise adap tation attenuates VEGF gene exp ression in human skeletal muscle[J].Am J Physiol Heart Circ Physiol, 2000,279(2):H772-H778.
[20]RODAS G,ERCILLA G,JAV IERRE C,et al.Could the A 2A 11 human leucocyte antigen locus correlate with maximal aerobic power?[J]Clin Sci(Colch),1997,92(4):331-333.
[21]SAUNDERS P U,PYNE D B,RICHARD D T,et al.Factors affecting running economy in trained distance runners[J]. Spo rts Med,2004,34(7):465-485.
[22]SHA HBAZIM,FRYER A A,PRAV ICA V,et al.Vascular endothelial grow th factor gene polymorphismsare associated with acute renalallograft rejection[J].J Am Soc Nephrol,2002,13 (1):260-264.
[23]STEPHEN J C,SCHNEIDER J A,TANGUA Y D A,et al. Haplotype variation and linkage disequilibrium in 313 human genes[J].Science,2001,293(5529):489-493.
[24]TIMMONSJ A,JANSSON E,FISHER H,et al.Modulation of extracellular matrix genes reflects themagnitude of physiological adaptation to aerobic exercise training in humans[J].BMC Biology,2005,3(2):19-28.
[25]WANG Y,ZHENG Y,ZHANG W,et al.Polymorphisms of KDR gene are associated with coronary heart disease[J].J Am Coll Cardiol,2007,50(8):760-767.
[26]WESTON A R,MBAMBO Z,M YBURGH K H.Running economy of African and Caucasian distance runners[J].M ed Sci Spo rts Exe,2000,32(6):1130-1134.
[27]ZHANG W L,SUN K,WANG Y,et al.Interaction of the Ile297 variant of vascular endothelial grow th factor receptor-2 gene and homocysteine on the risk of stroke recurrence[J].Circulation,2007,116(Supp l):521.
Association Study of VEGF and Its Receptors Genes Polymorphisms with Aerobic Performance Phenotypes
N IE Jing1,HU Yang2,XI Yi3,WEN Li3,HE Zi-hong4
Objective:To exp lo re the association between VEGF and its recep to rs genes polymorphisms and aerobic perfo rmance phenotypes.Methods:102 young men of Han nationality in northern China underwent an 18-week long-distance running p rogram.O2max,RE and left ventricular structure and function were tested befo re and after training.The genotypes were examined in VEGF and its recep tors genes by PCR-RFLP.Results:It was found that no significant differences in baseline levels and the response to training ofO2max and RE between VEGF and its recep to rs genotypes were observed.However,there was an association between VEGF and its recep tors genes polymorphism s and left ventricular structure and function.The carriers of the A allele(CA+AA genotypes)in SNP/C-2578A of VEGF gene, AA genotypes in SNP/A+193019G of VEGFR1 gene and A T or AA genotypes in SNP/A+ 18487T of VEGFR2 gene were better in left ventricular structure and function.Conclusion: The-2578C/A genetic polymo rphism of VEGF gene,the+193019A/G genetic polymorphism of VEGFR1 gene and the+18487A/T genetic polymo rphism of VEGFR2 gene may be the genetic markers for the response of left ventricular function to 18-week long-distance running.But the sample size would need to be expanded further.
VEGF;receptor;gene polymorphism;˙VO2max;running econom y;lef t ventricular function;aerobic exercise
G804.7
A
1000-677X(2010)08-0077-08
2010-07-14;
2010-08-09
国家科技部资助课题(2003BA 904B04)。
聂晶(1979-),女,江西南昌人,讲师,博士,主要研究方向为低氧训练、运动分子生物学,Tel:(0791)8506324,E-mail:niejing1214@163.com。
1.江西师范大学,江西南昌330022;2.北京体育大学,北京100084;3.天津体育学院,天津300381;4.国家体育总局体育科学研究所,北京100061 1.Jiangxi Normal University,Nanchang 330022,China; 2.Beijing Sport University,Beijing 100084,China;3. Tianjin Institute of Physical Education,Tianjin 300381, China;4.China Institute of Sport Science,Beijing 100061,China.
