肠道菌群介导的神经递质5-羟色胺合成对放射性肠炎的影响及可能机制
2025-01-25褚良妹姬倩胡静任永飞王旭戴春华
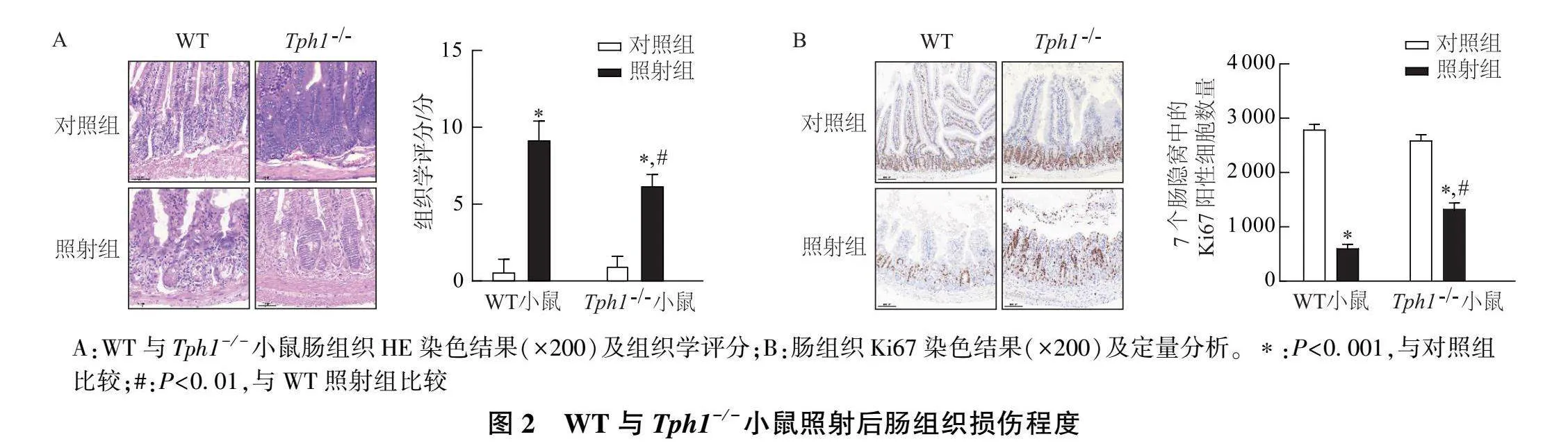
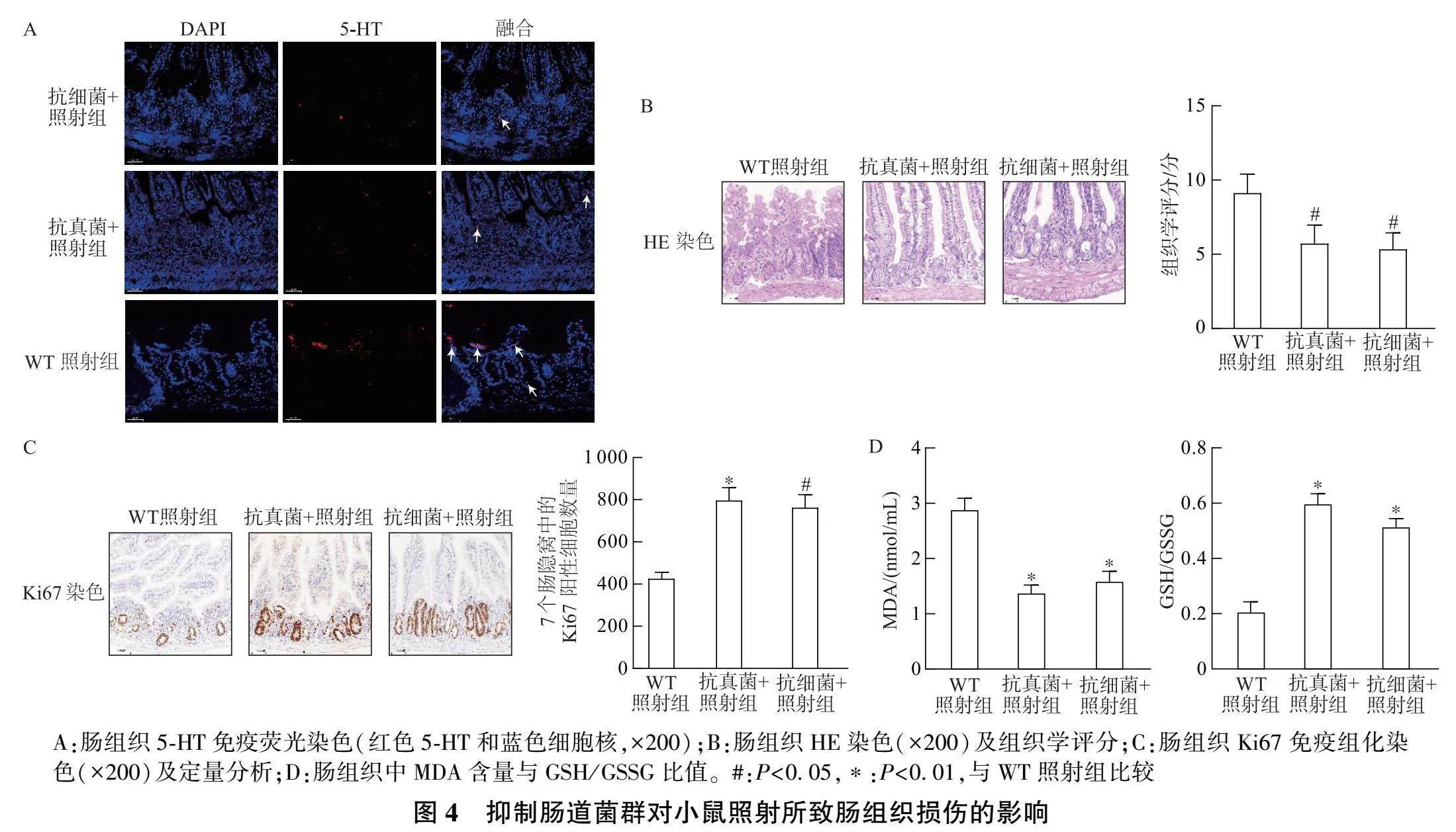

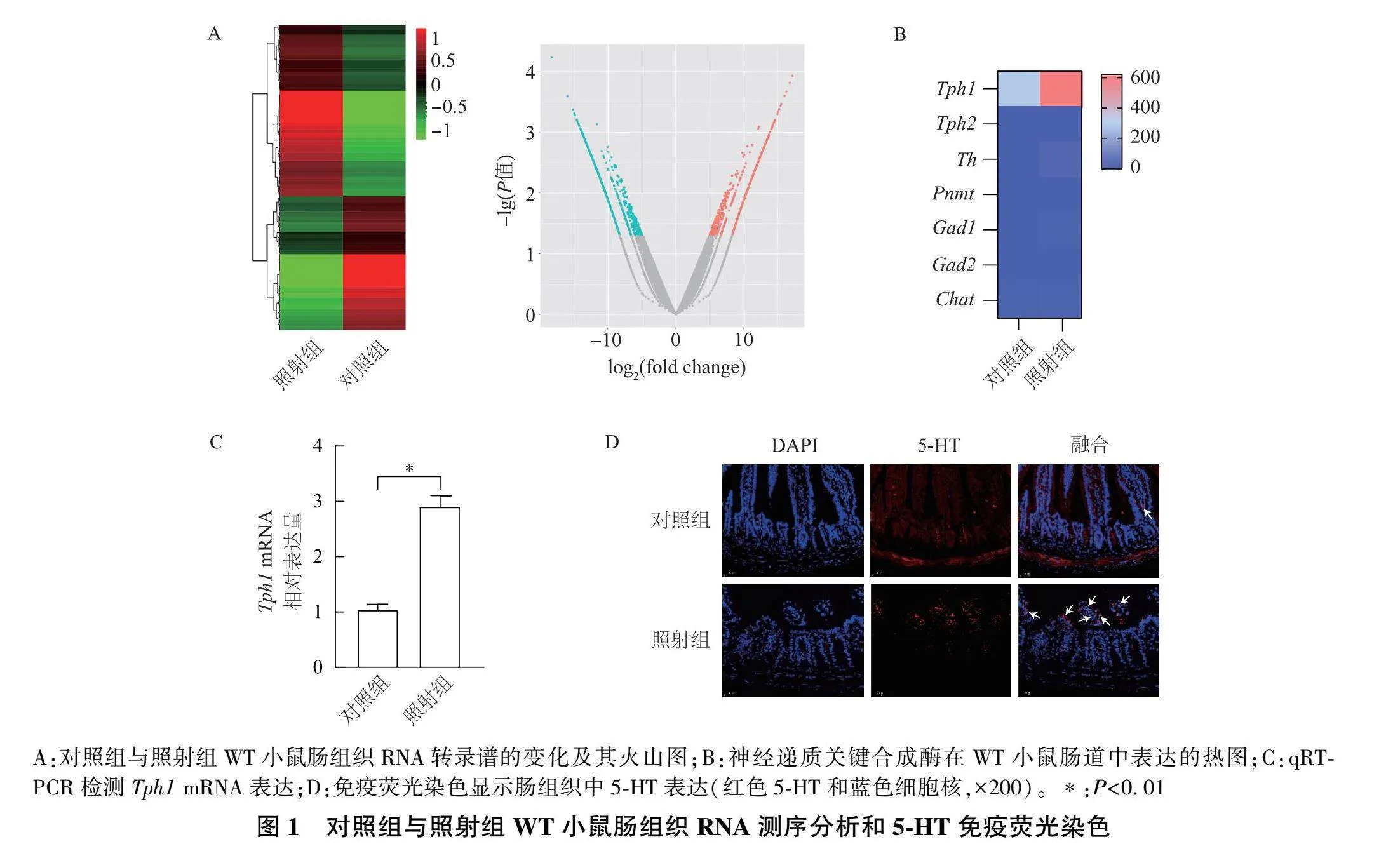
[摘要] 目的: 探讨神经递质5-羟色胺(5-HT)对放射性肠炎的影响及其机制。方法: 对对照组(未照射)和照射组的野生型(WT)小鼠肠组织进行RNA测序,分析其转录组变化,并用qRT-PCR进行验证;采用免疫荧光检测两组WT小鼠的肠组织5-HT水平。