水稻根系分泌物研究进展
2024-12-31张怡李建国张文君
关键词: 水稻; 根系分泌物; 影响因素; 种类; 功能
根系分泌物作为根土对话的媒介,其作用早在18 世纪已被发现,近年来,随着新技术和方法不断出现,根系分泌物研究逐步推进。被广泛接受的根系分泌物功能主要包括影响土壤养分有效性及养分循环[1−3],招募微生物定殖抑制根际病原微生物[4−5],调控根际微环境促进植物应对各种生物和非生物胁迫等[6]。另外,植物根际还可以通过化感或自毒作用直接影响植物间的共生关系[7]。深入了解根系分泌物种类及其功能,解析分泌物介导的植物−土壤−微生物互作过程,是当今土壤学、植物营养学、微生物学及环境学关注的热点。
水稻(Oryza sativa L.) 是全球最重要的粮食作物之一,我国约有60% 的人口以稻米为主食[8]。水稻的生长和产量受到多种环境因素的调控,根系分泌物承载着植物−土壤−微生物互作中的信息交流、物质循环和能量流动,因此,分泌物的组成对植物养分吸收、土壤养分有效性及相关微生物组成及多样性等都具有重要的影响[9−10]。本文综述了水稻根系分泌物的种类、影响因素、功能以及环境影响,并展望了根系分泌物需要重点关注的研究领域,以期为绿色农业可持续发展提供理论基础。
1 水稻根系分泌物研究发展历程及种类
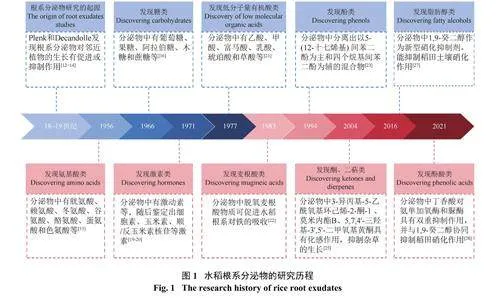
植物根系分泌物的研究发展从成分鉴定到相关功能分析经历了300 多年的时间(图1)。早在18 世纪Plenk (1795) 以及19 世纪Decandolle (1830) 都观察到根系分泌物对邻近植株的生长具有促进或抑制作用[11−13],随后德国科学家Hiltner 提出了“根际”的概念,并开展了大量关于根系分泌物的研究[14]。Andal等[15]收集了水稻根系分泌物,发现分泌物中含有氨基酸类化合物如胱氨酸、赖氨酸、冬氨酸等,随着研究的深入,天冬氨酸、异亮氨酸等氨基酸也逐步被鉴定出来。而后MacRae 等[16]进一步揭示根系分泌物中糖类主要是葡萄糖、果糖和蔗糖等,这些糖类分泌物不仅有利于根系下扎,而且对土壤颗粒具有很强的粘着力,可以促进微团聚体的形成,改善土壤的物理性状,也可改变土壤微生物群落结构,为许多有益根际微生物提供碳源,同时对稻田甲烷(CH4)的产生也有影响[17−18]。研究还发现水稻根系能分泌出激素类物质如激动素等[19],之后鉴定出细胞素、玉米素、顺/反玉米素核苷等激素[20]。根系分泌物中低分子量有机酸为乙酸、草酸和琥珀酸等,为土壤中磷、钾、硅养分的活化提供了有效途径[21],并且水稻根系分泌的脱氧麦根酸,其作为铁载体,可有效缓解缺铁胁迫,从而促进水稻对铁的吸收[22]。水稻根系分泌物中酚类物质也逐渐被发现[23],其中对香豆酸能抑制土传病原菌尖孢镰刀菌引起的枯萎病[24]。随着植物化感作用的发现,水稻根系分泌物发挥化感作用的物质有了明确的分类,如水稻根系分泌物中的黄酮、酮类和萜类均具有化感作用,其中莫米内酯B 会抑制周围杂草的生长[25]。另外,水稻根系分泌物中的激素类物质独脚金内酯,在响应植物养分缺乏时也能发挥其重要的作用,其可作为根际信号,增强丛枝菌根真菌的共生能力以提高水稻对无机磷的获取[26]。近年施卫明团队从水稻的根系分泌物中发现了1, 9-癸二醇,它能够抑制稻田中的硝化作用,减少氮的损失[27]。最新的研究还表明,水稻根系分泌物中的丁香酸对参与硝化作用的氨单加氧酶(AMO) 和脲酶具有双重抑制作用,并可与1, 9-癸二醇协同抑制稻田中的硝化作用[28]。
