基质与灌溉耦合对大叶绣球光合特性和水分利用效率的影响
2024-12-31杨君邢强秦俊

关键词: 大叶绣球; 基质; 灌溉; 光合特性; 水分利用效率
亏缺灌溉(deficient irrigation,DI ) 是一种有效的节水方式,可以在减少耗水量的同时维持植物产量[1]和生长品质[2]。除了优化灌溉方式外,园林绿化废弃物(garden waste compost)、生物炭(biochar) 等基质改良剂的添加,在提高植物生物量,缓解缺水干旱等方面表现出良好的应用潜力,成为国内外学者研究的热点。园林废弃物指苗木自然凋落或养护过程中所产生的修剪物,通过好氧发酵堆肥形成,可有效增加基质有机质,提高基质肥力,提升苗木生长品质。生物炭是由有机物在无氧或低氧的条件下热解炭化形成的稳定富炭物质,施入基质后可改善基质理化性质,显著增加基质碳储量,促进植物生长,提高植物抗逆性[3];但生物炭性质(原料来源、热解温度)、施用量、基质质地、植物类型等均是影响生物炭效应的主要因素[4]。因此,在不同基质和水分条件下,生物炭产生的效果仍需进一步探索。在水分亏缺条件下,提高植物水分利用效率是实现农业节水的最终目标。水分利用效率可分为叶片水平和整株植物水平,利用气体交换法测定的叶片内在和瞬时水分利用效率仅可表征植物水分利用过程中的瞬时响应;而碳同位素分辨率(Δ13C) 可综合反映植株在较长时期生理过程中的气孔行为、光合特性和蒸腾作用的积累过程,成为指示植物长期水分利用效率的有效途径[5]。
大叶绣球(Hydrangea macrophylla) 作为世界范围内广受欢迎的花灌木,其叶片大而肥厚,生长旺季主要在春夏季节,对水分需求较高,限制了其在干旱和半干旱区的应用推广。近年来,随着基质水分传感器技术的发展(substrate moisture sensors ,SMS),针对特定植物实现精准灌溉成为可能。SMS 可实时监测基质水分状态,当达到预设阈值的基质含水率时,灌溉控制器会自动将含水率补充到预设水平,可显著减少灌溉用水,但在阈值设置方面需依据不同基质类型确定合适的灌溉上下限。目前这一技术已被应用于栎叶绣球(H. quercifolia ‘Alice’) 和大叶绣球(H. macrophylla ‘Fasan’)[6]的需水量评估上。BasiriJahromi 等 [ 7 ]以圆锥绣球(H. paniculata ‘SilverDollar’) 为试材,研究了不同配比基质和灌溉方式对其生长的影响,结果表明添加10% 的生物炭结合基于基质含水量的灌溉方式,可有效提升其水分利用效率。然而,大叶绣球对不同灌溉方式的生长、生理和水分利用的响应规律尚未完全明确,在不同基质环境下会更为复杂,还有待进一步研究和探索。因此,本研究以大叶绣球‘花手鞠’(H. macrophylla‘Hanatemari’)为研究对象,设置田间盆栽试验,通过搭建智能灌溉系统,研究不同基质类型和灌溉方式对大叶绣球光合特性和水分利用效率的影响,揭示大叶绣球在不同基质和灌溉措施下的响应规律,以期为大叶绣球科学灌溉和高效栽培提供理论依据。