对WT和色氨酸羟化酶1( Tph1 )基因敲除( Tph1 -/- )小鼠进行照射,取肠组织行HE染色和Ki67免疫组化染色,比较肠组织损伤程度。对WT照射组和 Tph1 -/- 照射组小鼠肠组织进行RNA测序,分析其转录组变化。小鼠抗细菌和抗真菌处理后进行照射,采用免疫荧光检测5-HT水平,并取肠组织行HE染色和Ki67免疫组化染色。检测肠组织中丙二醛(MDA)和还原型谷胱甘肽(GSH)/氧化型谷胱甘肽(GSSG)指标。结果: "WT小鼠照射后的肠组织中 Tph1 "mRNA表达显著升高,且5-HT合成增多。WT照射组小鼠肠组织遭受明显损伤,而 Tph1 -/- 照射组小鼠肠组织损伤相对减轻。与WT照射组小鼠相比, Tph1 -/- 照射组小鼠肠组织中的促铁死亡基因显著下调,抑铁死亡基因显著上调,肠组织MDA含量明显下降,GSH/GSSG比值明显上升。经抗细菌和抗真菌处理后照射,WT小鼠肠组织中5-HT减少;同时,照射的肠组织损伤均在一定程度上减轻,且MDA含量下降,GSH/GSSG比值明显上升。结论: 肠道菌群可增加5-HT水平,可能通过诱导铁死亡加重放射性肠炎。
[关键词] 神经递质;5-羟色胺;放射性肠炎;铁死亡;肠道菌群;放射治疗
[中图分类号] R818" [文献标志码] A" [文章编号] 1671-7783(2025)01-0032-07
DOI: 10.13312/j.issn.1671-7783.y240049
[引用格式]褚良妹, 姬倩, 胡静, 等. 肠道菌群介导的神经递质5-羟色胺合成对放射性肠炎的影响及可能机制[J] . 江苏大学学报(医学版), 2025, 35(1): 32-38.
