菌剂对种养废弃物堆肥中腐殖质及酶活性的影响
2024-12-31陈鑫李昌宁晁艺李萍姚拓孙永平





摘要: 【目的】研究接种自制复合菌剂和商用菌剂对牛粪、菌糠与尾菜混合堆肥中腐殖化程度和酶活性的影响,为种养废弃物的资源化利用提供高效复合菌剂。【方法】以牛粪、菌糠与尾菜(湿重比5∶4∶1) 为原料进行混合堆肥,初始混合堆料碳氮比约为30∶1,堆料量为1.5 t,高度为100~120 cm,条剁式堆肥。CDS 菌剂主要由皮氏不动杆菌(Acinetobacter pittii)、枯草芽孢杆菌粪便亚种(Bacillus subtilis subsp. Stercoris) 和高地芽孢杆菌(Bacillus altitudinis) 组成,XY4 菌剂主要由短小芽孢杆菌(Bacillus pumilus)、解淀粉芽孢杆菌(Bacillusamyloliquefaciens) 和嗜热链霉菌(Streptomyces thermovulgaris) 组成。堆肥设接种自制菌剂CDS、XY4、商用菌剂EM 和无菌剂培养基对照(CK) 共4 个处理,以质量比为1.5% 的接种量接种。每日监测堆肥温度,分别在堆肥的第1、12、28、36 天进行样品采集,测定腐殖酸和黄腐酸含量、酶活性(脲酶、碱性磷酸酶、蔗糖酶和过氧化氢酶) 和腐熟指标(种子发芽指数、碳氮比和pH)。【结果】所有处理(CK、CDS、XY4 和EM) 的高温期(gt;50.0℃) 均持续超过20 天。在堆肥28 天后,CDS 和XY4 处理的堆肥浸提液处理的种子发芽指数分别达到85.1%、89.6%,显著高于CK 处理的73.8%,EM 处理与CK 处理无显著差异。XY4 处理堆肥结束时(36 天) 的C/N 值为14.6,显著低于CK 处理的19.1。第1~36 天CDS 和XY4 处理的黄腐酸含量始终高于CK (Plt;0.05),第36 天的腐殖酸含量也显著高于CK (Plt;0.05)。此外,CDS、XY4 处理较其他处理显示了较高的酶活性:XY4 处理第28 天的脲酶活性高达28.0 mg/(g·d),第12 天时的蔗糖酶活性达到97.6 mg/(g·d),显著高于其他处理组(Plt;0.05);CDS 处理第12 天和第28 天的碱性磷酸酶活性最高,分别为40.0 和32.1 mg/(g·d)。相关性和冗余分析显示,相较于CK,CDS 和XY4 处理中过氧化氢酶对腐殖酸形成具有重要作用(解释率分别为17.3% 和26.7%)。同时,CDS、XY4 和EM 处理腐殖酸与碱性磷酸酶活性呈显著负相关(Plt;0.05)。【结论】由短小芽孢杆菌(Bacillus pumilus)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens) 和嗜热链霉菌(Streptomyces thermovulgaris)组成的微生物菌剂(XY4),提升堆肥腐熟过程碱性磷酸酶和蔗糖酶活性的功能高于其他菌剂组合,因而加速了堆肥中有机物的快速分解和腐殖化,有效缩短了堆肥的腐熟周期,加快了堆肥的腐熟进程,显示了较好的市场前景。
关键词: 牛粪; 堆肥; 复合菌剂; 腐熟指标; 腐殖质; 酶活性
现代农业活动(大规模畜牧业和集约化作物生产) 产生了大量未被利用的固体和液体废物[1−2]。据统计,农业废弃物的上升速度为每年5% 至10%,年产量高达50 亿t[3]。其中,每年产生约15 亿t 畜禽粪便,但其综合利用率不到60%;尾菜(蔬菜废弃物) 的产量约为5 亿t,未利用比例超过90%[4];此外,种植100 kg 食用菌所需的原料,收获鲜菇后产生了60 kg 菌糠[5]。这些有机废弃物处理不当,不仅浪费资源,而且由于其富含有机物和营养物质,会对空气、土壤和水体等造成严重污染[6]。目前,堆肥技术被广泛认为是处理农业废弃物的一种实用且经济的方法,提高了资源化利用效率,减少了环境污染[7]。然而,单一物料堆肥存在含水率高、降解率低、碳氮比低等缺点。目前,对多组分物料混合堆肥进行了大量研究。多组分物料混合堆肥是一种利用多种有机废弃物混合堆肥的方法,堆肥产品具有生物利用度高和丰富的营养元素,其发酵分解能力优于单一堆肥[8]。然而,混合堆肥的过程较为复杂,涉及了多种物理、化学和微生物等因素的相互作用。微生物与其分泌的酶在堆肥中发挥着重要作用,将原始有机物质降解为简单物质,并将其合成为腐殖化合物[9]。虽然堆肥中原有的微生物对堆肥有降解作用,但畜禽粪便、菌糠和尾菜这类纤维素含量较高的有机废弃物,其堆肥进程均存在发酵温度上升缓慢、腐熟周期长等问题[10],这不仅降低了堆肥的农业价值,而且成为制约好氧堆肥处理农业废弃物商业化和产业化的关键因素[11]。因此,研究畜禽粪便、菌糠和尾菜等有机质降解的措施,对实现废弃物的资源化利用具有重要意义。
