流感病毒H1N1和H5N1感染小鼠小胶质细胞与星形胶质细胞诱导趋化因子转录水平上调①
2010-02-06王革非李卫中张丹桂陈幼莹陈小璇李康生
王革非 李卫中 张 衡 曾 俊 张丹桂 陈幼莹 陈小璇 李康生
(汕头大学医学院微生物学与免疫学教研室 广东高校免疫病理重点实验室,汕头 515041)
流感病毒H1N1和H5N1感染小鼠小胶质细胞与星形胶质细胞诱导趋化因子转录水平上调①
王革非 李卫中 张 衡 曾 俊 张丹桂 陈幼莹 陈小璇 李康生
(汕头大学医学院微生物学与免疫学教研室 广东高校免疫病理重点实验室,汕头 515041)
目的:探讨胶质细胞感染流感病毒后的天然免疫反应,检测流感病毒H1N1和H 5N1体外感染小鼠小胶质细胞和星形胶质细胞,是否会诱导胶质细胞趋化因子转录水平的变化及其规律。方法:从新生小鼠大脑皮质分离培养神经胶质细胞,并进一步纯化小胶质细胞和星形胶质细胞,经纯度鉴定后,用感染复数为2的流感病毒H1N1和H5N1进行体外感染,8小时后用免疫荧光检测流感病毒核蛋白(NP)的表达,以确认感染细胞比例。在感染早期(6小时)和感染中期(24小时)分别提取细胞RNA,检测趋化因子转录水平的变化。结果:分离得到小鼠的小胶质细胞和星形胶质细胞,病毒感染后超过95%的细胞可以被感染,感染后的小胶质细胞与星形胶质细胞的CCL-3、CCL-5、CXCL-2、CXCL-9和CXCL-10的转录水平发生不同程度的上调,其中CXCL-10的上调幅度最为明显,禽流感病毒H5N1感染能诱导更强烈的上调反应。结论:流感病毒H1N1和H5N1感染小鼠小胶质细胞与星形胶质细胞,可诱导趋化因子转录水平上调。
小胶质细胞;星形胶质细胞;流感病毒;趋化因子
流感病毒的感染除能造成人呼吸系统疾病外,在部分感染人群中会出现中枢神经系统(Central nervous system,CNS)病变[1],即流感相关脑病/脑炎(Influenza associated encephalopathy or encephalitis),是指在急性流感过程中伴随中枢神经系统功能障碍的一种临床综合征,起病快速、病情凶险,主要的临床表现为癫痫发作、意识消退、昏迷等,严重时引起神经后遗症或死亡,如雷耶氏综合征(Reye's syndrome)、出血性休克与脑病综合征(Hemorrhagic shock and encephalopathy syndrome)和急性坏死性脑病(A-cute necrotizing encephalopathy)等[2-4]。在流感患者,尤其是儿童中,具有一定的发病率和死亡率[4]。流感病毒可通过被破坏的血脑屏障、嗅球-三叉神经节、内脏-神经途径及其它可能途径感染CNS[4],在流感相关脑病患者的脑脊液和脑组织中可以检测到流感病毒RNA及相关抗原的存在[5,6]。但流感相关脑病的发病机制仍不明确。