[基金项目]国家自然科学基金资助项目(32170910);镇江市重点研发计划(社会发展)项目(SH2023002)
[作者简介]褚良妹(1995—),女,硕士研究生;戴春华(通讯作者),主任医师,硕士生导师,E-mail: daichunhua8@163.com
Effects and potential mechanisms of intestinal microbiota-mediated synthesis of neurotransmitter 5-hydroxytryptamine on radiation enteritis
CHU Liangmei1 , JI Qian1 , HU Jing1 , REN Yongfei2 , WANG Xu1 , DAI Chunhua1
(1. Department of Radiation Oncology, Affiliated Hospital of Jiangsu University, Zhenjiang Jiangsu 212001; 2. School of Medicine, Jiangsu University, Zhenjiang Jiangsu 212013, China)
[Abstract] Objective: To explore the effect of neurotransmitter 5-hydroxytryptamine (5-HT) on radiation enteritis and its mechanism. Methods: RNA sequencing was performed on intestinal tissues from control and irradiated wild-type (WT) mice to analyze transcriptome changes, and qRT-PCR was performed to verify the selected expression changes of genes. The level of 5-HT in control and irradiated intestinal tissues of WT mice was detected by immunofluorescence. Wild-type (WT) mice and tryptophan hydroxylase 1 ( Tph1 ) knockout ( Tph1 -/- ) mice were both treated with radiotherapy, and the intestinal tissues of mice were performed with HE staining and Ki67 immunohistochemistry staining to compare the degree of intestinal tissue damage between them. RNA sequencing was performed on intestinal tissues from WT irradiation group and "Tph1 -/- "irradiation group to analyze transcriptome changes. After antibacterial and antifungal treatment, WT mice were treated with irradiation, immunofluorescence was performed to detect the level of 5-HT in intestinal tissues. Meanwhile, HE staining and Ki67 staining were conducted. MDA and GSH/GSSG of intestinal tissues were detected. Results: The expression of "Tph1 "mRNA in intestinal tissues from WT mice was significantly increased, and the level of 5-HT was increased after irradiation. The intestinal tissues of WT mice were obviously damaged after irradiation, while the intestinal damage was relatively reduced after "Tph1 "knockout. Compared with WT irradiation group, the ferroptosis-promoting genes were significantly down-regulated and ferroptosis-inhibiting genes were significantly up-regulated in the intestinal tissues