水稻减氮密植研究进展与“极限密植”技术
2024-12-31吴大霞陈世勇袁先福马超张亚丽夏金林汪建飞

关键词: 水稻; 产量; 氮素利用效率; 氮水平; 栽培密度; 极限密植
水稻是我国重要的粮食作物之一,提高单位面积水稻产量是确保国家粮食安全的关键。现行的水稻高产栽培技术多以重施氮肥作为基础。氮肥施用是获得单位面积理想作物群体生产力的重要保证,但同时带来巨大的经济成本和环境压力[1]。根据国家“稳粮增收、提质增效”的发展要务,提高水稻产量和氮素利用效率是水稻栽培发展上的两项重要任务。
氮素营养和栽培密度是影响水稻生长的两个主要因素。在传统稀植栽培模式下,农民普遍通过增施以氮肥为主的基蘖肥提高分蘖数,进而提高水稻产量。对水稻需肥规律的研究发现,拔节期至齐穗期是水稻吸收氮素高峰期,前期将大量氮肥施入土壤中造成田间氮素损失严重。林晶晶等[2]应用15N 示踪方法研究了不同时期水稻氮肥利用率,结果表明基蘖肥的吸收利用率均较低,不足30%。究其原因,主要是土壤供氮量和水稻氮素的吸收量不协调,制约了氮素利用效率和产量的提高。近年来,诸多研究发现,减氮密植栽培模式下水稻、小麦、玉米等作物在产量和氮肥利用效率的协同上存在显著优势,可达到“减氮增效”的目的[3−5]。但如何协调氮肥用量与种植密度二者之间的平衡,进而最大限度挖掘水稻减氮增效的潜力,目前还缺少系统化深入研究。
徐新朋等[6]研究发现,水稻密植提高了移栽后群体质量,显著增强了植株吸收养分的能力,进而提高氮肥利用率。赵杰等[7]提出以主茎成穗为主的“独秆”水稻栽培模式,基本苗为380×104 株/hm2,结合“基肥不施氮肥配合全程氮肥在分蘖中后期施用”的氮肥运筹方式可达到提质增产的目的。在此基础上,陈世勇等[8]总结出一套水稻“极限密植”栽培技术,即将达到水稻高产需要的有效穗数确定为本田栽插的秧苗数,以‘南粳46’为例,本田栽插的秧苗数为400×104 株/hm2,结合氮肥运筹,免施分蘖肥,控制分蘖发生,主要依靠主茎成穗形成水稻产量。“极限密植”的试验小区氮肥农学利用效率(NAE) 达29.9~35.7 kg/kg,氮肥利用率显著提高。本文通过综述氮素营养和栽培密度对水稻分蘖及光合特性的影响,进一步探讨氮密互作对水稻产量及氮素利用效率的影响,重点阐述了“极限密植”栽培技术对水稻生产的重要意义,旨在进一步丰富与优化氮密互作调控水稻产量理论的同时,为水稻减氮增效种植技术提供新思路。
1 氮素营养对水稻农艺性状的影响
1.1 氮素营养对水稻分蘖特性的影响
1.1.1 水稻分蘖的生理特征 分蘖是禾本科作物株型的关键组成部分,对水稻群体产量起决定性作用。水稻节间上存在分蘖原基,原基激活后形成分蘖芽,新生分蘖芽受到植物内部或外界信号刺激可触发其伸长,形成分蘖[9]。分蘖的形成可分为两个阶段,即分蘖芽的形成和分蘖芽的伸长。除自身遗传因素外,营养条件、温度、水分、光照等环境因子对分蘖有重要影响。