在流感相关脑病病人的脑脊液中,发现促炎症细胞因子的浓度升高[6-8]。在CNS中,神经胶质细胞,尤其是小胶质细胞和星形胶质细胞是细胞因子分泌的主要来源[9]。在流感病毒感染中,促炎症细胞因子释放的强度被认为与疾病的严重程度存在可能的正相关联。中枢神经系统中细胞因子及趋化因子在免疫及神经内分泌网络的连接和调节中起到十分重要的作用,促炎症因子的过量释放不仅造成神经元和胶质细胞的免疫损伤和凋亡,还会影响外周免疫并诱导全身性反应[10,11]。中枢神经系统受流感病毒感染后,胶质细胞过度释放的促炎症细胞因子,可能会诱导严重的中枢神经系统紊乱及全身性症状。
因此,研究流感病毒是否会诱导胶质细胞发生促炎症细胞因子反应,明确促炎症因子与趋化因子的类型,将有助于理解胶质细胞在流感病毒感染CNS时的功能和作用。我们前期实验发现流感病毒感染胶质细胞可诱导 IL-1β、TNF-α和 IL-6的过度释放[12],流感病毒H5N1在肺部诱发的细胞因子风暴中,除上述3种促炎症因子外,IP-10、CCL-5等趋化因子的表达亦发生过度释放[13-15],因此,本研究利用人流感病毒H1N1和禽流感病毒H5N1感染胶质细胞,检测是否出现趋化因子表达的上调,明确小胶质细胞和星形胶质细胞在流感病毒感染后,所发生的促炎症因子反应中趋化因子类型与变化,为阐明流感相关脑病的发病机制提供依据。
1 材料与方法
1.1 实验动物 1至2日龄无特异病原(Specific pathogen free,SPF)的BALB/c小鼠由汕头大学医学院实验动物中心提供。
1.2 流感病毒毒株 人A型流感病毒A/Shantou/169/06(H1N 1)分离于2006年汕头大学医学院第一附属医院儿科门诊上呼吸道感染儿童[16]。禽流感病毒A/Chicken/Guangdong/1/05(H5N1)分离于2005年广东省。流感病毒在MDCK细胞和9~10日龄的鸡胚进行初步增殖收获病毒母液,经标准血清/血凝抑制实验鉴定病毒亚型,病毒为本室保存。利用10%醛化的豚鼠红细胞及鸡红细胞悬液,使用红细胞吸附/洗脱以及超滤的方法对A型流感病毒A/Shantou/169/06(H1N1)和A/Chicken/Guangdong/1/05(H 5N1)进行纯化[1-3]。经超滤离心管(M illipore)更换病毒缓冲体系后,-80℃保存,测定病毒滴度。
1.3 新生小鼠大脑皮层神经胶质细胞的培养 将1至2天新生小鼠大脑皮层组织取出,于DMEM/F12(1∶1,v/v)培养基中吹打分散后,用50μm滤器过滤得到细胞悬液。37℃放置30分钟去除成纤维细胞。未贴壁细胞于37℃、5%CO2培养箱中培养。接种后第2天和第4天,弃去未贴壁的细胞,更换培养基。以后每5~7天更换一次培养基,至胶质细胞长成单层。
1.4 小鼠大脑皮层小胶质细胞与星形胶质细胞的分离 长成单层的胶质细胞0.05%胰酶37℃作用约10分钟,将含有星形胶质细胞的胰酶液经终止、离心后培养。1天将细胞培养瓶固定于定轨摇床(上海智诚)中 37℃、200 r/min、振幅30 mm水平震荡60分钟,弃悬浮细胞,加入10%FBS的DMEM/F12培养基继续培养1天后,按上述方法再次进行震荡分离,得到纯化后的星形胶质细胞。