目前,接种外源微生物菌剂是优化堆肥技术的有效手段,具有代谢强度高、繁殖速度快等特点[12]。接种微生物菌剂能加快堆肥温度,并延长高温持续时间,提高pH 和种子发芽指数,促进有机质的碳化和腐殖化过程[13−14]。张祥永等[15]研究发现,接种菌剂增加了堆肥发酵的温度,延长了高温期,并且显著提高了酶(纤维素酶和蛋白酶) 活性。Pandey 等[16]研究表明,外源复合微生物接种有助于纤维素、半纤维素和木质素的分解,加速堆肥的腐殖化。李恕艳等[17]发现,在鸡粪中接种菌剂能增加堆肥腐殖质的含量,对其腐熟度和酶活性的提高有促进作用。上述研究结果表明,接种外源微生物菌剂能够提高堆肥效率并改善堆肥质量。由于堆肥原料的不同,接种的微生物菌剂也存在差异。研究表明,接种复合菌剂的效果通常优于单一菌剂[18]。目前,市售的微生物复合菌剂中很少有适应多种物料混合堆肥的菌剂,针对牛粪、菌糠渣和尾菜等原料的堆肥特性的复合菌剂研究和开发也相对较少。因此,本研究利用前期试验研究积累的降解菌,制备出两种自制的复合菌剂。以牛粪、菌糠渣和尾菜为堆肥材料,进行不同菌剂接种处理的对比研究,探讨自制菌剂对混合堆肥腐熟度、腐殖质含量和酶活性的影响,以及不同菌剂对堆肥发酵阶段动态变化的影响,旨在为接种外源复合菌剂促进混合堆肥高效腐熟提供理论支持。
1 材料与方法
1.1 试验材料
堆肥原料包括牛粪、菌糠和尾菜。其中,牛粪和菌糠渣来自甘肃省大行农业废弃物处理有限公司,尾菜则来自周边农村菜地。尾菜经过粉碎,碎片大小约为1~2 cm。所有堆肥原材料的物理化学特性参见相关文献[19]。
1.2 微生物菌剂
试验所用菌剂CDS 和XY4 由本课题组研制,CDS 菌剂包括皮氏不动杆菌(Acinetobacter pittii)、枯草芽孢杆菌粪便亚种(Bacillus subtilis subsp. Stercoris)和高地芽孢杆菌(Bacillus altitudinis),XY4 菌剂包括短小芽孢杆菌(Bacillus pumilus)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens) 和嗜热链霉菌(Streptomycesthermovulgaris)[19−20]。市售菌剂(EM) 由山东绿陇生物科技有限公司生产,主要菌种包括酵母菌、乳酸菌、放线菌等。
将菌剂接种于50.0 mL 蛋白胨纤维素液体培养基(peptone cellulose solution),该培养基配方为每升蒸馏水中含5.0 g 蛋白胨、5.0 g NaCl、2.0 g CaCO3、1 g 酵母粉,2.0×6.0 cm 滤纸条,于28.0℃ 下180 r/min摇床上培养5 天,制成有效活菌数均达1.0×108 CFU/mL数量级的菌液。
1.3 试验方法
堆肥试验于2021 年8—9 月在甘肃大行农业废弃物处理有限公司进行。牛粪、菌糠与尾菜按湿重比为5∶4∶1 混合混匀,初始C/N 约为30∶1,含水率约为60%。试验设计4 个堆肥接种处理,3 个菌剂CDS、XY4、EM 以质量比为1.5% 的接种量接种,以接种等量无菌剂为对照(CK)。将堆料充分混匀后堆制成长×宽×高=3.0 m×1.2 m×1.35 m 的条垛式堆,采用人工翻堆的形式,每4 天翻堆1 次,以保证供氧和堆料腐熟一致。
在堆肥的第1、12、28、36 天进行样品采集。将每个堆体分成4 段,在每段上层0~30 cm、中间层30~60 cm、下层60~90 cm 随机采集等量样品,充分混合后,取500.0 g 鲜样[21],分为两份:一份自然风干后用于测定腐熟指标和腐殖质含量;另一份在4.0℃ 下保存,用于测定堆肥中酶活性。
1.4 测定方法
1.4.1 堆体温度
利用针式数字温度仪(dot typedigital thermometer) 分别在堆肥顶部30、60 和90 cm 处进行测温,取平均值作为堆体温度,同时测定当日的环境气温,具体参考李昌宁等[21]的方法测定。
1.4.2 腐熟度、腐殖质含量和酶活性测定