水稻分蘖与品种的自身特性密切相关,不同水稻品种和基因型之间分蘖能力差异很大,直接影响水稻产量和氮素利用效率[10−12]。适宜的温度有利于水稻分蘖生长。分蘖发生的最适宜温度为30℃~32℃,温度过低或过高均可抑制分蘖,日均温度在15℃ 以下不利于分蘖的发生,气温超过38℃~40℃ 分蘖也会停止。低温主要导致水稻根系活力下降,水分和养分吸收不足,植株氮代谢活性减弱,分蘖生长受到抑制[13]。光照对水稻分蘖的影响主要通过调节光合作用实现。光照充足时水稻分蘖生长迅速,光照时间和光强不足均可影响光合速率,导致同化产物减少,抑制水稻分蘖生长[14]。土壤水分也是影响水稻分蘖的重要环境因素。分蘖发生需要充足的水分,在土壤水分不足时,水稻生理活动减弱,养分供应不足,营养物质无法满足分蘖芽的生长需求,导致分蘖茎细弱或者凋亡。生产中可通过控制稻田灌水深度调节水稻分蘖发生。研究表明,相较于深灌,浅水灌溉下水稻低分蘖节位处光照和温度较高,分蘖数显著增加[15]。水分过多则会导致水稻茎基部氧气与光照不足,抑制分蘖的发生。水稻分蘖受到多种营养元素的调控。土壤中的氮、磷、钾、硼、锌等养分均能影响水稻分蘖的生长[13],其中氮素对分蘖生长的影响最为显著,氮素不能激活水稻分蘖原基,而是为腋芽的伸长提供营养物质[16]。
1.1.2 氮素营养影响水稻分蘖的机理研究 自水稻“叶蘖同伸”规律提出后,水稻分蘖与叶片含氮量的关系可作为氮素营养诊断的依据。水稻分蘖的生长对氮素供应十分敏感,供氮量的小幅差异会引起分蘖的显著变化[17−18]。研究发现,过高或过低的供氮水平均不利于分蘖产生。当水稻叶片含氮量约2.5% 时,植株无法生成新的分蘖;叶片含氮量不足1.5% 时,部分分蘖开始消亡;当叶片含氮量大于3.5% 时,分蘖迅速产生,但当含氮量超过一定值时,分蘖生长受到抑制[19]。丁艳锋等[20]指出分蘖及新叶的产生与供氮水平及叶片、叶鞘的含氮量密切相关。在水培条件下,营养液中氮素浓度在10~70 mg/L 范围时,分蘖的生长与氮浓度呈显著正相关,当氮素浓度超过70 mg/L 后,分蘖的生长则会被抑制。当水稻处于三幼一基期,分蘖发生的条件必须满足营养液氮素供应浓度高于30 mg/L,表明该时期30 mg/L 是分蘖产生的氮临界浓度。潘寿等[21]发现,水稻品种“日本晴”分蘖数随供氮浓度的增加而增多,当供氮浓度增加至70 mg/L 时,分蘖数增加不显著。氮水平为0、90、180、270、360 kg/hm2 5 个梯度的田间试验结果显示,水稻的分蘖数随氮水平的增加而增多,但增幅逐渐降低,有效分蘖数呈先增后减的趋势[22]。
生产中除了氮肥施用量,氮肥的施用时期、不同阶段的施肥配比均可影响水稻的有效分蘖数[ 2 3 ]。随着水稻“三控”施肥技术的应用和推广,生产上可通过“控肥”,减少分蘖肥的施用量,推迟分蘖肥的施用时期,前氮后移,达到“控苗”、“控病虫”的目的[24−25]。虽然单位面积穗数是产量构成要素中最重要的组成部分,传统栽培模式下水稻穗数在很大程度上受制于分蘖数目,但分蘖数目并不是越多越好。通过调控水稻生长不同时期的氮素供应水平,适当控制水稻分蘖数目,可优化水稻株型,促进水稻栽培的高产高效。
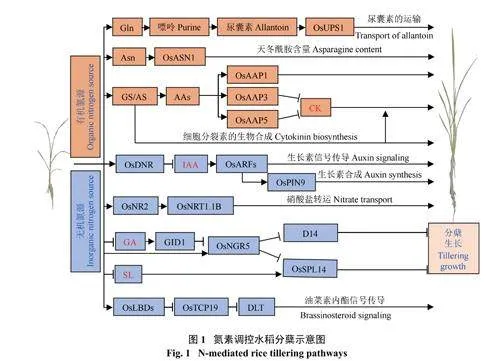