of "Tph1 -/- "irradiation mice. MDA was decreased and GSH/GSSG was increased in "Tph1 -/- "irradiation group. After antibacterial and antifungal treatment, the level of 5-HT in intestinal tissues of WT mice treated with irradiation was decreased, and the damage of irradiated intestinal tissues was alleviated to a certain extent. At the same time, MDA was decreased, and GSH/GSSG was increased. Conclusion: The intestinal flora can increase the level of 5-HT, potentially aggravating intestinal tissue injury after irradiation, in which ferroptosis may be involved.
[Key words] neurotransmitter; 5-HT; radiation enteritis; ferroptosis; intestinal flora; radiotherapy
放射治疗已经成为临床广泛应用于治疗各种癌症的重要手段[1] ,其使用高能电离辐射诱导DNA双链断裂,导致肿瘤细胞死亡[2] 。肿瘤细胞增殖迅速,对放疗敏感,且修复速度比正常细胞慢,因此放疗对许多类型的肿瘤都有良好的效果。同时,放疗还会损害正常细胞和组织,尤其是活性增殖细胞。这些细胞对放疗高度敏感,非常容易受到损伤。胃肠道尤其是小肠黏膜,对电离辐射极其敏感,因此放射治疗腹部和骨盆恶性肿瘤很容易引起放射性肠炎,出现如食欲不振、恶心、呕吐、血便、腹泻、黏液便等各种并发症[2-4] ,导致患者的生活质量下降[5] 。目前,放疗诱导肠道损伤的发生和发展机制在很大程度上仍未被探索[6] 。
肠道是神经支配最密集的外周器官,拥有数以亿计的神经元细胞。至今已有许多研究表明肠道神经可以维持肠道内稳态或参与肠道疾病的发生,如肠道炎症与神经递质密切相关[7-9] 。其中一个关键信号分子是血清素(又称5-羟色胺,5-HT),它作为一种神经递质,在胃肠道中发挥作用,调节肠道中的广泛生理功能[9-10] 。人体内绝大多数5-HT(>90%)都是由肠嗜铬细胞产生,由限速酶色氨酸羟化酶(TPH) 1合成,小部分5-HT也由肌丛中的TPH2产生[9,11-12] 。同时,胃肠道管腔内含有高达数十万亿的微生物群,被称为人体的“第二基因”。大量研究表明,肠道菌群影响神经递质5-HT的合成,这与TPH1的表达密切相关[13] 。
本课题组前期研究已证明放射性肠炎与铁死亡密切相关,抑制铁死亡可一定程度地减轻肠组织损伤[14] 。铁死亡是一种铁依赖性调节性细胞死亡,由过度的脂质过氧化引起。与其他经典的细胞死亡方式相比,其标志性的特征是在细胞死亡过程中积累了大量的铁离子和脂质活性氧[15-18] 。其是否参与肠道神经对放射性肠损伤的调节尚未可知。因此,本研究拟探讨神经递质5-HT对放射性肠炎的影响及在此基础上对铁死亡潜在的调控机制。
1 材料与方法
1.1 实验动物和主要材料
20只6~8周龄、体重25 g左右的野生型C57BL/6小鼠购自江苏大学动物实验中心;10只 Tph1 敲除( Tph1 -/- )小鼠购自赛业生物公司。Trizol试剂(日本TaKaRa公司);苏木素和伊红染液购自上海碧云天有限公司;氨苄西林、万古霉素、链霉素、甲硝唑、氟康唑、Triton X-100和抗5-HT抗体(德国Merck公司);多聚甲醛购自镇江奥力化学公司;抗Ki67抗体和DAPI购自武汉赛维尔生物有限公司;丙二醛(MDA)和还原型谷胱甘肽(GSH)/氧化型谷胱甘肽(GSSG)ELISA试剂盒购自上海爱萌优宁生物技术有限公司;qRT-PCR试剂盒购自南京诺唯赞科技生物股份有限公司。
1.2 动物模型
所有小鼠在温度20~25 ℃和相对湿度40%~60%的环境中饲养,并自由地饮水以及摄食。将20只WT小 鼠按随机原则分为对照组(未照射)、照射组(单纯照射)、抗细菌+照射组和抗真菌+照射组,每组各5只。将10只 Tph1 -/- 小鼠按随机原则分为对照组和照射组,每组各5只。抗细菌+照射组小鼠连续喂食含有氨苄西林(0.5 mg/mL)、万古霉素(0.5 mg/mL)、链霉素(0.5 mg/mL)、甲硝唑(1 mg/mL) 的抗生素水,为期2周。抗真菌+照射组小鼠腹腔注射氟康唑(2 mg/mL)2周。照射处理3 d后,断颈处死小鼠,取小鼠肠组织进行后续实验。本研究通过江苏大学附属医院医学伦理委员会批准(伦理编号:KY2024K0602)。
1.3 照射方法
将小鼠麻醉后用胶布固定小鼠四肢于泡沫板上,然后用Varian 23EX直线加速器装置照射,照射剂量为14 Gy,剂量率为600 Mu/min,腹部接受辐射。