1 材料与方法
1.1 试验设计
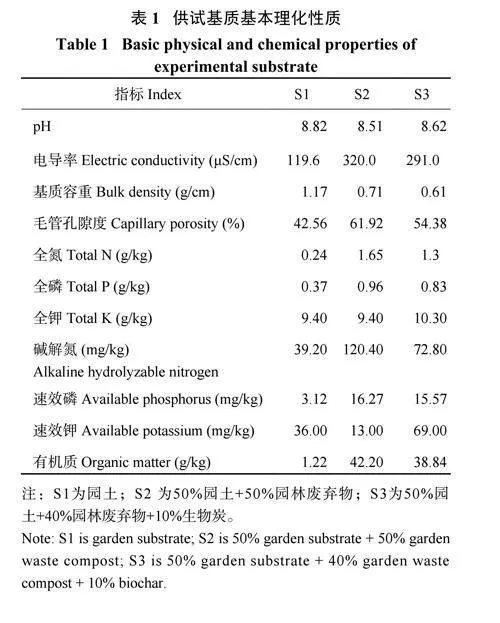
试验于2023 年7—9 月在上海辰山植物园行道树大棚中开展(31°08′N,121°18′E),配有遮雨棚,以防止自然降雨对试验的影响。场地位于上海市松江区,属于亚热带季风气候,大棚内的环境数据由温湿度记录仪(S21A,彭云物联) 自动记录(图1)。供试植物为2 年生大叶绣球品种‘ 花手鞠’ ( H .macrophylla ‘Hanatemari’)。供试基质为园土、园林绿化废弃物和生物炭。园土取自农田表层0—25cm 的沙壤土;园林绿化废弃物由植物园内日常修剪的树枝树叶自然堆肥腐熟形成;生物炭为竹炭,在600℃ 温度下热解形成,竹炭颗粒统一过2 mm 筛。试验开始前将不同配比的基质混匀后装填至体积为7.6 L 的盆中(盆栽容器上口直径23 cm,下口直径20 cm,高21.5 cm),装填基质约7 L 。
试验设置灌溉和基质两个因素,包括 3 种基质类型和两种灌溉方式,共6 个处理,每个处理8 次重复,完全随机区组排列。3 种基质分别为园土(S1)、50% 园土+50% 园林废弃物(S2) 和50% 园土+40% 园林废弃物+10% 生物炭(S3),基本理化性质见表1。两种灌溉方式分别为充分灌溉(FI) 和亏缺灌溉(DI)。灌溉系统采用基于云平台的智能灌溉控制系统,由上海润绿灌溉有限公司设计搭建,由电磁阀、基质水分传感器、流量传感器、数据采集终端等组成,可实现灌溉水量和灌溉上下限的精准控制。灌溉启动和关闭由电磁阀(EZP-23-54,托罗) 控制,水量由流量传感器(485,润绿) 监测,基质体积含水率由基质水分传感器(TDR,润绿) 实时监测。FI 处理通过喷灌方式每日定时灌溉固定水量以实现充分灌溉,喷头流量为80~280 L/h,工作压力150~300 Mpa,每日总灌溉量为2.8 mm。DI 处理通过滴灌方式实现亏缺灌溉,滴头流量2 L/h,工作压力100 Kpa;依据基质最大持水能力的不同设置灌溉上下限,通过基质水分传感器(量程为0%~100%,精度为±3%)实时监测基质体积含水率变化,每隔1 h 采集数据1 次。