近年来氮素调控水稻分蘖的机理(图1) 已被部分阐明。有机氮源和无机氮源两种氮素形态均可影响水稻分蘖生长。谷氨酰胺(glutamine,Glu) 作为嘌呤合成的底物,可促进尿囊素的合成。尿素渗透酶基因OsUPS1 过表达可影响尿囊素从根向芽部的运输,分蘖数显著增加[26]。水稻天冬酰胺合成酶基因OsASN1 负责根部天冬酰胺(asparagine,Asn) 的合成,osasn1 突变植株中Asn 含量降低、分蘖芽伸长受抑制[27]。谷氨酰胺合成酶基因OsGS1;2 可通过调节氮依赖的细胞分裂素的生物合成来促进分蘖芽的生长[28]。另外,氨基酸渗透酶(amino acid permease,AAP) 是一类氨基酸转运蛋白,其中OsAAP1 主要负责转运中性和酸性氨基酸,OsAAP1 过表达茎秆中的中性和酸性氨基酸含量增加,碱性氨基酸含量降低,水稻分蘖数目增加[29]。OsAAP3 主要吸收转运碱性氨基酸,osaap3 突变体植株分蘖数显著增加,可能由于植株体内细胞分裂素( C K ) 含量提高[ 3 0 ]。OsAAP5 主要转运碱性和中性氨基酸,OsAAP5 过表达植株分蘖数显著降低,而osaap5 突变植物体内CK 含量增加,分蘖数目增多[31]。
OsDNR1 (DULL NITROGEN RESPONSE1) 编码氨基转移酶,可抑制生长素合成。osdnr1 突变后导致生长素在植株体内大量积累并激活生长素响应因子OsARF(AUXIN RESPONSE FACTOR),OsARF 可调控下游氮代谢相关基因,从而抑制分蘖生长[11]。且大量研究证明,氮素对分蘖的调控依赖于植物内源性激素。OsPIN9 是一种生长素外排载体,与硝态氮处理相比,OsPIN9 受到铵态氮的诱导表达,可调控水稻分蘖芽生长[32]。OsNR2 编码硝酸还原酶(NR),籼稻和粳稻中OsNR2 等位基因编码结构不同的OsNR2蛋白,其中籼稻OsNR2 表现出更大的NR 活性,影响硝酸盐的摄取,且O s N R 2 与硝酸盐转运蛋白OsNRT1.1B 存在相互作用,籼型OsNR2 等位基因可促进OsNRT1.1B 基因的表达,提高籼稻的有效分蘖数、产量和氮素利用效率(NUE)[ 3 3 ]。NGR5 (Nmediatedtiller growth response 5) 为水稻分蘖正调控因子。有研究表明,NGR5 是赤霉素(gibberellin acid,GA) 受体GID1 的靶基因,NGR5 的丰度与GA 水平呈负相关,NGR5 抑制分蘖抑制基因D14 (Dwarf14)和OsSPL14 (Squamosa Promoter Binding Protein Like-14)的表达,影响水稻分蘖[34]。另有研究表明,提高水稻硝酸盐处理浓度,可促进细胞分裂素合成基因的表达,细胞分裂素(cytokinin,CK) 积累增多,影响水稻分蘖[35]。高浓度的氮素营养还可通过抑制独脚金内酯(strigolactone,SL) 和生长素(auxin,IAA) 的合成,调控水稻分蘖[36]。此外,OsLBDs-OsTCP19-DLT模块与氮素引起的水稻油菜素内酯(brassinosteroid,BR) 等信号反应密切相关,可促进水稻分蘖[37](图1)。
1.2 氮素营养对水稻光合特性的影响