1.4 RNA测序
分别收集WT小鼠的对照组和照射组以及 Tph1 -/- 照射组小鼠的肠组织送至Novogene Co公司进行RNA测序,对获取的RNA测序数据进行分析。
1.5 qRT-PCR检测肠组织中 Tph1 "mRNA的相对表达水平
用Trizol提取总RNA后,使用PrimeScript RT-PCR试剂盒将提取的RNA逆转录合成cDNA。根据PrimerBank查找的基因序列设计引物, Tph1 上游引物为5′-AACAAAGACCATTCCTCCGAAAG-3′,下游引物为5′-TGTAACAGGCTCACATGATTCTC-3′。再使用SYBR Premix Ex "Taq 进行实时PCR,配制20 μL的PCR反 应体系:8.2 μL DEPC水、10 μL 2×AceQ qPCR SYBR Green Master Mix(无ROX)、上下游引物各0.4 μL、1.0 μL cDNA,混匀后置于PCR仪中。反应程序:预变性95 ℃ 5 min;循环反应95 ℃ "10 s,60 ℃ 30 s,共40个循环;熔解曲线95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。将 Tph1 "mRNA的表达量标准化为 GAPDH "mRNA水平。最后,应用2-ΔΔCt 方法计算相对mRNA表达。
1.6 免疫荧光检测肠组织中5-HT表达
肠组织经过10%多聚甲醛固定、石蜡包埋、切片和0.1%Triton X-100渗透等一系列程序。之后,将切片与5-HT抗体在4 ℃下孵育过夜,再加入荧光二抗(1∶200),在室温下的黑暗环境中孵育1 h。DAPI(1 μg/mL)在黑暗中孵育15 min。最后,用倒置荧光显微镜观察图像。
1.7 肠组织HE染色及评估
切除小鼠的小肠组织,并用冰冷的PBS清洗以去除粪便。用10%多聚甲醛固定后,将处理好的肠组织样品进行脱水、石蜡包埋、切片、脱蜡,HE染色。最后用显微镜观察组织。基于以下几个标准从组织学角度量化照射引起的肠黏膜损伤程度(0分=无,1分=轻度,2分=中度,3分=高度),包括血管扩张、隐窝损伤、炎性细胞浸润和上皮结构的维持[14] 。
1.8 免疫组化染色观察肠组织中Ki67的表达
10%多聚甲醛固定肠组织,然后石蜡包埋,切片。将切片与血清在室温下孵育1 h,然后与抗Ki67抗体(1∶300稀释)在4 ℃下孵育过夜。之后,将切片在37 ℃的二抗中孵育30 min。最后通过Image J软件对阳性染色进行分析定量。
1.9 肠组织中MDA和GSH/GSSG的检测
将适量肠组织用PBS漂洗干净,制成匀浆液,以3 500 r/min离心30 min后取上清液。取EP管分别加入标准品/样品稀释液(1×)100 μL,对倍稀释加入标准品,将配置/分装好的标准品及待测样品分别加入提取液100 μL,振荡混匀后,室温静置20 min 。再加入显色液400 μL,混匀后90 ℃水浴10 min。然后将反应好的样品及标准品,8 000 r/min离心1 min,取上清液,记录532 nm和412 nm处的光密度( D )值,最后根据样品 D 值计算出相应含量。
1.10 统计学分析
每个实验至少进行3次,应用GraphPad Prism 9软件进行统计学分析,定量数据用均值±标准差表示,两组比较采用两样本 t 检验,3组或3组以上均数比较采用单因素方差分析和Tukey法多重比较。 P lt;0.05为差异有统计学意义。
2 结果
2.1 照射促进WT小鼠肠组织中 Tph1 "mRNA和5-HT表达升高
通过对WT小鼠对照组和照射组的肠组织进行RNA测序并分析,结果显示,两组之间的转录谱存在明显差异(图1A)。接着筛选不同神经递质关键合成酶的表达水平,发现肠道中 Tph1 的表达在照射组显著上调;然而,其他神经递质合成酶,包括 Tph2 、酪氨酸羟化酶( Th )、苯乙醇胺N-甲基转移酶( Pnmt ) 、谷氨酸脱羧酶1( Gad1 )、 Gad2 和胆碱乙酰基转移酶( Chat ),在对照组和照射组肠组织中的表达水平没有明显差异(图1B)。qRT-PCR进一步证实 Tph1 "mRNA的表达在照射组WT小鼠肠组织中显著上调(图1C)。免疫荧光结果表明,照射组WT小鼠肠组织中的5-HT表达较对照组明显升高(图1D) 。这提示5-HT在照射所致的肠损伤中可能有潜在作用。
2.2 敲除 Tph1 减轻放射性肠炎
HE染色结果表明,与WT对照组小鼠相比, Tph1 -/- 对照组小鼠肠组织没有明显差异,无损伤表现,而WT照射组和 Tph1 -/- 照射组小鼠的肠组织均遭受了不同程度的损伤。其中WT照射组小鼠肠组织损伤明显加重,表现为肠隐窝紊乱和黏膜水肿, Tph1 -/- 照射组小鼠肠组织损伤则明显减轻(图2A)。Ki67免疫组化结果显示,在WT和 Tph1 -/- 小鼠中,照射组Ki67阳性细胞数量较对照组均明显减少,同时 Tph1 -/- 照射组的Ki67阳性细胞数量显著高于WT照射组(图2B)。这些结果说明敲除 Tph1 可减轻放射性肠损伤,提示5-HT可能在放射性肠炎发生过程中起到了一定促进作用。
2.3 敲除 Tph1 减轻照射后铁死亡