DI 处理开始前参考前人的研究确定不同基质的最大持水能力[8−9],具体方法为:将装满土的盆钵放置在有湿润和松散的基质表面,通过底部尼龙网孔与花盆内的基质建立水力接触,然后灌溉所有盆钵使基质达到饱和,用基质水分传感器实时监测体积含水率,待基质中的重力水完全下渗,基质体积含水率几乎不变达到稳定,此时的体积含水率即为盆内的最大基质持水能力,测得的S1、S2 和S3 基质的最大持水能力分别为36.8%、49.1% 和41.7%。灌溉上下限设定为当基质体积含水率下降至基质最大持水能力的40% 时启动,当基质体积含水率上升到基质最大持水能力的95% 时则停止[10] (表2)。试验开始前,将基质体积含水率按不同基质类型统一灌溉至 95% 最大持水能力。
1.2 测定项目和方法
试验开始前选取长势一致的2 年生大叶绣球种苗移栽至盆中,每盆定植1 株。在试验开始和结束时,用钢卷尺(精确到1 mm) 测量大叶绣球盆栽苗的株高和冠幅。盆栽基质表面至植株最高点的自然高度为株高;测量盆栽分枝间的最大宽度记为冠幅1,垂直于冠幅1 的宽度记为冠幅2。植物生长指数 =(冠幅1+冠幅2+株高)/3,通过试验期间植物生长指数的增量/每盆总耗水量计算植株水分利用效率[11]。灌溉处理结束后获取大叶绣球地上组织,每个处理选取生长一致的植株3 株,清洗后放入烘箱105℃杀青0.5 h,70℃ 烘干至恒重,再在干燥玻璃器皿内冷却至室温后称量地上生物量。
在8 月晴朗无风的天气,于上午9:00—11:00用便携式光合仪(Li-6400,LI-COR) 测定叶片气体交换参数和光合-CO2 响应曲线。选取生长状况一致的植株,测定相同叶位且完全展开的成熟叶片的净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatalconductance,Gs)、胞间CO2 浓度(intercellular CO2concentration,Ci) 和蒸腾速率(transpiration rate,Tr),并计算叶片内在水分利用效率(intrinsic wateruse efficiency,WUEn,WUEn =Pn /Gs) 和瞬时水分利用效率(instantaneous water use efficiency,WUEi,WUEi =Pn /Tr)。光合−CO2 响应曲线选取相同叶片,测定条件设置为: 叶室温度 25°C,空气流速 500μmol/s,光合有效辐射1000 μmol/(m2·s),叶室 CO2浓度由安装的高压浓缩小钢瓶控制,CO2 浓度梯度先降低后升高,设定的CO2 浓度梯度为400、300、200、100、50、0、400、400、600、800、1000、1200、1500、1800、2000 μmol/mol,共15 个浓度。获取响应曲线后,使用叶子飘光合计算软件的直角双曲线模型进行拟合[12]。在光合−CO2 响应曲线测定完成后选用相同叶片使用叶绿素仪(TYS-3N,托普) 测定相对叶绿素含量SPAD 值。