水稻籽粒中物质的积累主要依赖于叶片的光合作用,光合作用产生的碳水化合物,为植物在各种生物合成过程中提供能量和碳骨架。水稻冠层叶面积大小、叶片氮含量及叶片的形态特征与群体光合作用密切相关[38]。叶绿素及Rubisco 酶在光合作用中起着重要作用。研究表明,叶片含氮量与叶绿素和Rubisco 酶的含量呈显著正相关关系[39]。在一定范围内可通过提高叶片含氮量促进叶片光合速率[40]。另外,氮素可促进细胞扩增,提高叶片气孔导度和叶肉导度,促进光合进程[41]。吕魏等[42]研究发现,不同氮形态对叶绿素含量影响不同,相较于NO3− -N,NH4+-N 处理下水稻叶片叶绿素含量更高。
水稻生长前期氮肥的施用可促进水稻群体出叶速率,显著增加叶面积指数[43−44]。但水稻生育前期氮肥施用量过高,导致水稻生长中后期叶面积指数过大,叶片互相遮荫,下部叶片不断凋亡,导致抽穗期叶面积指数下降,影响光合产物的形成[45]。另外,徐敏等[46]研究表明,提高施氮水平还可提高抽穗期群体及成熟期群体的总叶面积指数,但持续提高施氮水平则会降低总叶面积指数。因此,合理的氮肥用量和施用时期可以促进水稻叶片的生长及生育后期干物质积累。适量氮肥的后移,既可提高水稻生长前期的群体质量,又能促进中后期群体的光合效率,减缓叶片衰老,促进氮素吸收,进而提高抽穗期干物质积累量和群体质量,对提高作物群体生产力具有重要意义[43, 47−48]。
1.3 氮素营养对水稻产量及产量构成的影响
氮是影响水稻产量形成最活跃的元素,氮素供应水平与水稻的产量及其构成有密切关系。很多与氮代谢相关的基因可影响水稻产量构成及产量[49]。水稻施氮量与产量呈开口向下的抛物线关系,存在最适施氮量,即施氮量在一定范围内与产量呈正相关,但当氮肥用量超过一定范围后产量及部分产量构成因子下降[50]。最适的氮肥用量因水稻类型、土壤、气候条件、栽培措施等不同而存在差异[51−52]。
施氮水平显著影响水稻产量构成因素,其中穗数和穗粒数与氮水平存在显著正相关关系,但结实率和千粒重随施氮量的增加呈现降低的趋势[ 6 , 5 3 − 5 4 ]。孙伟晶等[55]发现,氮肥可促进水稻分蘖生长及颖花的分化,提高穗数及穗粒数。在高氮条件下,穗粒数显著增加,导致同化物供应水平下降,结实率及粒重显著降低,而在低氮条件下,水稻充实度高,千粒重增加。穗数及穗粒数对氮素的响应与产量相似,也存在最适施氮量。在一定施氮范围内,随施氮量的提高,穗数和穗粒数增加。但过多的氮肥会延长水稻营养生长时期,导致穗数和穗粒数下降,产量降低[56]。另外,水稻产量与施氮时期密切相关,适时追施氮肥,增加水稻中后期如幼穗分化至抽穗期的氮肥用量,可显著提高水稻产量[23, 48]。
2 栽培密度对水稻农艺性状的影响
2.1 栽培密度对水稻分蘖特性的影响
栽培密度对水稻生长发育及群体构成具有重要的调控作用。栽培密度对水稻分蘖的影响较为显著。近年来应用较广的稀植栽培技术,如“稀少平”、“旱育稀植省苗”等,可通过提高水稻单株分蘖数目,提高水稻产量[57],但密度过低,群体分蘖能力有限,增产效应无法实现。而栽培密度过高时,由于个体间的生存矛盾加剧,穗粒结构失调,水稻植株成穗率与结实率显著下降,产量降低[58]。总之,在一定范围内,栽培密度的增加导致水稻单株分蘖数和穗数减少,但单位面积的穗数随密度增加而提高,显著提高了群体生产力。
2.2 栽培密度对水稻光合特性的影响