对照射后WT小鼠和照射后 Tph1 -/- 小鼠的肠组织进行RNA测序并比较两者的转录组变化。结果显示,两组的转录谱存在显著差异(图3A);与WT照射组相比, Tph1 -/- 照射组的促铁死亡基因表达明显下调,而抑铁死亡基因表达上调(图3B)。通过测定肠组织MDA含量和GSH/GSSG比值,进一步评估照射后小鼠肠组织铁死亡程度,结果表明,与WT照射组相比, Tph1 -/- 照射组的MDA含量明显降低,GSH/GSSG比值明显升高(图3C)。这些结果说明 Tph1 敲除可减轻照射后铁死亡,间接提示5-HT可能通过诱导铁死亡来促进放射性肠损伤。
2.4 抑制肠道细菌和真菌减轻放射性肠炎
将WT小鼠分别进行抗细菌和抗真菌处理后照射,免疫荧光染色观察5-HT表达,结果显示抗细菌+ 照射组和抗真菌+照射组小鼠肠组织中5-HT水平明显低于单纯照射组(图4A)。HE染色显示,与单纯照射组相比,抗细菌+照射组和抗真菌+照射组的肠组织损伤明显减轻(图4B)。Ki67染色结果表明,抗细菌+照射组和抗真菌+照射组的Ki67阳性细胞明显多于单纯照射组(图4C)。进一步测量肠组织MDA含量和GSH/GSSG比值发现,抗细菌+照射组和抗真菌+照射组肠组织MDA含量较单纯照射组明显降低,而GSH/GSSG比值明显升高(图4D)。这些结果表明抑制肠道菌群可减轻放射性肠道损伤。
3 讨论
放疗作为治疗癌症的常用手段,不仅对肿瘤细胞有杀伤作用,也对正常细胞造成直接或间接的损伤,导致各种并发症。胃肠道细胞对辐射敏感,放疗很容易引起放射性肠炎,影响患者的生活质量,甚至长期生存。目前,放射性肠炎无法通过药物有效治疗。由于肠道中含有大量神经,神经可以直接或间接地调节肠道的各种生理功能。此前已有多项研究证明肠道神经与肠道炎症密切相关[7,19-20] 。本研究中对照射组小鼠的肠道组织进行RNA测序,寻找可能的治疗靶点,观察到照射组WT小鼠肠组织中 Tph1 "mRNA的表达明显增加。TPH1在神经递质5-HT的生成中起限速酶的作用。采用免疫荧光检测证实照射后WT小鼠肠组织中5-HT的合成显著增加。这表明神经递质5-HT可能加重放射性肠炎。
肠道神经在肠道炎症的发生和发展中起着至关重要的作用,而神经递质在肠道炎症中的作用尚不清楚。根据目前的实验结果推测神经递质5-HT在放射性肠炎中起重要作用。本研究发现 Tph1 -/- 照射组小鼠肠道损伤较WT小鼠照射后明显减轻。同样对WT小鼠和 Tph1 -/- 小鼠照射后的肠组织进行RNA测序,发现两组中铁死亡相关基因的表达都发生了显著变化。MDA反映组织脂质过氧化损伤程度,GSH/GSSG比值是细胞内氧化还原状态的关键动态指标[21] 。检测肠组织中MDA、GSH/GSSG发现,在 Tph1 敲除后,照射引起的小鼠肠组织中的铁死亡被抑制,提示5-HT加重放射性肠炎,铁死亡途径或许在其中发挥一定作用。这些结果揭示了神经递质5-HT与放射性肠炎之间先前未被认识到的联系。
越来越多的研究证实,辐射暴露会极大地改变肠道微生物群,并导致微生物种群不平衡。氧化应激、炎症和组织损伤的增加均与这种生态失调有关[22] 。由于5-HT与肠道菌群的调节密切相关[13,23] ,推测肠道菌群可能通过影响5-HT加重放射性肠炎。本研究中将WT小鼠分为照射组(单纯照射)、抗细菌+ 照射组和抗真菌+照射组进行比较,结果表明肠道内的细菌和真菌可能通过促进5-HT水平升高加重放射性肠损伤。本研究结果表明靶向肠道微生物群5-HT轴可能减轻放射性肠炎。同时,通过微生物治疗来调节肠道微生物群落,或许为减轻放疗引起的肠组织损伤提供了一种治疗方向。本课题组前期研究已证明铁死亡途径在加重放射性肠炎机制中具有重要作用[14] ,抑制铁死亡可能有助于减轻放射性肠损伤。而近年来大量研究表明[24-25] ,铁死亡具有天然的抗肿瘤作用,放疗可诱导癌细胞铁死亡,增强放射治疗的疗效,从而抑制癌症进展。总而言之,放疗诱导的铁死亡在癌症治疗中仍然是一把双刃剑,值得进一步研究。
[参考文献]
[1] Xiao "HW, Cui M, Li Y, et al. Gut microbiota-derived indole 3-propionic acid protects against radiation toxicity via retaining acyl-CoA-binding protein[J]. Microbiome, 2020, 8(1): 69.
[2] Yang "Q, Qin B, Hou W, et al. Pathogenesis and therapy of radiation enteritis with gut microbiota[J]. Front Pharmacol, 2023, 14: 1116558.
[3] Zhang "T, Shi L, Li Y, et al. Polysaccharides extracted from rheum tanguticum ameliorate radiation-induced enteritis via activation of Nrf2/HO-1[J]. J Radiat Res, 2021, 62(1): 46-57.