水稻根系分泌物从发现到鉴定,经历了漫长的探索历程。目前已知水稻分泌物除了常见的氨基酸、糖类、有机酸、酮类、脂肪酸、酚类等成分外,特有分泌物如脱氧麦根酸、1, 9-癸二醇等也被逐渐发现。水稻根系分泌物作为一个“宝藏”,在土壤养分活化、化感作用、抗病促生、抵抗生物和非生物胁迫等方面均发挥着重要的作用,这些分泌物的种类和组成对于水稻的生长和环境适应性具有调控作用。
2 水稻根系分泌物释放的影响因素
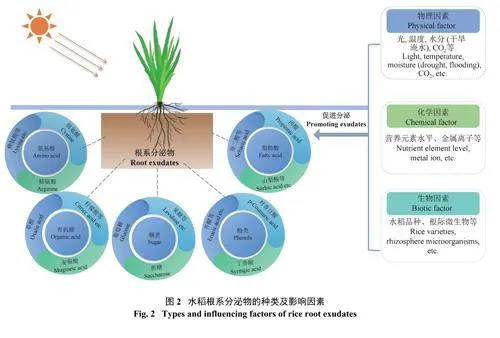
水稻根系分泌物的成分(氨基酸、有机酸、脂肪酸、酚类和糖类等) 和数量受到外界环境中物理、化学和生物因素的影响(图2),这些因素相互作用,共同调节着水稻根系分泌物的特性,进而调节着水稻在不同环境条件下的适应性和生长过程。
2.1 物理因素
光照、温度、水分等环境条件均能影响根系分泌物的产生。研究表明,光合作用中重要的同化物二氧化碳(CO2) 含量高时,会增加根系分泌物的释放[29−31],高浓度CO2 处理下,水稻根系分泌物中甲酸和乙酸的释放总量会增加[32],还有研究发现,随着CO2 浓度的升高,成熟期水稻根际原儿茶酸、芥酸、绿原酸等酚类化合物的浓度显著增加[33]。在适宜温度范围内,大多数植物的根系分泌物释放量会随温度升高而增强,并且环境温度升高还会改变其组成,如增加氨基酸并降低酚类化合物的相对丰度,原因是温度升高后土壤微生物群落从细菌优势向真菌优势转变[31]。水分状况也是影响水稻根系分泌的重要因素,干旱条件下,根系粘液释放量的增加可在根表面形成根鞘,以减少水分流失[34],且增加养分的吸收。另外,有机酸分泌上调和抑制氨基酸分泌是水稻应对干旱的重要策略,因为根际微生物组与根系分泌物代谢组高度相关,其中对干旱有响应的链霉菌、芽孢杆菌和放线菌丰度差异与根分泌物中有机酸和氨基酸含量显著相关。且进一步的研究表明,链霉菌在干旱条件下受脱落酸和茉莉酸的调控[35]。另外,干旱还可能增加土壤颗粒对根系的机械压力,从而刺激根系分泌物的产生[36],而根系分泌物作为土壤结构形成的联结者,当土壤干燥时,根系分泌物和微生物释放的多糖具有缓冲作用,又能使局部保持湿润,降低对土壤团聚体的破坏性。这些复杂的相互作用机制使得水稻根系分泌物在不同环境条件下表现出多样性,为植物适应不同生长环境提供了一系列策略。
2.2 化学因素
与正常条件下相比,在低营养胁迫条件下,植物根系分泌物的种类与数量会发生变化,这有助于植物适应环境变化,获取营养并缓解胁迫[37]。如低磷条件下,粳稻的脯氨酸、丝氨酸、苏氨酸、缬氨酸和4-香豆酸的分泌量均增加[38]。钾、钙营养水平对根系分泌物有调控作用,进而影响稻米品质,结实期钾、钙营养水平对水稻根系分泌物的影响可能与水稻自身对环境适应性有关,而相关机理还有待深入研究[39]。此外,水稻在缺铁条件下根系会分泌一种特殊的有机酸,即2´-脱氧麦根酸,这种酸能够活化并螯合土壤中的三价铁,从而缓解缺铁胁迫[40]。
2.3 生物因素