合理的栽培密度影响水稻株型特征和冠层结构的形成,可协调个体与群体,源与库之间的矛盾,既可以发挥较强的个体优势,又有利于增加群体的光合作用从而提高产量[59]。栽培密度除影响分蘖外,也可直接影响水稻群体对光能的利用。多种水稻品种有效叶面积率、穗长等均随着栽插密度的减小而增大[60]。当栽插密度过低时,单株水稻叶片数目、叶片宽度和披垂角度增加,剑叶、倒二叶和倒三叶的净光合能力降低,导致单位面积有效穗数和叶面积指数均降低,中等密度处理下,水稻齐穗期上层叶面积高于低密度和高密度处理[61]。但密度过高时,植株间竞争加剧,叶片相互遮挡,导致单株生产力低且易诱发病虫害等,严重损害水稻质量。
2.3 栽培密度对水稻产量及产量构成的影响
栽培密度对水稻产量及产量构成因素影响显著。水稻产量各构成因素之间相互协调、相互制约,适宜的栽培密度能够保证水稻充分利用资源,保证产量构成各因素协调以达到增产的目的。研究表明,随着种植密度增加,水稻产量呈先升后降的趋势,并且最适栽培密度受到水稻品种的影响[62]。种植密度对水稻群体穗数的影响较大,但对结实率和千粒重影响较小。在一定范围内,随种植密度的增加,单位面积有效穗数逐渐增加,而每穗粒数呈相反的变化趋势,种植密度与产量呈抛物线关系[58, 63]。
闫川等[64]研究结果表明,中等行距处理下(30.0 cm× 13.3 cm),水稻在齐穗期和灌浆后期具有适宜的叶面积指数和较高的剑叶光合速率,灌浆期干物质转运量最高。且中等行距下群体微气象环境良好,水稻产量最高。张数标等[65]认为,较短生育期的水稻品种适合较高密度种植,生育期较短的品种,适当增加种植密度,其结实率和千粒重高于较长生育期品种,实现增产。而生育期较长的品种更适合较低密度种植。籼粳杂交稻和杂交籼稻的产量在种植密度为13.9 cm × 30.0 cm 时最高[66]。郎有忠等[61]研究发现,结合当前水稻栽培条件,中等种植密度处理30万丛/hm2 下水稻可获得较高产量。但也有研究认为,密度对产量相关指标影响不大,由于穗数与每穗粒数之间具有良好的互补性,故在一定范围内密度对产量影响不大[67]。
3 氮素营养与栽培密度互作对水稻产量及氮素利用效率的影响
3.1 氮素营养与栽培密度互作对水稻产量构成因子及产量的影响
氮密互作对产量构成因素中影响最为显著的是有效分蘖数。增施氮肥可促进单株水稻分蘖,显著提高有效穗数,密度则通过增加基本苗数来提高单位面积的有效穗数。低氮低密种植时,个体数量偏少且分蘖不足,即使穗粒数和结实率提高,但难以弥补穗数偏少导致的产量降低。高氮高密条件下,分蘖数增加,但个体之间对养分、光照、空间等资源的竞争加剧,吸收营养的面积和光合效率降低,个体生长受阻,后期穗的发育受到影响,导致有效穗数虽高,但穗粒数、结实率等显著降低,不利于产量的提高[68]。当低密度种植时,由于密度较低,基本苗不足,增施氮肥提高群体有效穗数的空间有限,产量增加受到限制。只有施氮量和栽培密度处于最适条件下,才可能在保证有效穗数的同时,协调群体生长环境,优化群体结构,使群体有良好的通风、光照及光能截获能力,进而稳定或提高穗粒数和结实率,提高群体生产力[68−69]。