种子发芽指数(germination index,GI) 的测定:堆肥样品与蒸馏水按1∶10 比例充分混合,取10.0 mL 滤液以3000 r/min 离心10 min,吸取5.0 mL 上清液于铺有滤纸的培养皿中,用蒸馏水作为对照,放置30 粒白菜种子,3 个重复,30.0℃ 培养4 天,计算种子的发芽率,并用游标卡尺测量种子的根长。根据下列公式,GI 计算方法[21]如下:
GI=(堆肥浸提液处理种子发芽率×种子根长)/(蒸馏水处理种子发芽率×种子根长)×100%
酸碱度(pH) 采用固液比1∶5 浸提,用酸度计测定[22];总有机碳(TOC) 采用重铬酸钾氧化法[23]测定,全氮(TN) 采用凯氏定氮法[23]测定。
腐殖酸和黄腐酸采用1000.0 mL 碱性磷酸钠和氢氧化钠溶液从10.0 g 样品提取,提取液在25.0℃ 下振荡24 h,然后12000 r/min 离心15 min,以0.45 μm滤膜过滤。重复上述步骤,直到提取液几乎无色,合并所有提取液。用盐酸调节腐殖质溶液pH 至1,在4.0℃ 下静置12 h,随后以12000 r/min 离心15 min,上清液为黄腐酸,沉淀为腐殖酸。沉淀用NaHCO3溶解得到腐殖酸,最后,采用重铬酸钾氧化法,分析腐殖酸和黄腐酸的总有机碳,以得到腐殖酸和黄腐酸含量[24]。计算公式为:
腐殖酸或黄腐酸=(V0−V)×c×3×10−3×分取倍数×100/(m×f)
式中:V0、V 分别为空白测定和样品测定所消耗的硫酸亚铁的体积(mL);c 为硫酸亚铁的浓度(mol/L);m 为称取干物质量(g);f 为含碳量系数(系数为1.742)。
堆肥中脲酶、碱性磷酸酶、蔗糖酶以及过氧化氢酶的活性参照关松荫[25]方法测定,脲酶活性测定以尿素为基质,根据酶促产物氨与苯酚钠−次氯酸钠作用生成蓝色的靛酚,采用比色法来测定脲酶活性;碱性磷酸酶活性的测定以磷酸苯二钠为基质,采用分光光度计比色法测定;根据堆肥因生化反应生成3-氨基-5-硝基水杨酸而呈橙黄色,采用分光光度计比色法测定蔗糖酶活性;过氧化氢酶活性采用高锰酸钾溶液滴定法测定。
1.5 数据处理与分析
采用Microsoft Excel 2010 进行数据整理;用SPSS26.0 软件进行单因素方差分析和LSD 法进行多重比较;并运用Canoco 5.0 进行冗余分析(redundancyanalysis,RDA),用Origin 2021 绘图。
2 结果与分析
2.1 菌剂对堆肥过程中温度的影响
由图1 可知,4 组处理的温度变化均经历了升温期、高温期、降温期和腐熟期4 个阶段,各处理高温期温度均达到50.0℃ 以上。堆肥第1 天,XY4 处理首先进入高温期,其他3 个处理于堆肥第2 天进入高温期。CK、CDS、XY4 和EM 处理的高温期分别维持了22、23、25 和26 天。由于堆肥过程中的翻堆,各处理高温期的温度变化幅度较大,主要集中在堆肥的4~2 2 天。上述结果表明,X Y 4 和EM 处理有利于加速堆肥升温和延长高温持续时间。
2.2 菌剂对堆肥腐熟指标的影响
2.2.1 菌剂对堆肥种子发芽指数(GI) 的影响
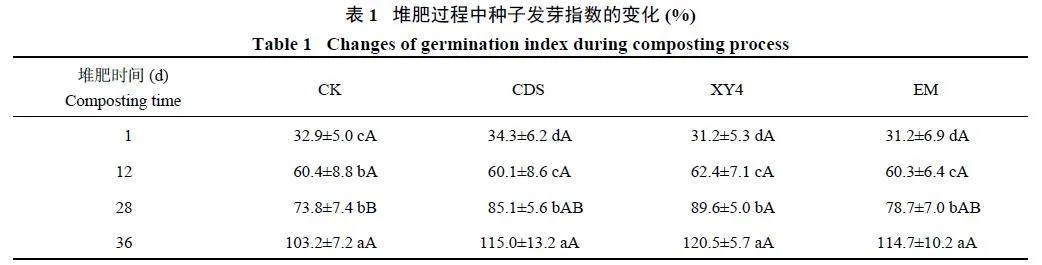
如表1 所示,各组的GI 值随堆肥时间延长均呈上升趋势,堆肥样品对植物的毒性逐渐降低。堆肥前12天,GI 值均小于80.0%;在28 天时,与CK 相比,XY4 处理显著提高了GI 值(Plt;0.05),且大于80.0%。这表明XY4 处理接种微生物菌剂可增强堆肥浸提液的促生能力。堆肥进行至第36 天,各处理GI 值均大于80.0%,表明已完全腐熟。
2.2.2 菌剂对堆肥pH 的影响
如图2 所示,前28 天堆肥的pH 呈现急剧上升至9.7~10.2,随后逐渐降低,在36 天时降至8.3~8.7,XY4 和EM 处理的pH 显著低于CK,符合国家有机肥标准要求。
2.2.3 菌剂对堆肥C/N 的影响
如表2 所示,各组堆肥C/N 随堆肥时间延长均呈下降趋势。在前28天,C/N 下降迅速,之后下降速度减缓并趋于稳定。CK、CDS、XY4 和EM 处理的C/N 分别从初始的约30∶1 降至36 天时的19.1、16.3、14.6 和15.4。其中,XY4 处理的C/N 较低(Plt;0.05),表明菌剂接种加速了堆肥中总有机碳的降解,促进了总氮含量相对增加,最终导致堆肥的C/N 降低。
2.2.4 菌剂对堆肥有机碳的影响
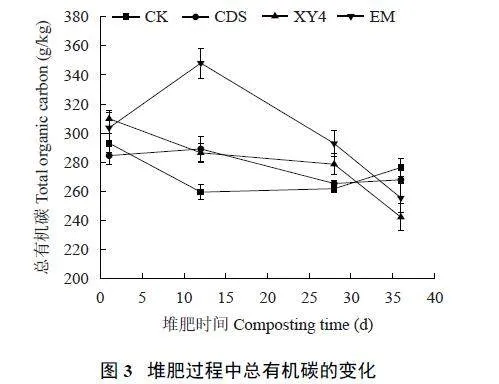