2.3.1 植物品种差异及生长发育阶段 植物根系分泌物的组成和含量会受到物种、生长阶段等因素的影响而存在差异。不同物种植物的根系分泌能力差异显著,例如,在缺铁条件下,水稻、小麦根系分泌脱氧麦根酸,而大麦则分泌羟基麦根酸[40]。水稻在不同生育期的有机酸分泌量也存在显著差异,研究结果表明,水稻一般在苗期根系分泌物质量最低,在开花前增加,在成熟时降低,并且随着生育期的推进,有机酸的分泌量逐渐超过了糖的分泌量[41]。此外,通过对不同水稻品种根系分泌物中有机酸的量化发现苹果酸的浓度最高,其次是酒石酸、柠檬酸和乙酸,这种基于有机酸类型和分泌量的差异,表明根系分泌物的生物合成和释放受到基因型和营养特异性的调控[42]。
2.3.2 根际微生物 水稻根系分泌物会影响根际微生物的生长和活动,同时植物根际的微生物也可以通过根系信号机制影响根分泌物的种类和数量,原因是根际微生物的代谢物质可以影响植物信号的产生,从而改变根系分泌物的成分并诱导释放更多的分泌物,促进水稻根系富集更多的促生菌[43−45]。根际微生物活动对植物根系分泌物的影响比较复杂,例如,一些根内生菌可能通过分泌细胞壁纤维素分解酶(如纤维素酶、木聚糖酶、果胶酶和内聚糖酶) 来改变植物细胞壁结构,同时固氮内生菌释放的内切葡聚糖酶会影响内生菌在水稻中的定殖[46],进而影响根系分泌物的释放;当根系受到有害微生物如细菌、病毒感染时,植物为抵御环境胁迫会改变糖、氨基酸等初生代谢物种类和产量,并刺激次生代谢物发生变化。另外,在病原微生物的持续攻击下,拟南芥、水稻、玉米等植物根系会分泌抗菌吲哚、萜类、苯并恶嗪酮和类黄酮或异黄酮等天然产物,以抵御病菌的入侵和感染,发挥抗菌的作用[47];此外,微生物能够改变根际的矿质营养状况和植物激素含量,调节植物的生理生化过程,从而影响根系分泌物的组成和数量[29];例如,内生菌株通过合成吲哚-3-乙酸(IAA)、细胞分裂素和赤霉素等植物激素来促进植物生长[48]。植物内生菌产生的IAA 增加了侧根和不定根的数量,增强根系分泌物数量,促进养分的获取,为土壤微生物与根系相互作用提供了更多能源物质[49]。同时根际微生物通过消耗和转化根系分泌物中的有机物,造成各种有机物的浓度梯度,而根际微生物释放出一系列的代谢产物作为化学信号,激发根系分泌更多的分泌物[50]。
因此,影响水稻植物根系分泌物质的因素复杂多变,通过深入了解影响根系释放分泌物的因素,可以更好地掌握其对根系生理活动的调控作用,这将有助于深入理解根系释放分泌物的功能。
3 水稻根系分泌物的功能
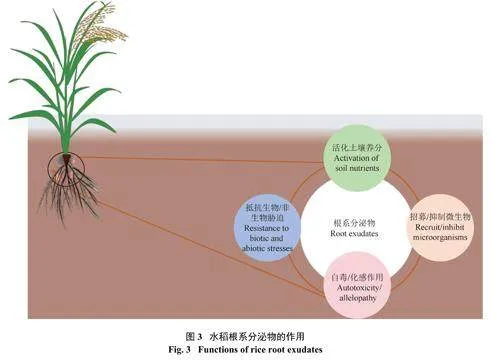
水稻根系分泌物在其整个生命周期中发挥着关键作用,主要体现在以下几个方面(图3)。首先,水稻根系分泌物中的有机酸对土壤养分(氮、磷、钾、铁和硅等) 的活化具有重要作用 (图4)。其次,分泌物与微生物之间存在互作效应,能够促进菌丝生长,形成植物−微生物的共生网络,这有利于养分循环,提高土壤的肥力。此外,水稻根系分泌物中含有的化感物质能有效抑制杂草的生长,从而减少杂草对养分的竞争,维持良好的生长环境。总之,水稻能通过调节根系分泌物的种类和数量,从而增强抵抗生物/非生物胁迫的能力。另外,水稻根系分泌物不仅对其本身有益,还可能在一定程度上影响其所处的土壤环境及周围植物,发挥着重要的生态作用。这种互惠作用机制使得水稻根系分泌物成为维持水稻健康生长和土壤生态平衡的重要因素。
3.1 根系分泌物对土壤养分的活化作用
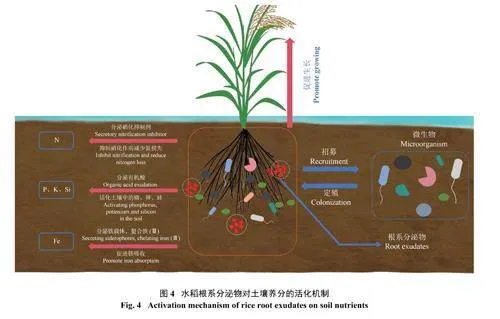