近年来,关于氮素与密度互作对水稻产量构成因子及产量的影响研究取得了一定的进展。有研究者通过建立回归模型寻求最佳的氮密配比。郑克武等[66]发现,‘两优培九’在施氮225.0 kg/hm2、移栽密度为22.5 万丛/hm2 条件下,可达到穗数、千粒重和结实率的相互协调,进而获得高产。吴朝晖等[70]探索了超级稻最佳的氮密比,发现施氮量为270.0 kg/hm2、栽培密度为20.0 cm × 26.6 cm 时水稻群体结构最优,增产效果最好。施氮量、种植密度与水稻的结实率之间存在互作关系[71],低栽培密度条件下,适量增施氮肥可提高结实率。邓中华等[72]相继发现,‘丰两优9 号’随施氮量和移栽密度的增加,水稻有效穗数增加,但结实率和千粒重下降,并指出在22.1 万丛/hm2 的种植密度下施氮194.9 kg/hm2 是实现节氮高产的最佳组合。张悦等[73]利用两种杂交稻品种‘深优9516’和‘荣优225’为试材,研究发现,氮密互作对水稻株高、叶面积指数、干物质积累量、有效穗数、产量具有正效应,综合水稻的生长和产量效应,‘深优9516’最适宜的氮密组合为氮水平180 kg/hm2、密度27 万穴/hm2,‘荣优225’在氮水平180 kg/hm2、密度30 万穴/hm2 下生长最佳。另外,有研究指出,在高密减氮条件下近交系水稻品种优于杂交水稻品种[74]。
总之,氮素与密度互作对水稻产量的影响主要通过调节水稻群体构成而实现,取决于穗数、千粒重及结实率等各产量构成因子之间的相互协调,且受到水稻品种的影响。另外,氮素与密度互作对稻米品质产生影响,减氮密植栽培技术有利于实现稻米高产优质和减氮增效的协同[75−76]。
3.2 减氮密植对水稻产量和氮素利用效率的效应研究
众多田间试验结果表明,水稻减氮密植对产量和氮素利用效率的协同效果良好。王振洋等[77]研究结果表明,适当增加密度可提高水稻氮肥偏生产力、氮素吸收效率,并指出在施氮量为240 kg/hm2 条件下,20.83 万~27.78 万穴/hm2 的栽插密度可达到氮素利用效率及产量协同提高的目的。陈健晓等[78]探明了‘湘两优900’在热带稻区的最适氮素和密度组合,施氮量300 kg/hm2 与移栽密度25.0 万穴/hm2 对产量和氮肥利用效率综合效益最佳。樊红柱等[79]利用‘川香9838’为试材,比较了水稻不同移栽密度下水稻产量对氮肥响应程度的差异,结果表明,栽培密度和氮肥施用量分别为40 cm×40 cm 和150 kg/hm2对该地区水稻产量、氮素利用及环境污染等因素的综合效应最佳。在低氮高密和高氮低密条件下,单位面积分蘖均较多,叶面积指数和冠层光合作用潜力较大,因此,当种植密度从常规密度25.5 万穴/hm2提高到32.5 万穴/hm2,密植的施氮量180 kg/hm2 与常规密度下270 kg/hm2 施氮量相比,产量基本持平,减氮密植可显著提高氮素利用效率[22]。刘红江等[80]也得出相似的结论,在氮水平300 kg/hm2 条件下,将种植密度提高至150 万株/hm2,氮肥投入降低20%仍可保证水稻产量,显著提高氮素利用效率。梁青铎[81]发现,相较于常规种植模式,水稻减氮密植不仅可以保证产量、提高氮肥利用率,还可降低水稻倒伏指数。另外,有研究结果表明,低氮高密处理下穗数增加,且低氮条件下,密度增加可改善根系形态和分布,影响干物重、增加水稻的产量和氮利用效率[82]。