长成单层的混合胶质细胞用0.1%胰酶37℃作用约15分钟,镜下观察星形胶质细胞脱落后,弃去消化液,DMEM/F12清洗 2遍,加入胰酶消化液(0.25%trysin/0.02%EDTA)37℃作用5~10分钟,得到小胶质细胞组分[6],经终止、离心后用10%FBS的DMEM/F12培养基培养。
分离后的小胶质细胞与星形胶质细胞,使用CD11b和胶质细胞纤维酸性蛋白(Glial fibrillary acidic protein,GFAP)抗体进行免疫荧光法检测纯度。星形胶质细胞与小胶质细胞按每孔105个细胞接种12孔细胞培养板,用于后续实验。
1.5 流感病毒感染神经胶质细胞 小鼠小胶质细胞和星形胶质细胞,用0.5ml含有2×105pfu流感病毒(H1N1和H5N1)的病毒液感染,感染复数(Multiplicity of infection,MOI)为2,37℃吸附1小时。吸附结束后,每孔加入1m l无血清DMEM/F12,37℃、5%CO2培养箱中培养。设立未感染孔为阴性对照。感染/刺激后6小时和24小时收集细胞,经PBS清洗后,用于RNA提取、逆转录及半定量PCR。
1.6 流感病毒核蛋白的免疫荧光检测 为检测流感病毒是否感染在小胶质细胞和星形胶质细胞,将感染8小时的胶质细胞,经固定与穿孔后,利用抗甲型流感病毒NP单克隆抗体(Vector Lab)为一抗,Cy3标记的抗小鼠IgG抗体(碧云天生物技术研究所)为二抗,对流感病毒核蛋白NP免疫荧光检测。
1.7 胶质细胞RNA的提取及反转录 感染后的胶质细胞,使用RNeasy Minikit(Qiagen)试剂盒提取细胞的RNA,实验方法参照Qiagen公司说明书。取1μg的 RNA,用SuperScript Ⅲ RNaseH-Reverse Transcriptase(Invitrogen)和随机六碱基引物(hexaprimers,大连宝生物工程公司)反转录,反应体系为20μl。25℃反应5分钟、50℃继续反应60分钟完成反转录,70℃15分钟灭活反转录酶,得到反转录后的cDNA用于半定量PCR反应。
1.8 小鼠胶质细胞感染流感病毒后炎症相关因子的半定量PCR检测 cDNA用特异性引物进行半定量PCR,用于检测小胶质细胞及星形胶质细胞在流感病毒感染后,趋化分子(CCL-2、CCL-3、CCL-5、CXCL-2、CXCL-9、CXCL-10)的表达变化情况。引物见表1,退火温度根据引物碱基长度组成及预实验结果为56℃~60℃,反应循环次数根据预实验结果为32~40循环。PCR使用预混的PCR反应液(MasterM ix,北京天根),反应体系25μl,其中,Premix Taq Mixture 12.5 μl、引物 12 μl(0.5 μmol/L)、cDNA 0.5 μl,按照产品说明书和引物参数进行PCR反应。PCR产物经电泳、拍照后,用BandScan4.3分析条带的灰度值。各个基因的PCR产物灰度值同样品内参GAPDH基因PCR产物灰度值进行均一化,以未感染对照样品相应基因PCR产物均一化后的比值为100%,计算感染样品基因转录变化。