3.1.1 氮 研究发现,某些植物释放的特定根系分泌物能抑制或减缓硝化作用[51],这种根分泌物被称为生物硝化抑制剂(BNIs)。这些物质能够选择性地抑制土壤中硝化细菌的活性,从而减缓土壤中铵态氮向硝态氮转化的速度。目前已经从多种植物中鉴定出多种BNIs,如臂形草内酯[52]、高粱酮、樱花素[53]和对羟基苯基丙酸甲酯等[54]。在水稻中鉴定的两种硝化抑制剂1, 9-癸二醇以及丁香酸[27−28]能有效抑制稻田的硝化作用,尤其是1, 9-癸二醇对我国的3 种典型农田土壤(碱性潮土、中性水稻土和酸性红壤) 的硝化活性均有显著抑制作用[55]。良好的通气状况有利于水稻分泌更多1, 9-癸二醇,因此,根系分泌BNIs 和根系泌氧可共同调控根际硝化强度[56],从而有利于水稻对氮素的吸收[57]。研究还发现,水稻根际中亚硝化细菌能够特异刺激1, 9-癸二醇的分泌,而反硝化细菌则没有这种诱导效应[56]。最新的研究表明,水稻根系分泌物中鉴定出的酚类化合物丁香酸作为硝化和尿素水解的潜在抑制剂,不仅对AMO 表现出双重抑制作用,而且还显著抑制了氨氧化细菌(AOB) 和氨氧化古菌(AOA) 的丰度,同时在与1, 9-癸二醇协同抑制硝化作用过程中表现出更强大的效果[28]。因此,当水稻根系感应到有利于硝化作用的因子后,能主动产生特定信号物质以分泌BNIs,调控根际硝化强度至适宜范围内。因此,这些研究有望为稻田的施肥管理提供新思路,尤其是BNIs 物质作为新型的氮肥增效剂,能提高水稻的氮肥利用率而达到增效减排的效果,同时也为土壤氮素循环中植物−微生物相互作用过程提供了新见解[27]。
3.1.2 磷 磷是植物生长发育所必需的大量矿质养分元素之一,参与蛋白质合成,调节能量代谢、光合作用和呼吸作用等[36,58]。尽管土壤中的磷很丰富,但由于磷很容易被土壤矿物吸附和固定,形成Fe-P、Al-P、Ca-P 等难溶性磷酸盐,植物能吸收利用的磷却很有限。在缺磷土壤中,植物根系会通过多种方式增强对磷的吸收能力,包括改变根系构型,与菌根真菌建立共生系统,增加根系分泌物以及增强磷转运蛋白的表达等多种方式来增强其获取磷的能力[59−60]。其中,根系分泌物在植物转化利用土壤中潜在磷库的过程中起着关键作用[61]。根系分泌的多种有机酸,如苹果酸、柠檬酸和草酸,已被证明可以通过螯合和酸化作用活化土壤中难溶性磷,以供植物根系吸收[62]。在土壤有效磷含量较低的情况下,根系分泌物通过刺激磷酸酶的合成及释放,促进土壤有机磷的水解[ 6 3 ]。水稻根系分泌有机酸,如柠檬酸、草酸等,对土壤中磷的转化机制与大多数植物相似,主要共性机理有以下几个方面:1) 吸附位点的竞争 有机酸以完全解离的阴离子形式存在于土壤中[64],占据土壤矿物质的吸附位点,占据磷在Fe/Al氧化物表面的吸附位点,或与磷竞争土壤颗粒的结合位点,减少磷在土壤中的吸附和固定[65],2) 表面电荷的变化 土壤中的Fe/Al 氧化物通常带正电荷,在吸附了有机阴离子后,降低了矿物与磷之间的静电作用,从而减少了对磷酸盐的吸附和固定,提高了土壤磷养分的生物有效性[29];3) 矿物溶解 酸的增溶和络合作用共同促进了矿物质磷的溶解[ 6 6 ];4) 招募微生物 招募的解磷微生物通过释放有机酸和磷酸酶,分别促进土壤中无机磷的释放和有机磷的分解,提高土壤中磷的有效性[67]。有些植物根际促生菌,如假单胞菌,它们不仅可以释放有机阴离子和磷酸酶来活化及利用土壤磷,而且还能分泌激素来促进根系发育,从而增加根系对磷的吸收[68]。此外,外生菌根菌丝可以向土壤中释放草酸和酶等,以调动土壤无机磷和有机磷的循环转化[69]。