相较于水稻,玉米作为典型的高密度增产作物,前人对其减氮密植机理的挖掘较为深入。耐密型玉米植株构型包括株高降低,耳位减少,叶片直立,流穗分枝数和开花间隔减少等。有研究指出,玉米中光敏色素介导的光信号通路与miR156/鳞状启动子结合蛋白-LIKE (SQUAMOSA-PROMOTER BINDINGPROTEIN-LIKE,SPL) 调控模块可协同调控高密度下遮荫回避响应和植株结构建成各个方面。ZmSPL基因在调控玉米开花时间、株高、叶夹角、穗构型等方面发挥着重要作用[83]。在高密度种植下,无论是水稻还是玉米直立叶片可以更好地利用冠层的光能,提高光合效率、通风和抗逆性,最终提高籽粒产量[84],这也是针对高密度种植的谷类作物的遗传改良目标之一。
综上所述,通过减氮密植可优化水稻群体构成,实现作物最大群体生产力和氮肥高效吸收利用。但不同地区,不同水稻品种对氮素和密度互作的响应略有不同,以往对氮密互作的研究重点在于提高产量,肥料减量并不突出,氮肥减量一般在20%~30%左右,且减氮处理同样重视分蘖肥的施用。因此,在目前的水稻氮密互作研究中,还是重视以氮促分蘖获得一定的穗数,结合密度的增加,提高产量,氮素高效利用的潜力非常有限。赵杰等[7]提出水稻“独秆”栽培模式,即基肥不施氮肥配合全程氮肥在分蘖中后期适宜叶龄时施用,该模式可实现水稻提质增产。该实践技术显示,常规的水稻氮素营养与栽培密度种植模式很难满足国家化肥减量增效的迫切需求,势必要在栽培密度和氮素营养供应上进行深度革新。
3.3 “极限密植”对水稻产量和氮素利用效率的影响
为探究水稻减氮密植栽培的节氮增产潜力,陈世勇等[8]自2015 年开始,通过在安徽省及江苏省不同稻作区进行减氮控蘖密植栽培田间小区试验和示范推广,总结出一套水稻“极限密植”栽培技术,即通过优化施肥、氮肥运筹、免施分蘖肥、控制分蘖数目、协调群体生长与主茎成穗提高水稻产量,可显著减少氮肥用量,实现水稻高产和氮素资源高效利用协同。
“极限密植”可采用该水稻品种传统高产模式下的单位面积有效穗数作为“极限密植”模式的栽插密度,以优质中熟晚粳‘南粳46’为例,控制密度约400×104 株/hm2[85],既可以保证高产,同时为减肥创造条件。以氮素供应为例,水稻体内氮素含量必须达到3.5% 以上才会分蘖,否则分蘖受到抑制[19]。而这势必需要显著提高土壤中氮的供应强度才可能实现,导致土壤氮素的大量损失,降低氮肥利用率。因此,水稻体内氮素含量必须达到3.5% 以上才能分蘖被我们视为传统水稻高产模式的系统性风险,也是传统水稻高产模式虽然高产,但养分资源难以高效利用的主要原因。另外,由于土壤对养分的吸附固定,且养分迁移受到迁移距离、养分浓度等因素的影响,稻田土壤中各种速效养分的迁移能力有限,能有效到达根际的比例较低。显然,常规栽培密度不利于水稻根系,尤其是苗期小而弱的根系吸收利用土壤中的养分少(图2),从而造成肥料的经济效益降低。而“极限密植”技术大幅度提高了本田中苗期水稻根系与土体的接触面积,相应地增加了养分吸收。