2 结果
2.1 小鼠胶质细胞的分离与培养 图1所示,小鼠原代培养的混合胶质细胞生长约20~25天,细胞长成单层,即可用于分离小胶质细胞和星形胶质细胞。利用不同浓度胰酶的分步消化可得到纯化的小胶质细胞,通过低浓度胰酶分离和摇床震荡培养可得到纯化的星形胶质细胞。小胶质细胞选用细胞特异性表面标志CD11b进行免疫荧光检测,星形胶质细胞用GFAP为细胞特异性标志进行检测,细胞均出现特异性的荧光染色,相应的对照细胞或一抗缺失样品则没有荧光出现,小胶质细胞和星形胶质细胞的纯度均超过95%。
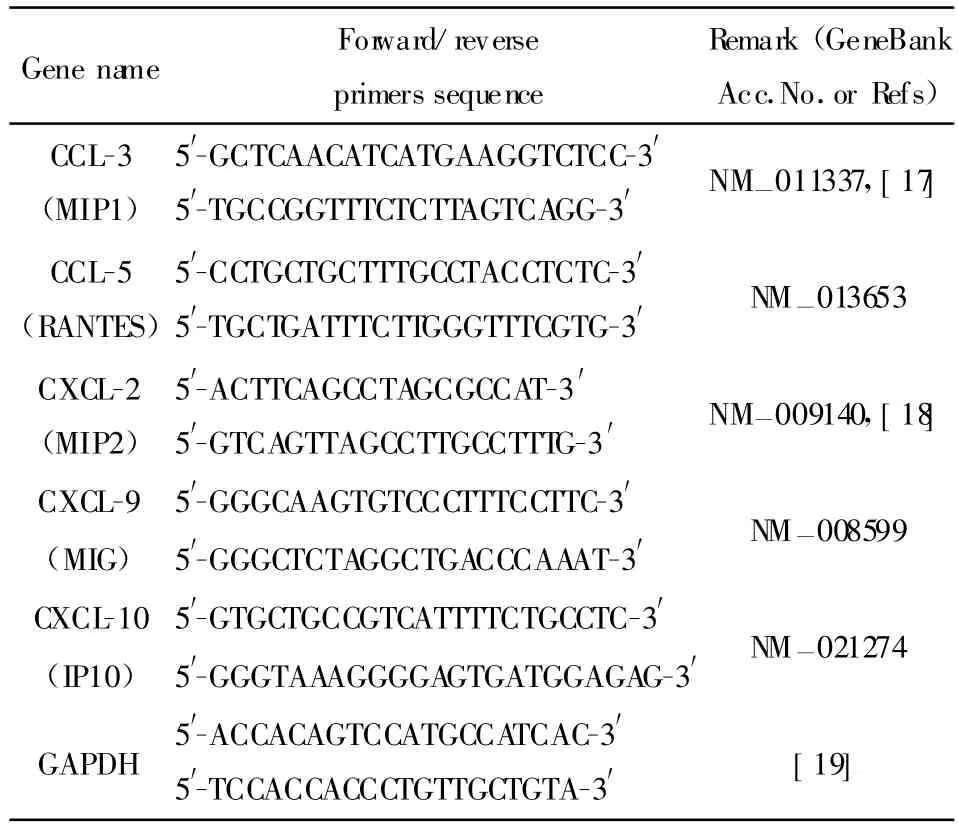
表1 趋化因子PCR引物列表Tab.1 The detail information of the primers of chemokines
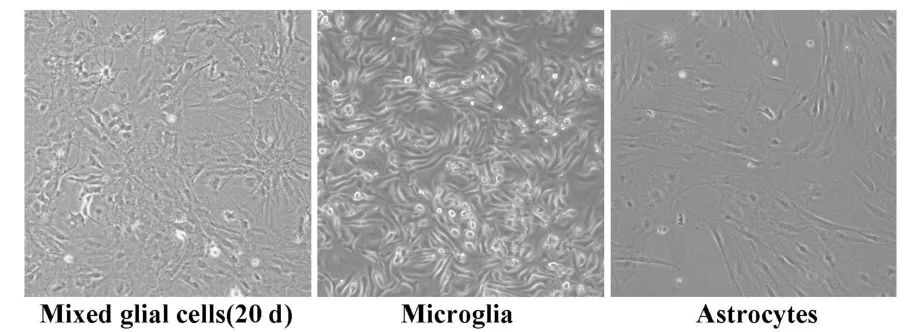
图1 混合胶质、小胶质细胞和星形胶质细胞的分离与培养(40×)Fig.1 Isolation and culture ofm ixed glial cells,m icroglia,and astrocytes(40×)
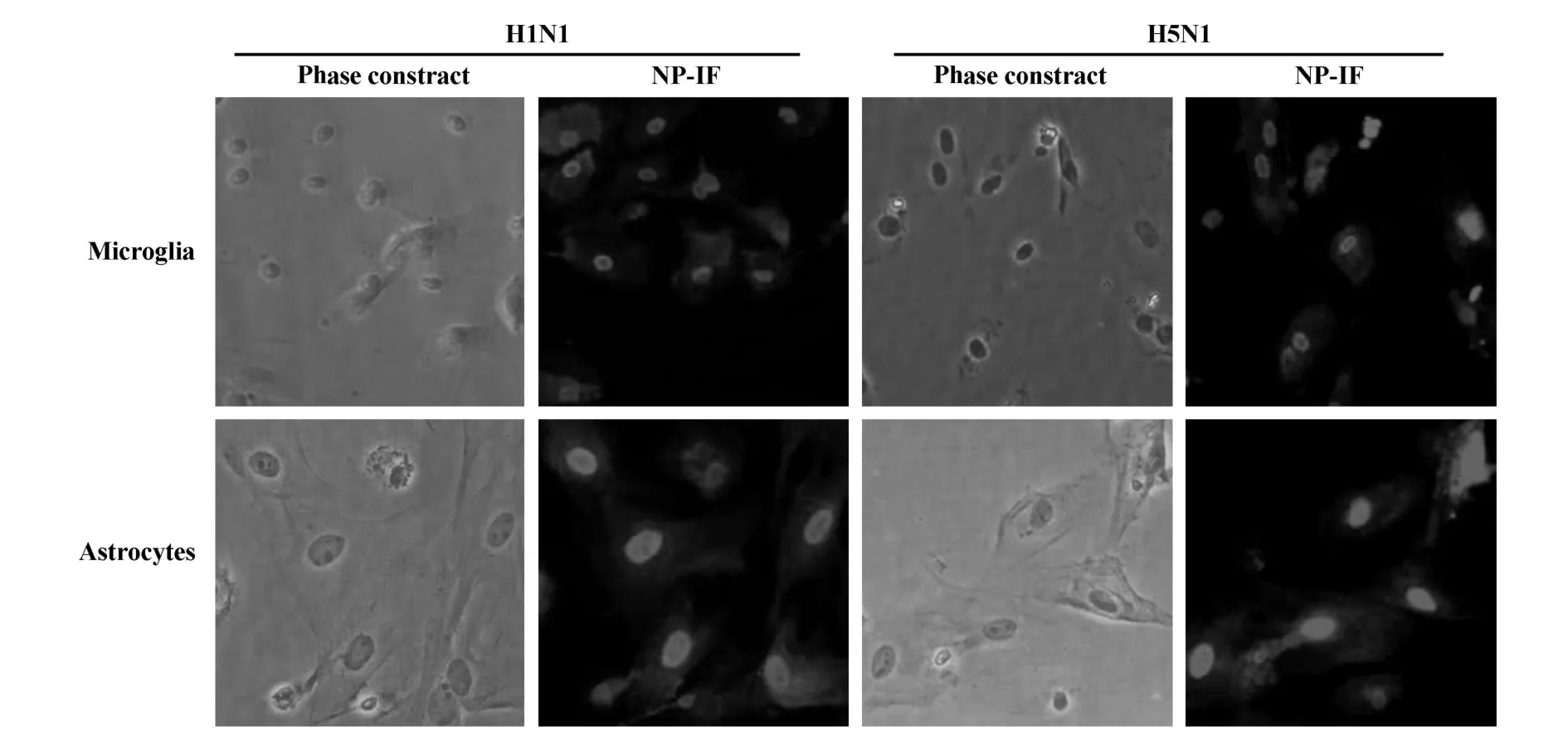
图2 胶质细胞感染流感病毒 H1N1和H5N1 8小时后的NP蛋白染色(200×)Fig.2 NP immunofluorescence staining of H1N1 and H5N1 infected microglia and astrocy tes at 8 h p.i.(200×)
2.2 流感病毒感染胶质细胞的NP蛋白检测 小胶质细胞和星形胶质细胞经流感病毒H 1N1和H5N1吸附后继续培养8小时,对甲型流感病毒的NP蛋白进行免疫荧光染色,如图2所示,在H1N1和H5N1吸附培养的小胶质细胞和星形胶质细胞的细胞核中,均出现阳性的荧光信号,表明流感病毒在吸附细胞后,能够进入细胞进行相应的生物合成和复制,表达流感病毒复制所需的NP蛋白。当流感病毒H1N1和H5N1在MOI为2时,首轮培养即可以导致超过95%以上的细胞感染流感病毒。
2.3 流感病毒感染胶质细胞诱导趋化因子转录水平上调 使用半定量PCR检测小胶质细胞及星形胶质细胞在流感病毒感染后,趋化因子CCL-3、CCL-5、CXCL-2、CXCL-9、CXCL-10的表达变化情况。根据预实验的结果,对引物进行了优化,选取了扩增效率高、特异性强的引物用于该实验,确定了相应的引物最适退火温度,同时选择PCR产物的对数增长阶段后期的循环数为最适循环次数,对上述基因的表达变化进行检测(图3)。
对PCR产物电泳照片进行灰度分析,各样本以32个循环的内参基因GAPDH的灰度值进行均一化处理,然后各基因以对照组细胞的表达量为参照(100%)进行比值处理,基因的表达变化结果见图4。结果表明,小胶质细胞和星形胶质细胞在流感病毒感染后6小时和24小时,趋化因子的表达发生不同程度的上调。图3和图4的结果提示,流感病毒不同亚型在同等感染条件下,H5N1比H1N1能够诱导胶质细胞更强烈的炎症因子反应,趋化因子的表达上调幅度更大。