CDS 和EM 处理在堆肥第12 天后总有机碳含量开始明显下降,第28~36 天期间保持相对稳定(图3)。相比之下,XY4 和CK 处理在前12 天下降明显,之后下降速度逐渐减缓并趋于稳定。第36 天CK 总有机碳含量略微回升至276.4 g/kg,XY4 和EM 处理总有机碳含量分别为242.2 和255.5 g/kg。最终,XY4 处理的总有机碳含量最低,表明接种自制菌剂促进了总有机碳的转化。
2.3 接种菌剂对堆肥过程中腐殖质的影响
2.3.1 菌剂对堆肥后期腐殖化的影响
堆肥过程中各处理的腐殖酸含量均呈增加趋势(表3)。CDS 和XY4 处理在堆肥28~36 天,腐殖酸含量分别从25.5和25.7 g/kg 上升至28.0 和30.1 g/kg,表明在接种菌剂后28~36 天,CDS 和XY4 处理使腐殖酸含量增加。此外,第36 天,接种菌剂CDS、XY4 处理的腐殖酸含量均显著高于CK (Plt;0.05),这说明接种自制菌剂可以促进降温期和腐熟期腐殖酸的形成,加快堆肥有机质的腐殖化。
2.3.2 菌剂对堆肥过程中黄腐酸的影响
由(表3)可知,堆肥过程中各处理黄腐酸含量随堆肥时间延长呈现先上升后下降逐渐趋于稳定的趋势。堆肥第1~12 天,各处理的黄腐酸含量逐渐增加至最大值,其中XY4 处理含量最高,CK 处理最低,两者相差4.2 g/kg (Plt;0.05)。在12~36 天,CK、CDS、XY4和EM 处理黄腐酸含量分别从16.9、19.3、21.1 和20.1 g/kg 降至12.7、15.4、14.9 和14.0 g/kg,与CK 相比,CDS 和XY4 处理的黄腐酸含量较高(Plt;0.05)。这表明CDS 和XY4 处理接种菌剂后促进了整个堆肥过程中黄腐酸的形成。
2.4 接种菌剂对堆肥过程中酶活性的影响
2.4.1 菌剂对堆肥中脲酶活性的影响
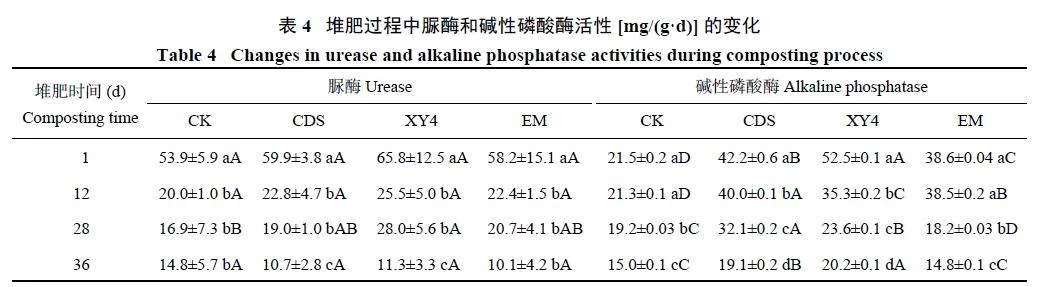
在堆肥过程中,随堆肥时间延长脲酶活性逐渐降低(表4)。CK、CDS、XY4 和EM 处理的脲酶活性分别为14.8~53.9、10.7~59.9、11.3~65.8 和10.1~58.2 mg/(g·d)。在堆肥第1、12 和36 天,CK、CDS、XY4 和EM处理中脲酶活性没有显著差异(Pgt;0.05)。然而,在第28 天,XY4 处理的脲酶活性显著高于CK (Plt;0.05),为28.0 mg/(g·d),表明XY4 处理在降温期有助于提高脲酶活性。
2.4.2 菌剂对堆肥过程碱性磷酸酶活性的影响
各处理的碱性磷酸酶活性随堆肥进程呈不断下降趋势,CK、CDS、XY4 和EM 处理的碱性磷酸酶活性分别为15.0~21.5、19.1~42.2、20.2~52.5 和14.8~38.6 mg/(g·d) (表4)。堆肥第12 和第28 天,CDS 处理的碱性磷酸酶活性分别达到最高值(Plt;0.05),分别为40.0 和32.1 mg/(g·d)。在第1 天(升温期) 和第36 天,XY4 处理的碱性磷酸酶活性分别为52.5 和20.2 mg/(g·d) (Plt;0.05),说明在升温期和腐熟期XY4 处理有效提高了碱性磷酸酶活性。
2.4.3 菌剂对堆肥过程中蔗糖酶活性的影响
各处理蔗糖酶活性随堆肥时间延长呈先下降后趋于平缓,CK、CDS、XY4 和EM 处理的蔗糖酶活性分别为44.6~74.3、44.1~118.2、41.7~98.6 和44.6~97.1 mg/(g·d) (表5)。在堆肥第1 和28 天,CDS 处理的蔗糖酶活性最高(Plt;0.05),分别为118.2 和64.5 mg/(g·d)。第12 天,XY4 处理的蔗糖酶活性显著高于其他处理(Plt;0.05),为97.6 mg/(g·d),表明CDS 处理在升温期和降温期提升蔗糖酶活性的效果显著,XY4 处理在高温期提升蔗糖酶活性的效果显著。
2.4.4 菌剂对堆肥过程中过氧化氢酶的影响
堆肥各处理的过氧化氢酶活性随堆肥时间延长呈先下降后上升逐步趋于稳定的趋势(表5)。在堆肥第1 天,CK、CDS、XY4 和EM 处理过氧化氢酶活性分别为5.3、6.9、7.4 和5.7 mg/(g·d) (Plt;0.05),表明接种菌剂有助于增加堆肥的过氧化氢酶活性,其中XY4效果较好。第12 天,CK、CDS、XY4 和EM 处理的过氧化氢酶活性均急剧下降。第36 天,CDS、XY4、EM 处理的过氧化氢酶活性较CK 显著降低(Plt;0.05),其中XY4 处理的过氧化氢酶活性最低,为5.0 mg/(g·d)。