当亏缺灌溉下基质体积含水率降至最低点时,各处理取每盆光合测定的目标叶,用PSYPRO 露点水势仪测定叶片午间叶水势。测定完成后用印记法测定叶片气孔形态,先用透明指甲油获取叶片下表皮印记,随后用镊子将表皮印记转移至显微镜载玻片上,用Axio Scope A1 显微镜在40×镜下观察并获取图像,随机获取 6 张图像后用 ImageJ 软件计数所有气孔,计算单位面积的气孔即气孔密度(SD,stomataldensity);并测量气孔长(stomatal length, SL) 和气孔宽(stomatal width,SW),计算气孔大小(SS,stomatalsize),公式为SS = (π×SL×SW)/4[13]。
光合测定完成后每处理选取6~8 片成熟叶片测定叶片碳同位素自然丰度。叶片中的δ13C 含量采用稳定同位素质谱仪(Delta plus XP,赛默飞) 测定,并根据公式将δ13C 转换为碳同位素分辨率Δ13C:
1.3 数据分析方法
试验所得数据用Excel 整理分析后,采用 SPSS进行不同处理间单因素方差分析、双因素交互作用分析和Duncan 多重比较,GraphPad Prism 制图;使用R 语言Psych 包进行Pearson 相关性分析及制图,主成分分析使用标准主成分法,通过FactoMineR 包进行分析和制图。图表中数据除标注外均为平均值±标准差。
2 结果与分析
2.1 基质与灌溉对基质水分状况的影响
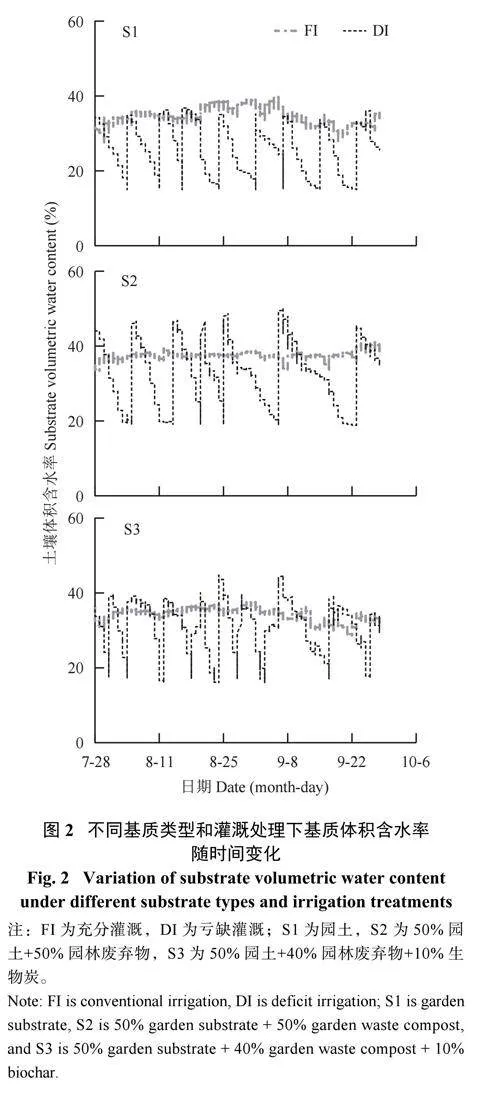
不同基质类型的基质体积含水率在充分灌溉(FI) 条件下保持相对平稳的变化,在亏缺灌溉(DI)时存在明显差异(图2)。S1 在整个DI 灌溉周期经历了8 个干湿循环,第一个最低点出现在灌溉处理后的第8 天(8 月4 日);S2 则经历了6 个干湿循环,第一个最低点出现在灌溉处理后第9 天(8 月5 日);S3 则经历了9 个干湿循环,第一个最低点出现在灌溉处理后第4 天(7 月31 日)。由此可见,亏缺灌溉的基质体积含水率根据设定的阈值变化,当基质含水率逐渐降至低阈值后复水升高,依次循环。不同基质的灌溉频率存在差异,S2 的灌溉频率最低,并且在夏季(7—8 月) 的灌溉频率高于秋季(9 月)。
2.2 基质与灌溉对大叶绣球叶片光合特性的影响
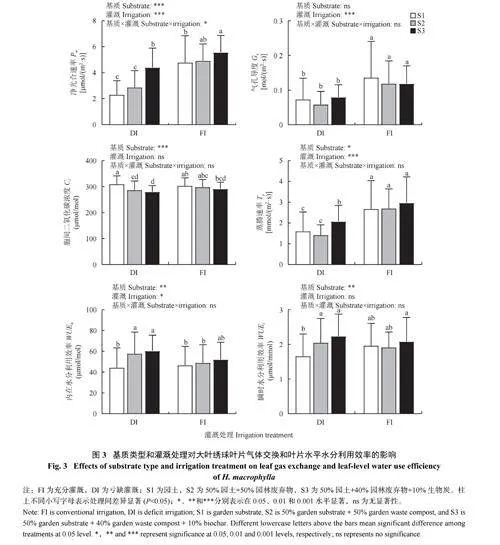