[4] Li "M, Gu Y, Ma YC, et al. Krüppel-like factor 5 promotes epithelial proliferation and DNA damage repair in the intestine of irradiated mice[J]. Int J Biol Sci, 2015, 11(12): 1458-1468.
[5] Andreyev "J. Gastrointestinal complications of pelvic radiotherapy: are they of any importance?[J]. Gut, 2005, 54(8): 1051-1054.
[6] Bo "Z, Yongping S, Fengchao W, et al. Identification of differentially expressed proteins of γ-ray irradiated rat intestinal epithelial IEC-6 cells by two-dimensional gel electrophoresis and matrix-assisted laser desorption/ionisation-time of flight mass spectrometry[J]. Proteomics, 2005, 5(2): 426-432.
[7] Teratani "T, Mikami Y, Kanai T. Neuroimmune crosstalk in the gut and liver[J]. Int Immunol, 2022, 34(9): 475-484.
[8] Hatamnejad "MR, Baradaran Ghavami S, Shirvani M, et al. Selective serotonin reuptake inhibitors and inflammatory bowel disease; Beneficial or malpractice[J]. Front Immunol, 2022, 13: 980189.
[9] Gershon "MD, Tack J. The serotonin signaling system: from basic understanding to drug development for functional GI disorders[J]. Gastroenterology, 2007, 132(1): "397-414.
[10] Berger "M, Gray JA, Roth BL. The expanded biology of serotonin[J]. Annu Rev Med, 2009, 60: 355-366.
[11] Maffei "ME. 5-hydroxytryptophan (5-HTP): natural occurrence, analysis, biosynthesis, biotechnology, physiology and toxicology[J]. Int J Mol Sci, 2020, 22(1): 181.
[12] Zhu "P, Lu T, Chen Z, et al. 5-hydroxytryptamine produced by enteric serotonergic neurons initiates colorectal cancer stem cell self-renewal and tumorigenesis[J]. Neuron, 2022, 110(14): 2268-2282. e4.
[13] Yano "JM, Yu K, Donaldson GP, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis[J]. Cell, 2015, 161(2): 264-276.
[14] Ji "Q, Fu S, Zuo H, et al. ACSL4 is essential for radiation-induced intestinal injury by initiating ferroptosis[J]. Cell Death Discov, 2022, 8(1): 332.
[15] Tang "D, Kroemer G. Ferroptosis[J]. Curr Biol, 2020, 30(21): R1292-1297.
[16] Badgley "MA, Kremer DM, Maurer HC, et al. Cysteine depletion induces pancreatic tumor ferroptosis in mice[J]. Science, 2020, 368(6486): 85-89.
[17] Chen "Y, Zhang P, Chen W, et al. Ferroptosis mediated DSS-induced ulcerative colitis associated with Nrf2/HO-1 signaling pathway[J]. Immunol Lett, 2020, 225: 9-15.
[18] Probst "L, Dchert J, Schenk B, et al. Lipoxygenase inhibitors protect acute lymphoblastic leukemia cells from ferroptotic cell death[J]. Biochem Pharmacol, 2017, 140: 41-52.
[19] Zhang "W, Lyu M, Bessman NJ, et al. Gut-innervating nociceptors regulate the intestinal microbiota to promote tissue protection[J]. Cell, 2022, 185(22): 4170-4189. e20.
[20] Sharkey "KA, Mawe GM. The enteric nervous system[J]. Physiol Rev, 2023, 103(2): 1487-1564.
[21] Chen "X, Tsvetkov AS, Shen HM, et al. International consensus guidelines for the definition, detection, and interpretation of autophagy-dependent ferroptosis[J]. Autophagy, 2024, 20(6): 1213-1246.
[22] Wang "W, Cui B, Nie Y, et al. Radiation injury and gut microbiota-based treatment[J]. Protein Cell, 2024, 15(2): "83-97.
[23] Strandwitz "P. Neurotransmitter modulation by the gut microbiota[J]. Brain Res, 2018, 1693(Pt B): 128-133.
[24] 李 政, 刘雅雯, 陈家希, 等. hnRNPA2B1通过抑制转铁蛋白受体增强胰腺癌细胞对铁死亡的抵抗[J]. 江苏大学学报(医学版), 2024, 34(1): 1-10.
[25] Lei "G, Zhang Y, Koppula P, et al. The role of ferroptosis in ionizing radiation-induced cell death and tumor suppression[J]. Cell Res, 2020, 30(2): 146-162.
[收稿日期] 2024-04-07" [编辑] 何承志