以江苏省为例,水稻种植密度在24~33 穴/m2,每穴2~3 株苗[22, 86],“极限密植”基本苗数可达到常规栽培密度的4~8 倍。“极限密植”不追求分蘖形成产量,但也不是说完全控制分蘖。由表1 可知,“极限密植”条件下分蘖也不会被完全抑制,但是与常规栽培条件下水稻每株3.5 个有效分蘖相比,极限密植每株有效分蘖数显著降低,为1.2 个分蘖,分蘖生长受到显著抑制。因此,“极限密植”主要是通过增加基本苗栽植密度达到高产所需的有效穗数,分蘖期不提供分蘖发生的营养条件,使得稻株不发生分蘖或分蘖数极少。同时,由于免施分蘖肥,不会发生群体过大,相互荫蔽、影响生长而造成倒伏或减产。极限密植水稻田间生长状态良好,且与常规栽培相比,抗倒伏情况有显著改善(图3)。我们前期关于“极限密植”的研究依据主要考虑到目标产量、土壤养分供应量、肥料利用率、干湿沉降及灌溉水对土壤养分的影响等情况,9000 kg/hm2 目标产量的氮素施用量定为90 kg/hm2,适用于中等及以上肥力的土壤。陈世勇等[8]试验结果表明,极限密植优化施肥处理下,产量和氮肥利用效率均显著高于常规密度栽培模式下的施肥处理。进一步对其产量构成因子分析可得,相较于常规栽培,极限密植每穗粒数降低了9.7%,但穗数、千粒重和结实率分别增加了2.2%、12.75%、4.1%,最终产量增加了27.4%,氮肥农学利用效率增加了120% (表1)。因此,通过优化施肥下极限密植增加水稻产量和氮素利用效率的潜力是巨大的。
4 研究展望
作物“减氮增效”是现阶段我国农业绿色发展面临的重要挑战,氮素营养与移栽密度在调控水稻产量和氮素利用效率方面起到重要作用。受“分蘖促控”的影响,现阶段水稻高氮低密的种植模式还很普遍,氮素高效利用的潜力十分受限。近年来水稻减氮密植种植模式可通过氮密互作协调群体构成,提高水稻产量与氮素利用效率,为水稻“减氮增效”提供了新的技术路径。但氮密互作受到水稻品种、田间管理、土壤条件等限制,最适的氮密互作技术参数尚需进一步探索。
未来氮素营养与移栽密度调控水稻产量和氮素利用效率亟待从以下几方面深入探究:作物高产栽培要求良种与良法相结合,不同水稻品种,因其株型、养分利用效率等存在差异,对氮密互作的响应不同,粳稻和籼稻的分蘖特性及氮素利用效率存在显著差异[11],因此,应针对不同的水稻品种开展氮密互作的研究,探索更大的水稻增产和氮肥减量潜力;水稻氮密互作的研究离不开适宜的栽培措施,尤其是减氮密植种植模式下,正在兴起的水稻无人机飞播技术恰好具有高密度精准播种的特性,可望在氮密互作的研究中发挥重要作用;目前,氮密互作结合土壤类型和土壤肥力状况的研究较少,应开展针对不同地区、不同土壤状况和肥力状况进行最适氮密互作的探索,也可结合有机肥配施开展氮密互作技术的相关研究。
另外,目前关于氮密互作的研究大多停留在产量和氮素利用效率的生理表型上,具体的生理和分子调控机制还需深入研究。下一步可探究在一定种植密度条件下氮肥减量对水稻植株构型的影响,包括叶面积指数、叶夹角、不同冠层叶片光合性能、根系分布状况等;结合15N 示踪技术探究水稻在不同的生育时期,植株各个部位氮素吸收和转化特性,进一步找到在最适氮密互作条件下,调控水稻株型和氮素利用效率的关键基因。
此外,氮和碳均是植物生长发育不可或缺的重要元素,碳氮协同对植物生长和生态环境有重大意义。因此,需要进一步探究氮密互作对水稻氮代谢和碳固定的作用机制,进而在生产上实现水稻高产和资源高效利用的协同,为绿色低碳农业发展和我国粮食安全提供新的理论支撑和实践路径。