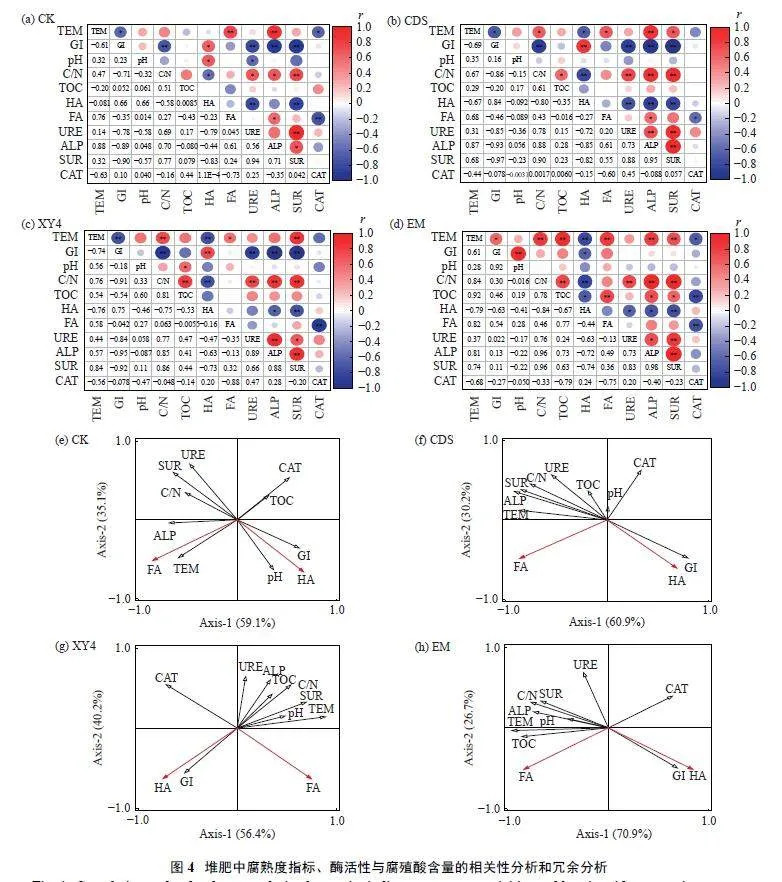
2.5 腐熟度指标、酶活性与腐殖质之间的相互关系
通过相关性与冗余分析,分别对4 个处理的腐熟指标、腐殖质和酶活性进行分析(图4 a~h)。不同菌剂致使腐殖酸和黄腐酸形成的主要驱动因素存在差异,各个处理RDA 分析前两主轴的总解释率在91.1%~97.6%。相比CK,CDS 和XY4 处理中过氧化氢酶对腐殖酸的形成有重要作用(解释率分别为17.3% 和26.7%);相比CK 和CDS 处理,XY4 和EM 处理温度与脲酶活性是腐殖酸形成的主要解释变量。除CK 外,CDS、XY4 和EM 处理蔗糖酶与温度显著正相关(Plt;0.05) (图4b、c、d)。同时,CDS和EM 处理腐殖酸也与碱性磷酸酶活性均呈显著负相关(Plt;0.05) (图4b、d)。
3 讨论
3.1 接种微生物菌剂对堆肥温度和腐熟指标的影响
温度是堆肥中各生理生化过程顺利进行的基础,也是决定堆肥成功的关键因素[26]。本研究表明,将XY4 菌剂(含有芽孢杆菌) 接种到XY4 处理可导致温度峰值最高,并延长高温期。Zhao 等[27]研究发现,芽孢杆菌菌剂接种能促进堆肥中的纤维素和果胶等有机物的降解,形成的营养物质也使微生物大量繁殖,产生大量热量。此外,高温期堆肥翻堆,各处理的温度变化幅度较大,但堆肥回温较快,究其原因可能是翻堆改变了堆肥的孔隙度,并增加其中的氧气含量,从而提升微生物的代谢效率,加速了堆肥热量的产生[21]。
种子发芽指数(GI) 可以通过评估植物毒性来估计堆肥腐熟,GIgt;80.0% 表明堆肥对植物生长毒害较低,这是堆肥腐熟并实现无害化的基本要求之一[28]。本研究中XY4、EM 处理发酵腐熟时显著提高了GI,可能是由于接种菌剂后加速堆肥中有机物的降解和无害化,使其具有良好的稳定性和较低的植物毒性,有助于植物的生长发育[22]。pH 值是影响堆肥腐熟过程的关键因素之一,可以调节微生物的活性以及碳、氮等营养物质的转化,通常堆肥腐熟后pH值在8.0~9.0[29−30]。堆肥结束时,较堆肥初期pH 均有所增加,可能的原因是接种菌剂后堆肥腐熟期的有机物分解加快,同时产生丰富的可挥发性有机酸,如乙酸、丁酸等[21]。C/N 是评价堆肥成熟度和稳定性的重要参数,堆肥腐熟最终的C/N 约为15~20[31]。李季等[32]对微生物群落组成与理化指标间的相关性进行研究后,发现厚壁菌门在低C/N 堆肥中占主导地位,其中以芽孢杆菌最为活跃。本研究中XY4 处理的C/N 降幅最大,可能其加速了氨态氮被转化为亚硝酸盐和硝酸盐,氮素水平升高,最终使堆肥C/N 下降。堆肥是微生物降解有机废弃物的生理生化过程。在这一过程中,碳具有重要作用,既是微生物的能量来源,也是有机物分解的基本元素[33]。本研究发现,接种菌剂后堆肥的有机碳含量减少,分解速度加快,其中XY4 处理的效果最为显著,分解较为彻底。王义祥等[34]研究发现,在堆肥中有机碳会被降解,而碳的转化受到堆肥原料、温度、含水率和微生物等多种因素的共同影响,其中微生物对这一过程的影响最为显著,通过接种纤维素降解菌株,可以促进有机碳更有效地腐殖化,这与本研究所得结果基本一致。
3.2 接种微生物菌剂对堆肥腐殖质的影响
腐殖酸和黄腐酸是腐殖质的主要组成部分。He等[35]研究发现,接种菌剂的堆肥处理提高了微生物木质纤维素的分解能力,使合成腐殖酸的化合物增多,致使腐殖酸含量升高。李昌宁等[21]研究表明,在猪粪堆肥中接种菌剂的腐熟期能促进腐殖酸的芳香结构增加,并被微生物利用再生,合成大量稳定的腐殖酸。在接种菌剂的堆肥升温阶段,与黄腐酸合成和分解相关的微生物和土著微生物差异不显著,但在高温期和降温期增加了黄腐酸含量[23]。本研究表明,CDS 和XY4 处理在堆肥第1 天腐殖酸含量显著高于CK 和EM 处理;在28~36 天,CDS、XY4 处理在堆肥初期促进腐殖化作用,提高了腐殖酸含量。1~12 天,4 组处理黄腐酸含量均达到最大值,其中XY4 处理最高,提高了高温期堆肥中黄腐酸含量,这与上述研究结果基本一致。此外,在第36天,接种菌剂处理腐熟期黄腐酸含量较高,这主要是因为黄腐酸含量的变化受到矿化和腐殖化平衡效应的影响,其分子量小、结构简单、容易被微生物降解,但接种菌剂可以使黄腐酸合成大于分解,致使黄腐酸含量增加[23, 36]。