3.1.3 钾 钾作为肥料三要素之一,它对植物的生长发育具有重要影响[70]。水稻全钾含量中约有39.0%~72.7% 来自于非交换性钾[71]。当植物面对缺钾胁迫时,分泌有机酸能活化土壤中的矿物态钾,从而促进土壤钾的有效化,增强植物对缺钾环境的适应能力[65]。而有机酸对土壤中钾养分释放的作用机制主要是通过酸化和络合两种方式共同发挥作用。有机酸的浓度影响着其对钾的活化类型,高浓度时以酸化作用为主,低浓度时则以络合作用为主[72]。研究表明,低分子量有机酸,如酒石酸和草酸,能显著提高不同含钾矿物(黑云母、白云母和微斜长石) 的钾释放量,黑云母、白云母和微斜长石的累积释钾量均表现为草酸gt;酒石酸gt;水,不同矿物中均以草酸作用的钾释放量最大,分别为4734.7、4661.2 和812.0 mg/kg。其中,草酸主要是通过酸性水解和络合溶解双重作用来有效活化土壤矿物钾[73]。因此,水稻根系分泌的有机酸通过促进土壤中含钾矿物的溶解效应,为水稻提供可吸收利用的钾养分。
3.1.4 铁 铁在土壤中的含量也较高,然而在大多数情况下,特别是通气良好条件下,铁主要以三价铁(Fe3+) 存在,如水铁矿、针铁矿等,很难被植物吸收利用[74]。水稻作为一种特殊的禾本科植物,在长期淹水的生长环境中,形成了两种铁的吸收机制,既能利用还原作用吸收土壤 三价铁 (机制 I),又能通过螯合作用,吸收吸附在根表的三价铁 (机制 II)[75]。机制I 主要是基于还原机制提高对土壤中Fe3+的吸收与利用。根表皮上的三价铁螯合还原酶将Fe3+还原成Fe2 +,由根系分泌的次生代谢物质尼克酰胺螯合游离的Fe2+,再由铁转运蛋白作为转运体运输进入胞内[76−78]。机制II 主要是水稻通过根系分泌铁载体麦根酸类物质来增强根际铁的移动性。不同的禾本科作物分泌的麦根酸种类不同,如水稻、小麦仅能分泌脱氧麦根酸(DMA),而大麦则能合成DMA、麦根酸(MA) 和3-表-羟基麦根酸(epi-HMA) 3 种[79]。铁载体对Fe3 +具有很强的络合能力,可以特异性螯合Fe3+,从而提高铁的迁移率和利用效率[80]。
3.1.5 硅 水稻是典型的硅(Si) 积累作物,平均硅含量可达茎干重的10%[81]。水稻吸收的硅主要来自土壤,在不同形式的硅中,植物只能利用可溶性硅酸,但土壤溶液中的可溶性硅含量非常有限,通常为3~40 mg/L。水稻的连续种植带走了土壤中大量的硅,这很容易导致水稻土中可溶性硅的缺乏,这种现象在高风化的热带地区中尤为严重。因此仅依靠土壤中硅的自然释放难以满足作物对硅的吸收需求[82]。而水稻根系分泌物对硅的释放具有重要的调控作用,如分泌物中的有机酸主要通过酸增溶、螯合、离子交换等方式,加速土壤中硅的解吸和硅铝酸盐的溶解[83]。在含有铁氧化物的土壤中,由根系大量分泌的草酸、柠檬酸、苹果酸3 种有机酸能显著增加硅和铁的共释放[83]。同样,绿泥 石等 2∶1 型粘土矿物、高岭石等 1∶1 型粘土 矿物,以及长石等原生矿物均可通过有机酸配体 的增溶效应释放出硅酸[84]。此外,除了有机酸的作用外,根系分泌质子可引起根际局部酸化[85−86],从而减少单硅酸的聚合或吸附,增加被矿物固定的硅酸盐的解吸,提高土壤溶液中可溶性硅的浓度。总之,水稻根系分泌的有机酸和质子都可增加硅的释放,从而提高植株对土壤硅的利用效率。