2.2.1 基质与灌溉对大叶绣球叶片气体交换参数的影响 不同基质类型显著影响了大叶绣球叶片的Pn、Ci、Tr、WUEn 和WUEi;与S1 相比,S2 和S3 的Pn 分别显著增加了10.0% 和40.9%;Ci 分别显著降低了4.5% 和6.7%;WUEn 分别显著增加了17.5% 和24.1%;WUEi 分别显著增加了9.4% 和19.4%。不同灌溉类型显著影响了大叶绣球叶片的Pn、Gs、Tr、WUEn;与 FI 相比,DI 显著降低了Pn(37.3%)、Gs (43.4%) 和Tr (38.9%),显著增加了WUEn (10.2%)。此外,叶片的净光合速率显著响应灌溉×基质的交互作用,在DI 条件下,与S1 相比,S3 大叶绣球叶片的Pn 显著提高了91.4% (图3)。
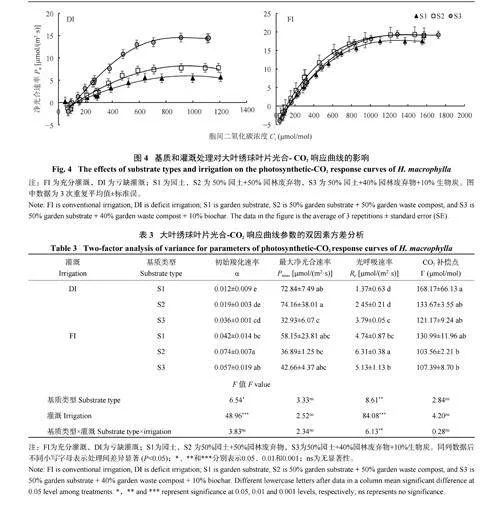
2.2.2 基质与灌溉对大叶绣球叶片光合-CO2 响应曲线的影响 由图4 可知,在低CO2 浓度条件下,净光合速率随着胞间CO2 浓度增加快速上升,随着CO2 的进一步升高,净光合速率呈线性快速上升,直至净光合速率稳定在一定水平。与S1 相比,S2和S3 处理明显放大了不同基质类型的光合-CO2 响应曲线的差异,尤其在亏缺灌溉(DI) 中更为明显。表3通过直角双曲线模型拟合得出了大叶绣球光合-CO2 响应曲线的重要参数。双因素方差分析表明基质和灌溉处理显著影响了大叶绣球叶片的羧化效率α 和光呼吸速率Rp,其中Rp 显著响应了灌溉×基质的交互作用。与FI 相比,DI 的α 值和Rp 值分别显著降低了61.4% 和53.0%。在DI 条件下,3 种基质类型的α 值和Rp 值均表现为S1
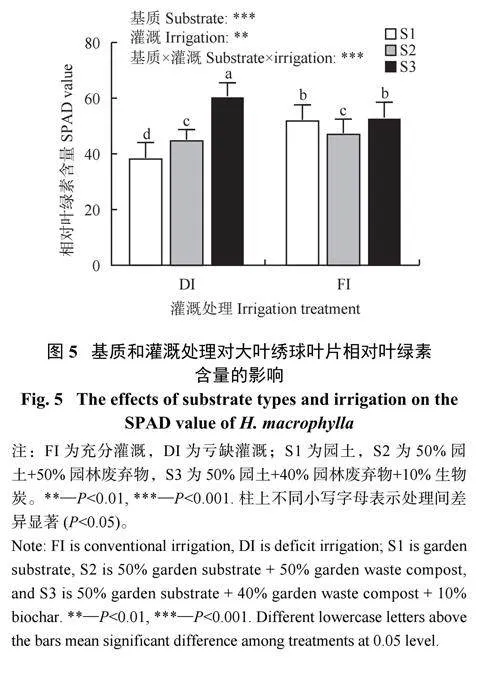
2.2.3 基质与灌溉对大叶绣球叶片SPAD 的影响
不同基质类型和灌溉方式显著影响了大叶绣球的SPAD 值。与FI 相比,DI 条件下SPAD 显著下降了5.5%。3 种基质类型中S3 处理的SPAD 值在不同灌溉条件下均为最大值。在DI 条件下,与S1 相比,S3 的SPAD 值显著上升了57.5% (图5)。
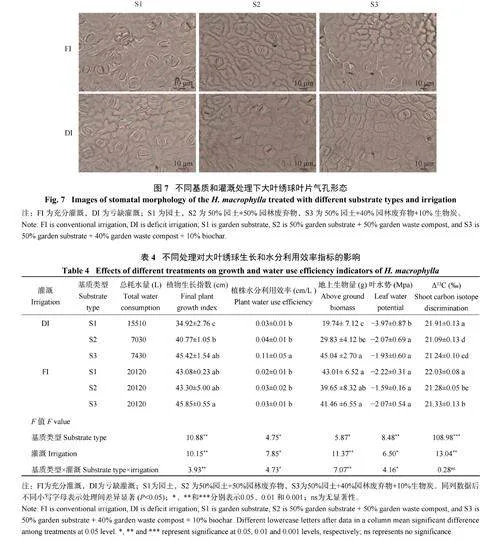
2.3 基质与灌溉对大叶绣球叶片气孔形态的影响
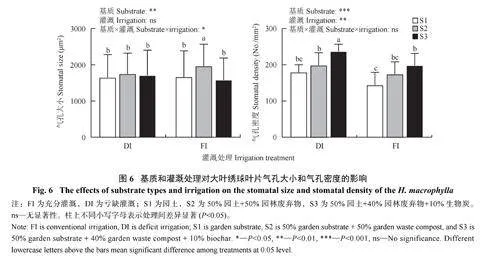
由图6、图7 可知,不同基质类型显著影响了大叶绣球叶片气孔大小,且叶片气孔大小显著响应基质×灌溉的交互作用。不同基质类型下叶片气孔大小表现为S3
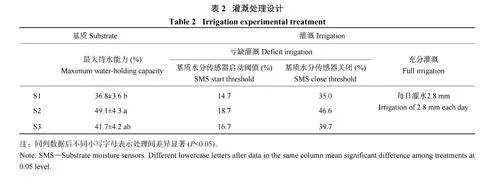
2.4 基质与灌溉对大叶绣球生长和水分利用效率的影响