3.3 接种微生物菌剂对堆肥酶活性的影响及与堆肥腐熟的相互关系
在堆肥中脲酶是氨化过程中的关键酶,且氨化是硝化作用的关键和限速步骤[37]。4 个堆肥处理脲酶活性随堆肥时间延长均呈逐渐下降趋势。其中,在堆肥1~12 天,各处理脲酶活性较高,但相较于CK,接种菌剂不能有效提高堆肥升温期和高温期脲酶活性,这可能是堆肥在升温期和高温期含有大量的含氮有机物,致使脲酶活性较高,但随着温度的升高,含氮有机物分解生成氨,抑制了脲酶活性,此外高温也会使脲酶菌群死亡,从而降低脲酶活性[37]。在第28 天,降温期XY4 处理提高了脲酶活性,这可能是接种菌剂提高了XY4 处理降温期脲酶相关菌群的代谢,加快含氮有机物的矿化和分解速度,从而提高了脲酶活性[12]。碱性磷酸酶对堆肥中含磷有机物具有分解和矿化作用[38]。本研究中,碱性磷酸酶活性随堆肥时间延长呈逐渐降低的趋势,这与堆肥在升温期存在大量有机磷化合物有关,提高了堆肥中碱性磷酸酶的活性,并增加了有机磷的转化[39]。随着堆肥的不断发酵腐熟,有机磷被分解完全,导致碱性磷酸酶活性不断降低。在第12 和28天,CDS 处理提高了高温期和降温期碱性磷酸酶活性,究其原因可能是接种菌剂引起的耐高温菌种丰度发生变化,导致碱性磷酸酶活性升高,从而影响了有机磷的转化途径[40]。
蔗糖酶能够分解蔗糖,生成葡萄糖、果糖等小分子物质,这些小分子物质恰好是微生物繁殖所需的碳源[37]。在堆肥1~28 天,CDS、XY4 与EM 处理的蔗糖酶活性呈下降趋势,但均高于CK,可能是接种菌剂促进了堆肥中蔗糖酶的协同代谢,进而提高了堆肥过程中的代谢活性[41]。此外,在堆肥第12天,XY4 处理的蔗糖酶活性为97.6 mg/(g·d),有效提高了高温期蔗糖酶活性。Li 等[11]研究发现,在猪粪堆肥中接种芽孢杆菌等微生物能更快到达高温期,并提高蔗糖酶活性。XY4 处理的蔗糖酶贡献度高达59.4%,说明在堆肥中蔗糖酶起着至关重要的作用,同时,CDS、XY4 和EM 处理蔗糖酶与温度、C/N 均呈极显著正相关,较高的温度有助于激发蔗糖酶活性,提高堆肥腐熟度,促进有机质的降解[37]。过氧化氢酶主要反映堆肥有机物的降解强度和对微生物的毒性影响[ 4 2 − 4 3 ]。本研究表明,堆肥第1 天,CK、CDS、XY4 和EM 处理过氧化氢酶活性分别为5.3、6.9、7.4 和5.7 mg/(g·d),表明接种菌剂有助于增加堆肥的过氧化氢酶活性,其中XY4 处理效果较好。研究表明,接种菌剂能加速堆肥微生物新陈代谢,分解有机物(主要是木质素),使堆肥有机物的转化率升高[12, 44]。此外,XY4 处理中较高的过氧化氢酶活性也降低了过氧化氢对微生物的毒害,促进作物生长繁殖。第36 天,XY4 处理的过氧化氢酶活性最低,为5.0 mg/(g·d),说明能被微生物利用的可降解有机化合物减少,过氧化氢酶的活性也随之降低[45],从而反映出接种菌剂加速了堆肥的快速腐熟。
微生物分泌的各种胞外酶在堆肥中发挥着重要作用,几乎参与了养分循环中所有生化反应[46]。在堆肥过程中监测酶活性可以获取与腐殖质形成相关的信息。有机物在水解酶(蔗糖酶、淀粉酶等) 的作用下进行矿化反应,而在氧化酶的作用下进行腐殖化反应[47]。齐熙平[48]研究表明,接种外源菌剂时,碱性磷酸酶、过氧化物酶是影响腐殖酸组分变化的主要因素,且腐殖酸主要通过类蛋白和类木质纤维素缩合来形成,同时碱性磷酸酶对各处理腐殖酸的形成都有影响,这与本研究的结果相似。
4 结论
包含短小芽孢杆菌(Bacillus pumilus)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens) 和嗜热链霉菌(Streptomyces thermovulgaris) 的菌剂(XY4) 较其他菌剂更有效地提高了堆肥中的碱性磷酸酶和蔗糖酶活性,促进有机质更快分解,腐殖化程度更高,缩短堆肥的腐熟时间,因此,具有很好的市场应用前景。
参 考 文 献:
[ 1 ]Wu D, Wei Z M, Qu F T, et al. Effect of Fenton pretreatmentcombined with bacteria inoculation on humic substances formationduring lignocellulosic biomass composting derived from ricestraw[J]. Bioresource Technology, 2020, 303: 122849.