3.2 根系分泌物对植物−微生物/植物之间相互作用的影响根系分泌物除了直接参与土壤中的养分活化外,其分泌的酚类等次生代谢产物等信号物质具有间接调控的作用,成为植物与植物、植物与微生物之间传递信号的桥梁 (图4)。
3.2.1 植物−微生物 根系分泌物对根际微生物的影响具有多样性,在植物的生长过程中,这些分泌物能刺激细胞外酶,如蛋白酶、水解酶等的产生,可以降解复杂的有机物质并用于植物和微生物的营养吸收,为微生物提供碳源,根系分泌的有机酸能通过促进细菌的生长、趋化性和定殖招募有益微生物,调节根际微生物群落结构和种群分布[87−88]以及根系与土壤微生物之间的互作效应[89],含有特定次生代谢物的分泌物还通过招募或排斥不同的微生物群落,塑造独特的根际微生态环境[90]。例如,水稻根系分泌物作为电子供体和碳源可促进微生物生长,如厌氧杆菌利用乙酸作为电子供体,将Fe3 +转化为Fe2+,进一步促进了微生物对Fe3+的还原过程[91]。同时,水稻根系通过分泌柠檬酸驱动水稻土壤中微生物硫和铁的循环,促进铬(VI) 的减少和固定化[92]。此外,化感水稻PI312777 的根系分泌物能够明显促进水稻根际细菌、固氮菌、氨化细菌和硫化细菌等微生物的生长,而对真菌、反硫化细菌和反硝化细菌有一定的抑制作用[93]。
根系分泌物是微生物趋化系统的触发因子,它可以通过诱导根际细菌的趋化反应来招募并帮助细菌在植物根系表面定殖[65, 94]。例如,苹果酸和柠檬酸被证明是枯草芽孢杆菌(RR4) 等根际微生物的碳源和化学引诱剂[51, 95]。水稻根系分泌物中的柠檬酸对高地芽孢杆菌具有趋化作用,其机制是柠檬酸通过增加细菌趋化和生物膜形成等相关基因的表达,来促进高地芽孢杆菌的定殖,两者进而协同作用促进种子萌发和水稻生长[51]。此外,小信号分子(如有机酸和氨基酸) 可以招募植物根际促生菌(PGPR) 并调节根际微生物组的结构[96−97]。有研究表明,PGPR 通过诱导生物合成来促进根系分泌物的分泌[ 9 8 ],例如,PGPR (RR4) 的存在可以促进水稻根系苹果酸合成酶的表达,增加苹果酸分泌量。而且也可通过PGPR产生有益的代谢产物(如赤霉素和生长素),增加根系生物量来增强植物分泌[99],例如,从水稻根际分离的PGPR 菌株释放吲哚乙酸,分别调控植物茎和根的向光性和向地性,促进植物生长[100]。除了直接作用外,PGPR 还可以通过释放代谢产物如抗生素、挥发性物质、酶类等物质提高水稻对病原菌或逆境的抗性,间接促进植物生长。
3.2.2 植物−植物 根系分泌物在植物与植物之间的相互作用中发挥着特殊作用。作为介导植物间相互作用的信号因子,根系分泌物以不同的方式介导同种或不同物种植物间的相互作用,而这取决于根系分泌物的组成、相互作用的机制以及植物的信号受体[101]。其中,化感作用是一种植物间相互作用的形式,植物通过分泌化学物质来抑制同一物种(自毒性) 或不同物种的其它植物的生长或引起其死亡[102]。以水稻为例,Li 等[103−104]研究表明水稻根系分泌的苯甲酸和肉桂酸衍生物等化感物质,在抑制周围杂草的生长中发挥着重要作用。此外,水稻分泌的萜类物质如莫米内酯B,是一种内源性天然除草剂,能够抑制稻田杂草的生长[105−106]。研究还发现化感水稻苗期根系在土壤中的横向分布与其原位抑草效果有关,这些化感物质通过诱导细胞结构改变,破坏抗氧化系统稳定,以及影响呼吸/光合作用和养分吸收等方式对其它植物产生负面影响[107]。这为提高水稻苗期抑制杂草生长的能力提供了新方法,同时也为通过加强水稻品种根系性状或改变水稻根系在土壤中的分布,抑制田间杂草生长提供了科学依据[103]。