DI 条件下,不同基质处理在整个灌溉周期的总耗水量表现为S1gt;S3gt;S2;与FI 相比,S1、S2 和S3 分别节约用水22.9%、65.1%、63.1%。双因素方差分析结果表明基质和灌溉显著影响了大叶绣球生长。在DI 条件下,不同基质处理的植物生长指数和地上生物量均表现为S1
不同灌溉和基质显著影响了大叶绣球植株水分利用效率。与FI 相比,DI 条件下的大叶绣球植株水分利用效率显著增加了1.2 倍,其中S3 基质在DI 下表现出最高的植株水分利用效率。不同基质和灌溉处理显著影响了大叶绣球叶水势。与FI 相比,DI 使得S1 基质的叶水势显著降低了78.7%,而S2 和S3基质的叶水势在不同灌溉条件下差异不显著,这表明园林废弃物和生物炭的添加一定程度上维持了亏缺灌溉条件下的叶水势。此外,DI 条件下不同基质类型的叶水势发生了显著变化,与S1 相比,S2 和S3基质的叶水势分别显著增加了47.9% 和51.4%。不同基质和灌溉方式对大叶绣球叶片碳同位素(Δ13C) 影响显著,但二者的交互作用影响不显著。3 种基质处理在不同灌溉条件下的叶片Δ1 3C 值均表现为S1gt;S3gt;S2,S2 与S3 较S1 的Δ13C 值均显著下降。与FI相比,同一基质处理下DI 具有更低的Δ13C 值,其中S2、S3 显著降低了95.7%、42.5% (表4)。
2.5 相关性分析和主成分分析
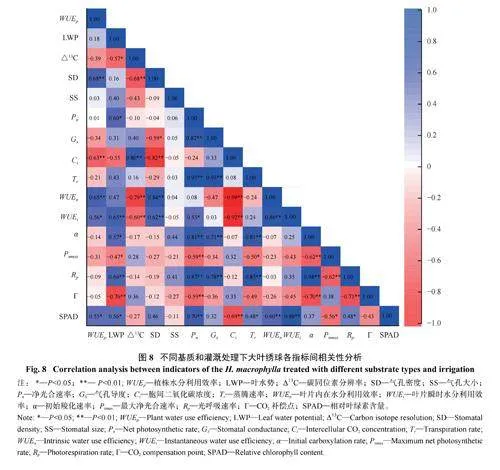
对测定的16 个指标进行相关性分析,发现气孔形态、光合和水分利用效率相关指标之间存在相关关系。其中,SD 与WUEp、WUEn 和WUEi 极显著正相关,与Ci 和Δ13C 极显著负相关;此外,Δ13C 与WUEn、WUEi 极显著负相关,相关系数分别为−0.79和−0.60,且与WUEn 的相关性高于WUEi (图8)。
对测定的16 个指标进行主成分分析,提取前2 个主成分,累计方差贡献为74.5%,可代表大部分的原始数据。由图9 可知,第1 主成分(PC1) 包含了原始信息的42.9%,主要由Rp 、Pn 和α 等光合相关指标共同决定,其Rp 在PC1 的投影较大,在PC2 投影较小,表明其对PC1 的贡献较大;第2 主成分(PC2) 包含了原始信息的31.6%,主要由WUEn、SD和WUEp 等水分利用效率指标共同决定,其中WUEn在PC2 的投影较大,在PC1 投影较小,表明其对PC2的贡献较大。主成分分析有效区分了不同灌溉处理和基质处理。不同基质处理沿PC1 轴分离,由左至右依次为S1,S2 和S3;与S1 相比,添加园林废弃物(S2) 和生物炭(S3) 后整体沿PC1 轴向右移动(图9-a)。亏缺灌溉(DI) 处理主要集中于第三和第四象限,相对于P C 2 轴呈负向分布;而充分灌溉(FI) 主要集中于第一、二象限,相对于PC2 轴呈正向分布(图9-b)。