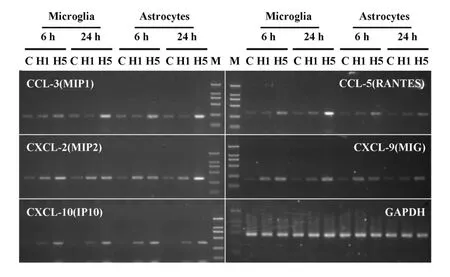
图3 胶质细胞感染流感病毒H1N1和H5N1后趋化因子电泳图Fig.3 The PCR products electrophoresis photo of chemokines of in fluenza H 1N1 and H5N1 infected glial cells
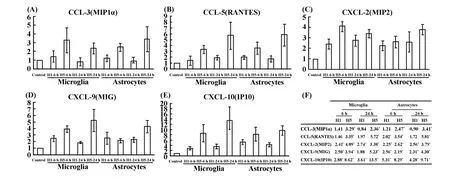
图4 胶质细胞感染流感病毒H1N1和H5N1后趋化因子的转录水平变化Fig.4 The transcrip t levels of chemokines of influenza H1N1 and H5N1 in fected glial cells
3 讨论
尽管流感病毒能够感染中枢神经系统,流感病毒相关的脑病与脑炎综合征的发病机制仍然不明确。流感病毒相关脑病与脑炎综合征患者的脑脊液和血浆中,促炎症细胞因子水平异常升高[6-8]。异常升高的促炎症细胞因子常常引发局部或系统性的免疫病理损伤[10,11]。可以推测在流感病毒相关脑病与脑炎综合征的发生和发展中,不仅存在着病毒的直接破坏作用,同时可能伴随着炎症细胞因子大量释放而造成的免疫病理损伤。
流感病毒,尤其是H 5N1毒株,在体外可诱导多种细胞释放高浓度的促炎症细胞因子,在体内的局部器官和血浆中也发现存在着大量促炎症细胞因子的积累,大量释放和积累的促炎症因子,如TNF-α、IL-1β等,通过细胞因子的级联反应(Cytokine cascade),产生以细胞因子分泌失调和过度释放为表现的细胞因子风暴效应(Cytokine storm),从而可能引发相应的免疫病理损伤,这种以促炎症细胞因子过度释放导致的细胞因子风暴被认为是高致病性H5N1引起严重疾病的可能原因[13-15]。
小胶质细胞和星形胶质细胞在中枢神经系统的免疫应答过程起关键作用,同时也是中枢神经系统中细胞因子的主要分泌来源[9]。因此,探索小胶质细胞和星形胶质细胞在流感病毒感染后所发生的变化及免疫反应,对理解流感病毒相关的脑病与脑炎综合征的发病机制有一定的帮助。本研究以小鼠大脑的小胶质细胞和星形胶质细胞为靶细胞,用人流感病毒H1N1和禽流感病毒H5N1感染细胞,检测细胞趋化因子转录表达水平。
对人流感病毒H1N1和禽流感病毒H5N1吸附后8小时的胶质细胞进行A型流感病毒NP蛋白免疫荧光染色,发现在细胞核内存在大量表达的NP蛋白,表明H1N1和H 5N1病毒可以感染星形胶质细胞和小胶质细胞,并在细胞中进行了病毒复制相关的蛋白表达。实验结果究表明,流感病毒感染后趋化因子 CCL-3、CCL-5、CXCL-2 、CXCL-3、CXCL-10 等的表达出现上调。其中,禽流感病毒H5N1感染的神经胶质细胞,其促炎症细胞因子及趋化因子的表达上调水平高于H 1N1感染的细胞。