除了对其它物种产生化感作用外,根系分泌物还对相同物种或家族的植物生长产生影响,这被称为自身化感作用。例如,水稻的残茬在水中的分解物(对羟基苯甲酸、香豆酸) 会抑制下一季水稻的生长,生产上主要表现为连作障碍[108]。不仅如此,双季稻中化感物质引起的自毒作用导致产量减少约20%[109],引起这种现象的主要原因可能是根系分泌酚酸类、萜类和黄酮类物质的化感作用。
3.3 水稻根系分泌物的环境影响
稻田是CH4、CO2、N2O 等温室气体排放的主要来源之一[110],其中中国稻田CH4 排放量约占世界稻田总排放量的18%[111]。稻田CH4 的排放包含CH4 产生、氧化和运输排放3 个过程[112−113],有机底物和无氧环境是稻田土壤产CH4 菌释放CH4 的必要条件,产CH4 菌等在较低氧化还原电位下,利用根系分泌物、地上部植株脱落物等物质产生CH4[114−115]。水稻植株可通过凋落物及根系分泌有机酸、碳水化合物等物质为产CH4 菌提供底物,产CH4 菌进一步将分泌物转化为较简单的产CH4 前体乙酸、甲胺、甲醇、甲酸等物质,并代谢转化为CH4[116]。有研究证明,稻田CH4 排放量与根系分泌物的分泌量相关[117],根系分泌物中有机酸和有机碳含量越高,CH4 产生量也就越高[118]。另外,不同水稻品种和水稻不同生育期的根系分泌物成分存在差异,在水稻幼穗分化期根系分泌物较多,可增加CH4 的产生和排放[119]。在水稻生长的中后期根系分泌物和脱落物的量进一步增加,导致成熟期稻田土壤CH4 排放量远高于分蘖期和孕穗期[120]。因此,水稻根系分泌有机物质的强度越大,产生和排放的CH4 就越多。最新研究认为根系分泌物(苹果酸、琥珀酸、柠檬酸) 会增强CH4 氧化菌的丰度和活性,降低CH4 的排放[121]。同时根系分泌物是微生物可利用碳的不稳定来源,可导致CO2 排放增加,加速碳的周转[122]。最近有研究表明不同碳氮比的根系分泌物增加了水稻土的CO2排放[123]。碳充足时,根系分泌物是限制微生物竞争的重要因素[124]。此外,根系分泌物中含氮化合物可以诱导微生物分解有机碳化合物释放更多的CO2。因此,根系分泌物碳氮比和土壤速效氮可共同调节CO2 排放。同样,根系分泌物中有机酸、氨基酸和糖类作为外源物料添加到土壤中可调控N2O 的排放,如高浓度葡萄糖和低浓度草酸处理下N2O 排放量最高,且根系分泌物添加的浓度越高,为土壤所提供的碳源越多,越促进异养微生物作用下的反硝化过程,对N2O 排放的贡献越大[125],与此同时,水稻根系分泌的1, 9-癸二醇这类生物硝化抑制剂,恰好能通过抑制硝化作用而降低土壤N2O 的排放[55],从而动态调控稻田土壤N2O 的排放量。
水稻根系分泌物不仅对稻田温室气体排放有显著影响,还会影响土壤中的重金属和有机污染物的固定和活化,进而影响污染物在土壤−植物系统中的迁移转化行为。如水稻根系分泌物中的大分子粘胶类物质(如多糖)、低分子量有机酸和铁载体等天然螯合剂会影响根际的重金属形态及其有效性。其中,根系分泌的黏性物质可以与根际中的镉络合,使镉元素转化为不易被水稻根系吸收的螯合物,降低水稻植株中的镉含量[126]。然而,稻田土壤中的镉在酸性条件下可溶,当水稻根系分泌物中的H+增加,导致pH 值降低时,难溶性镉会转化为可溶性状态[127]。