[ 2"]Greff B, Szigeti J, Nagy Á, et al. Influence of microbial inoculants onco-composting of lignocellulosic crop residues with farm animalmanure: A review[J]. Journal of Environmental Management, 2022,302: 114088.
[ 3 ] Xing W N, Liu Q, Wang J Y, et al. High selectivity and reusability of biomass–based adsorbent for chloramphenicol removal[J].Nanomaterials (Basel), 2021, 11(11): 2950.
[ 4 ]王宇蕴, 赵兵, 马丽婷, 等. 堆肥腐殖化过程及微生物驱动机制[J].生物技术通报, 2022, 38(5): 22−28.
Wang Y Y, Zhao B, Ma L T, et al. Humification process andmicrobial driving mechanism of composting[J]. BiotechnologyBulletin, 2022, 38(5): 22−28.
[ 5 ]陈智坤, 路强强, 任英英, 等. 蘑菇渣腐熟微生物组方研究[J]. 农业资源与环境学报, 2019, 36(4): 494−501.
Chen Z K, Lu Q Q, Ren Y Y, et al. Study on microbiological formulaof spent mushroom compost[J]. Journal of Agricultural Resourcesand Environment, 2019, 36(4): 494−501.
[ 6 ]Ren X N, Wang Q, Awasthi M K, et al. Improvement of cleanercomposting production by adding diatomite: From the nitrogenconservation and greenhouse gas emission[J]. Bioresource Technology,2019, 286: 121377.
[ 7 ]Chen M L, Huang Y M, Liu H J, et al. Impact of different nitrogensource on the compost quality and greenhouse gas emissions duringcomposting of garden waste[J]. Process Safety and EnvironmentalProtection, 2019, 124: 326−35.
[ 8 ]Qian X Y, Shen G X, Wang Z Q, et al. Co-composting of livestockmanure with rice straw: Characterization and establishment ofmaturity evaluation system[J]. Waste Management, 2014, 34(2):530−535.
[ 9 ]Franke-Whittle I H, Confalonieri A, Insam H, et al. Changes in themicrobial communities during co-composting of digestates[J]. WasteManagement, 2014, 34(3): 632−641.
[10]Kuhad R C, Singh A, Eriksson K E. Microorganisms and enzymesinvolved in the degradation of plant fiber cell walls[J]. Advances inBiochemical Engineering/Biotechnology, 1997, 57: 45−125.