流感病毒感染所诱导神经胶质细胞的炎症因子反应中,除了促炎症因子上调外[12],本研究亦发现趋化因子的转录水平发生上调。促炎症因子和趋化因子的上调,能够导致其它炎症细胞趋化因子、粘附分子、急性时相蛋白和组织蛋白酶等的合成,当其表达上调过于强烈时,则引发细胞因子风暴。趋化因子可以募集并活化巨噬细胞、单核细胞等免疫细胞(如 CCL-3/MIP1α,CXCL-2/MIP2,CXCL-9/M IG 等)、激活细胞抗病毒免疫(如CCL-5/RANTES,CXCL-10/IP10等),从而抵御病毒攻击,但趋化因子表达的异常上升会使免疫细胞过度集中及活化而导致免疫病理损伤。胶质细胞感染H5N1病毒后,促炎症细胞因子以及趋化因子的水平发生了强烈的上调。在流感病毒感染中,促炎症细胞因子释放的强度被认为与疾病的严重程度存在可能的正相关联[15]。细胞因子及趋化因子在免疫及神经内分泌网络的连接和调节中起到十分重要的作用[10,11]。中枢神经系统中,促炎症因子的过量释放不仅造成神经元和胶质细胞的免疫病理损伤和凋亡,还会影响外周免疫并诱导系统性反应[6,20]。因此,中枢神经系统受H5N1感染可以诱导比人流感病毒感染更严重的中枢神经系统紊乱及系统性疾病。
促炎症细胞因子与趋化因子的过度释放,可能参与了对神经胶质细胞的直接破坏作用和对脑部组织的间接免疫病理损伤作用,这些可能是诱导及加重流感相关脑病和脑炎的重要因素。另一方面,应用具有多向性生物活性的免疫抑制因子IL-10以及其它促炎症因子抑制剂,如IL-1受体拮抗肽(IL-1 receptor antagonist,IL-1ra)、TNF拮抗肽等,来平衡或抑制严重的促炎症反应,为缓解或减轻细胞因子风暴可能造成的继发性免疫病理损伤提供思路。
1 Nakai Y,Itoh M,M izuguchi Metal.Apoptosis andm icroglial activation in influenza encephalopathy[J].Acta Neuropathol,2003;105(3):233-239.
2 Studahl M.Influenza virus and CNS manifestations[J].J Clin V irol,2003;28(3):225-232.
3 Gooskens J,Kuiken T,Claas ECetal.Severe influenza resembling hemorrhagic shock and encephalopathy syndrome[J].JClin Virol,2007;39(2):136-140.
4 Toovey S.Influenza-associated centralnervoussystem dysfunction:a literature review[J].TravelMed InfectDis,2008;6(3):114-124.