因此,水稻根系分泌物通过直接或间接作用(如影响根际微生物) 对根际重金属的形态及迁移转化产生重要影响。此外,有机污染物方面有研究表明,人工添加植物根系分泌物可促进有机污染物多环芳烃(如菲和芘) 在土壤中的解吸,并提高其降解率[127],根系分泌物在此过程中具有协同作用,有效促进有机物最终分解为简单无机物[128]。另外,研究认为根系分泌物也可加速污染物的释放,如根系分泌物中的低分子量有机酸,特别是柠檬酸,显著增加了多环芳烃残留物在土壤中的释放[129]。Du 等[130]通过水稻盆栽试验和添加根系分泌物后的土壤培养试验发现,根系分泌物通过增加土壤溶解有机碳(DOC),促进了土壤中增塑剂邻苯二甲酸酯(PAE) 的解吸,增加了土壤中邻苯二甲酸二丁酯(DBP) 和邻苯二甲酸二(2-乙基己基) 酯(DEHP) 的含量。总之,水稻根系分泌物作为根际环境的重要组分,在调节温室气体排放、调控土壤中的重金属和有机污染物迁移转化等方面具有重要的意义及广阔的研究前景。
4 展望
随着越来越多不同种类的分泌物被鉴定出来,根系分泌物的功能也逐渐被发掘和重新认识,特别是在活化土壤养分,塑造根际微生物群落结构,调控环境效应中的重要作用。但是关于水稻根系分泌物的研究,还需要关注以下几个方面。
4.1 根系分泌物组分的鉴定方法仍需改进
在不同生长环境条件下,水稻根系分泌物具有异质性、复杂性、不可见性。目前分泌物的收集主要通过溶液培养、基质培养等试验方法,无法从植物所处的真实环境去原位收集。此外,收集根系分泌物对根系不可避免的造成扰动和伤害,无法保证根系分泌物的成分被完全收集并被鉴定。建立根系分泌物的原位检测分析或无损收集方式,是今后精确鉴定不同环境条件下的分泌物成分的必要前提。另外,水稻根系分泌物会随着生育期变化而动态变化,因此,对水稻整个生育期分泌物的动态监测,才能更深入客观了解植物−土壤互作关系。水稻特殊的生长环境必定有其特异的根系分泌物,对其特异成分的鉴定仍将是未来研究的热点。
4.2 深入研究根系分泌物中各成分的协同机制
虽然水稻分泌物的功能大体已有结论,其活化土壤养分(如磷、钾等)、调控微生物群落、抵御生物和非生物胁迫等功能也已得到共识。但在复杂多变的自然环境中,作为混合物,水稻根系分泌物中的各种组分是如何有效的协同响应不同环境条件的胁迫,目前尚不明确。如水稻同时面对多种养分缺乏胁迫时,根系如何有效协调有机酸、糖类、酚类等物质的分泌,并在微生物的协同响应下应对复杂环境下的变化,也是需要进一步重点关注的问题。另外,随着对不同种类分泌物功能的逐渐挖掘,以及特异分泌物的鉴定,可合理有效的开发专性肥料添加剂等产品,为绿色农业的发展提供有效支撑。
4.3 根系分泌物的环境效应
根系分泌物是植物与土壤交流的化学语言,其对环境的感知可转化为信号分子,刺激根系产生化感作用,或者避开土壤中的污染物。因此,探索如何通过靶向调控根系分泌物种类和数量,优化土壤微生物群落结构,来提升根系分泌物在养分活化、污染物降解等多方面的功能。此外,水稻根系分泌物作为CH4 产生的底物,在“双碳”新形势下,研究根系分泌物对有效、绿色的减排增汇有重要意义。
总之,根系分泌物多维度的交叉研究,涉及生态平衡、土壤修复和农业生产等多个层面,深入了解植物−土壤−微生物−生态体系的运转及调节机制,可为实际生产中充分利用根系分泌物,实现经济、环境效应的最大化和最优化提供理论基础。