3 讨论
水分是调控植物生长的主要因子,基质水分的亏缺会诱发一系列植物光合生理特性改变,使植物适应水分胁迫[14]。亏缺灌溉(DI) 在节水的同时,对植物产生的水分限制会降低光合速率,抑制Rubisco酶活性,导致植物碳同化能力降低[15 − 16]。本研究发现DI 处理下,大叶绣球叶片的净光合速率Pn、气孔导度Gs 和蒸腾速率Tr 均显著降低;进一步分析灌溉对大叶绣球CO2 响应曲线的影响发现,DI 的初始羧化速率α 值和光呼吸速率Rp 值均显著降低,这与前人[17 − 18]的研究结果基本一致。一般认为,α 值与Rubisco 酶活性呈正相关,其值减小反映了Rubisco酶的量及活性降低, 从而导致大叶绣球叶片对CO2 的利用效率降低;而Rp 值的下降可能是水分胁迫限制了大叶绣球的光合作用,植物通过抑制呼吸作用减少对光合产物的消耗[19]。此外,与其他两种基质相比,生物炭的添加显著提高了大叶绣球在亏缺灌溉条件下叶片的Pn、α、Rp、WUEi 和WUEn,主成分分析也表明Rp、Pn 和α 在光合相关指标中起主导作用;这可能由于施入生物炭后,缓解了水分亏缺对于光合作用的限制,提高了叶片水分利用效率,一定程度上维持了大叶绣球叶片净光合速率,提高了叶片对 CO2 的利用能力,并通过消耗过剩光能减轻水分胁迫对其的伤害,有利于保护光合机构并维持较高的光合速率。此外,大叶绣球的SPAD 值与光合参数表现出相似的趋势,即与其他两种基质相比,生物炭的添加显著提高了大叶绣球在亏缺灌溉条件下叶片的SPAD 值,缓解了水分亏缺条件下叶绿素含量的下降。
随着基质水分状况的改变,气孔密度和大小会发生可塑性形态变化。亏缺灌溉显著增加了大叶绣球叶片的气孔密度,但并未引起气孔大小的显著变化;这与干旱胁迫会导致玉米[20]、小麦[21]等植物气孔密度增加一致;但本研究并未发现水分亏缺导致气孔明显变小,这可能与植物种类的差异和水分胁迫程度有关[14]。本研究发现亏缺灌溉显著降低了大叶绣球的叶水势,表明灌溉量的减少影响了大叶绣球体内的水分状况,影响了叶片膨压及其活力,加剧了叶片的水分胁迫[22];但是园林废弃物和生物炭的添加,显著提高了亏缺灌溉条件下大叶绣球叶片的叶水势,表明这两种基质的施入一定程度上缓解了水分亏缺对大叶绣球水分状况的不利影响。植物碳同位素分辨率(Δ13C) 与稳定碳同位素组成 (δ13C) 一般为负相关,可表征植物长期的水分利用效率,一定程度上弥补了以气体交换为基础的水分利用效率评价的局限性[23]。