5 Fujimoto S,KobayashiM,UemuraOetal.PCR on cerebrospinal fluid to show influenza-associated acute encephalopathy or encephalitis[J].Lancet,1998;352(9131):873-875.
6 Ito Y,Ichiyama T,Kimura Hetal.Detection of influenza virus RNA by reverse transcription-PCR and proinflammatory cytokines in influenzavirus-associated encephalopathy[J].JMed V irol,1999;58(4):420-425.
7 Kawada J,KimuraH,Ito Yetal.System ic cytokine responses in patients with influenza-associated encephalopathy[J].JInfectDis,2003;188(5):690-698.
8 Ichiyama T,Isumi H,OzawaHetal.Cerebrospinal fluid and serum levels of cytokines and soluble tumor necrosis factor receptor in influenza virusassociated encephalopathy[J].Scand J Infect Dis,2003;35(1):59-61.
9 Jack CS,ArbourN,Manusow Jetal.TLR signaling tailors innate immune responses in human m icroglia and astrocytes[J].J Immunol,2005;175(7):4320-4330.
10 Nunoi H,Mercado M R,Mizukam i Tetal.Apoptosis under hypercytokinem ia isa possible pathogenesis in influenza-associated encephalopathy[J].Pediatr Int,2005;47(2):175-179.
11 Hu S,Peterson P K,Chao C C.Cytokine-mediated neuronal apoptosis[J].Neurochem Int,1997;30(4-5):427-431.
12 Wang G F,Zhang J,LiW Zetal.Apoptosis and pro-inflammatory cytokine responses of mousem icroglia and astrocytes induced by human H1N1 and avian H 5N1 influenza viruses[J].CellMol Immunol,2008;5(2):113-120.
13 Konstantinos A P,Sheridan JF.Stressand influenza viral infection:modulation of proinflammatory cytokine responses in the lung[J].Respir Physiol,2001;128(1):71-77.
14 Osterholm MT.Preparing for the next pandemic[J].N Engl JMed,2005;352(18):1839-1842.
15 Cheung CY,Poon L L,Lau A Setal.Induction of proinflammatory cytokines in humanmacrophagesby influenza A(H5N1)viruses:amechanism for the unusual severity of human disease?[J].Lancet,2002;360(9348):1831-1837.
16 Wang,G F,Lin S Y,Zhang Hetal.Apobec 3F and apobec 3G have no inhibition and hypermutation effect on the human influenza A virus[J].Acta Virol,2008;52:193-194.
17 MylesM H,Dieckgraefe B K,Criley JMetal.Characterization of cecal gene expression in a differentially susceptiblemouse model of bacterialinduced inflammatory bowel disease[J].Inflamm Bowel Dis,2007;13:822-836.
18 Luo Y,Lloyd C,Gutierrez-Ramos JCetal.Chemokine amplification in mesangial cells[J].J Immunol,1999;163:3985-3992.
19 Blaeser F,Bryce PJ,Ho Netal.Targeted inactivation of the IL-4 receptor alpha chain I4R motif promotes allergic airway inflammation[J].J Exp Med,2003;198:1189-1200.
20 SilviaG C,MarianaM,V irginia ERetal.Cytokinesand the immune neuroendocrine network:Whatdidwe learn from infection and autoimmunity?[J].Cytokine Growth F R,2007;18:125-134.
[收稿2009-05-26 修回2009-08-19]
(编辑 张晓舟)
Upregulation of chemokine transcriptive levels induced by avian H 5N1 and human H1N1 in fluenza viruses inmousem icroglia and astrocytes
WANGGe-Fei,LIWei-Zhong,ZHANGHeng,ZENGJun,ZHANGDan-Gui,CHENYou-Ying,CHENXiao-Xuan,LI Kang-Sheng.DepartmentofMicrobiologyandImmunology,theKeyImmunopathologyLaboratoryofGuangdongProvince,ShantouUniversityMedicalCollege,Shantou515041,China
Objective:To investigate the innate immune response of influenza virus-infected glial cells,the transcription levels in chemokines inmousemicroglia and astrocyteswere detected which pre-infected by human H1N1 or avian H5N1 influenza viruses.Methods:The glial cells isolated from neonatalm ice cerebral cortex were cultured and furthermicrogliaand astrocyteswere purified.The primarymouse microglia and astrocyteswere infected in vitroby H1N1or H5N1 influenza virusesinamultip licity of infection(MOI)2.Eighthours post infection,the influenza virus nucleoprotein(NP)was detected by immunofluorescence to identify the proportion of infected cells.The cellular RNA were extracted at 6 h and 24 h to detect the transcriptional levelof chemokines by semi-quantitative RT-PCR.Results:More than 95%of the microglia and astrocyteswhich isolated from micewere infected.The transcription levels of CCL-3,CCL-5,CXCL-2,CXCL-9and CXCL-10 from infectedm icroglia and astrocyteswere upregulated.Futhermore,themRNA levelof CXCL-10 increasedmuchmore.In addition,avian H5N1 influenza virus could inducemore strongerup regu lation of those chemokines than human H1N1 did.Conclusion:Themousemicroglia and astrocyteswhich are infected by H1N1 influenza virus orH5N1 influenza virus could induce upregulation of transcription levelof chemokines.
Microglia;Astrocyte;Influenza virus;Chemokine
R392.6 R373.1
A
1000-484X(2010)01-0029-06
①本文受国家自然科学基金项目(30771988,30972766)、广东省自然科学基金(9451503102003499,8151503102000022)和广东高校优秀青年创新人才培育项目(LYM08056)资助
王革非(1978年-),男,博士,讲师,主要从事神经免疫与抗感染免疫研究,E-mail:gfwangcn@gmail.com;
及指导教师:李康生(1953年-),男,教授,博士生导师,主要从事神经免疫与抗感染免疫研究,E-mail:ksli@stu.edu.cn。