本研究发现,与FI 相比,同一基质处理下亏缺灌溉具有更低的Δ13C 值,提高了大叶绣球长期水分利用效率;这是由于水分胁迫是通过影响植物气孔和Rubisco 酶,产生碳同位素分馏效应来改变 δ13C,已在许多研究中得到证明[24];同时与园土相比,园林废弃物和生物炭的添加也使得Δ13C 显著下降;相关性分析表明Δ13C 与气孔密度、WUEn 和WUEi呈极显著的负相关关系,这与魏镇华等[5]对不同灌溉条件下番茄Δ13C 与叶片水平水分利用效率关系的研究结果相似,即Δ13C 可作为量化表征大叶绣球叶片水分利用效率的重要指标;同时也表明在基质和灌溉的共同作用下,气孔形态的重塑可能也会影响大叶绣球的Δ13C,进而调节水分利用效率。相关性和主成分分析结果表明大叶绣球叶片的Δ13C 与WUEn的相关性明显优于WUEi,且WUEn 在水分利用效率相关指标中起主导作用,这与姜浩等[25]的结论一致,这是因为WUEn 能更直接反映气孔阻力对叶片胞间与大气CO2 浓度比(Ci/Ca) 的影响,从而导致Δ13C 对WUEn 的指示性优于WUEi。
一般而言,水分胁迫条件下,植物生产力会下降。本研究结果表明,在亏缺灌溉条件下,园林废弃物和生物炭的添加使得大叶绣球在降低耗水量的同时维持了较高的地上生物量和植物生长指数,这可能由于亏缺灌溉下形成的干湿交替环境,通过优化气孔形态,改善了植物营养状况,提高了大叶绣球碳同化能力[26];并且园林废弃物与生物炭改良基质后,促进了植物对水分、养分的吸收、利用和分配[27]。但这种影响也会因基质类型、植物种类存在差异[28],对植物的水分吸收也可能产生负面影响[29]。在相同灌溉条件下,生物炭的添加虽然显著提高了大叶绣球叶片的净光合速率,但蒸腾速率也显著增加,这一定程度上造成了水分无效损失。尽管这样,亏缺灌溉条件下生物炭的添加可以显著提高大叶绣球植株水分利用效率,一方面这可能由于生物炭和亏缺灌溉的结合在降低植物耗水量的同时维持了较高的地上生物量和植物生长指数;另一方面,二者通过协同作用调节大叶绣球生长和生理特性,如提高净光合速率,增加气孔密度,增加叶水势、降低叶片 Δ13C 等提高水分利用效率[14]。因此,生物炭结合亏缺灌溉可缓解水分限制的负面影响,在维持大叶绣球生长的同时提高了水分利用效率。
4 结论
亏缺灌溉处理下,大叶绣球叶片的净光合速率、气孔导度、蒸腾速率、初始羧化速率、光呼吸速率和相对叶绿素含量均显著降低,但生物炭的添加一定程度上维持了大叶绣球叶片净光合速率、初始羧化速率、相对叶绿素含量和光呼吸速率,提高了大叶绣球叶片对 CO2 的利用能力。
亏缺灌溉显著增加了大叶绣球叶片气孔密度,显著降低了叶水势和Δ13C。但园林废弃物和生物炭的添加使得大叶绣球在降低耗水量的同时维持了较高的地上生物量,尤其是生物炭的添加显著提高了大叶绣球水分利用效率。因此,生物炭和亏缺灌溉的联合应用是提高大叶绣球水分利用效率适宜的栽培组合。