[11]Li C N, Li H Y, Yao T, et al. Effects of microbial inoculation onenzyme activity, available nitrogen content, and bacterial successionduring pig manure composting[J]. Bioresource Technology, 2020,306: 123167.
[12]Zhu L, Zhao Y X, Yao X W, et al. Inoculation enhances directionalhumification by increasing microbial interaction intensity in foodwaste composting[J]. Chemosphere, 2023, 322: 138191.
[13]曾金樱, 吴榕贵, 唐秀媚, 萧政祯. 不同微生物菌剂对鸡粪堆肥的影响研究[J]. 黑龙江畜牧兽医, 2023, (20): 57−63.
Zeng J Y, Wu R G, Tang X M, Xiao Z Z. Study on the effects ofdifferent microbial agents on chicken manure composting[J].Heilongjiang Animal Science and Veterinary Medicine, 2023, (20):57−63.
[14]王佳, 刘瑞生, 徐建峰, 等. 不同微生物菌剂组合对猪粪好氧堆肥发酵效果的影响[J]. 中国饲料, 2023, (12): 9−13.
Wang J, Liu R S, Xu J F, et al. Effect of different microbial agentcombinations on the fermentation of pig[J]. China Feed, 2023, (12):9−13.
[15]张祥永, 任静, 马彦霞, 郁继华. 微生物菌剂对牛粪堆肥中酶活性的影响[J]. 甘肃农业大学学报, 2016, 51(3): 65−71.
Zhang X Y, Ren J, Ma Y X, Yu J H. Effects of microbial agents onenzymes activities during cow manure composting[J]. Journal of Gansu Agricultural University, 2016, 51(3): 65−71.
[16]Pandey S, Singh S, Yadav A N, et al. Phylogenetic diversity andcharacterization of novel and efficient cellulase producing bacterialisolates from various extreme environments[J]. Bioscience,Biotechnology, and Biochemistry, 2013, 77(7): 1474−1480.
[17]李恕艳, 李吉进, 张邦喜, 等. 菌剂对鸡粪堆肥腐殖质含量品质的影响[J]. 农业工程学报, 2016, 32(增刊2): 268−274.
Li S Y, Li J J, Zhang B X, et al. Influence of inoculants on contentand quality of humus during chicken manure composting[J].Transactions of the Chinese Society of Agricultural Engineering,2016, 32(S2): 268−274.
[18]Mikesková H, Novotny C, Svobodová K. Interspecific interactions inmixed microbial cultures in a biodegradation perspective (review)[J].Applied Microbiology and Biotechnology, 2012, 95(4): 861−870.
[19]陈鑫, 李昌宁, 肖金玉, 等. 接种不同微生物菌剂对牛粪好氧堆肥腐熟的影响[J]. 草原与草坪, 2023, 43(6): 58−65.
Chen X, Li C N, Xiao J Y, et al. Effect of inoculation of differentmicrobial agents on the maturation of cattle manure by aerobiccompost[J]. Grassland and Turf, 2023, 43(6): 58−65.
[20]Li C N, Li H Y, Yao T, et al. Microbial inoculation influencesbacterial community succession and physicochemical characteristicsduring pig manure composting with corn straw[J]. BioresourceTechnology, 2019, 289: 121653.
[21]李昌宁, 苏明, 姚拓, 等. 微生物菌剂对猪粪堆肥过程中堆肥理化性质和优势细菌群落的影响[J]. 植物营养与肥料学报, 2020, 26(9):1600−1611.
Li C N, Su M, Yao T, et al. Effects of microbial inoculation oncompost physical and chemical propertiesand dominant bacterialcommunities during composting of pig manure[J]. Journal of PlantNutrition and Fertilizers, 2020, 26(9): 1600−1611.
[22]沈丹青, 于鑫, 韩捷, 等. 芽孢杆菌固体菌剂对园林绿化废弃物堆肥的影响[J]. 西北农林科技大学学报(自然科学版), 2024, 52(1):127−135.
Shen D Q, Yu X, Han J, et al. Effect of Bacillus solid bacterialinocula on compost of green waste[J]. Journal of Northwest A amp; FUniversity (Natural Science Edition), 2024, 52(1): 127−135.
[23]鲍士旦. 土壤农化分析(第三版)[M]. 北京: 中国农业出版社, 2000.
Bao S D. Soil agricultural chemistry analysis (3rd edition)[M].Beijing: China Agriculture Press, 2000.
[24]Li C N, Li H Y, Yao T, et al. Effects of swine manure composting bymicrobial inoculation: Heavy metal fractions, humic substances, andbacterial community metabolism[J]. Journal of Hazardous Materials,2021, 415: 125559.
[25]关松荫. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986.
Guan S Y. Soil enzymes and their research methods[M]. Beijing:Agriculture Press, 1986.
[26]韩丽娜, 王泽槐, 李建国. 接种外源微生物菌剂对香蕉茎秆堆肥的影响[J]. 环境工程学报, 2012, 6(11): 4215−4222.
Hang L N, Wang Z H, Li J G, et al. Effects of inoculating microbeson banana pseudostem composting[J]. Chinese Journal of EnvironmentalEngineering, 2012, 6(11): 4215−4222.
[27]Zhao Y, Lu Q, Wei Y Q, et al. Effect of actinobacteria agentinoculation methods on cellulose degradation during composting based on redundancy analysis[J]. Bioresource Technology, 2016,219: 196−203.
[28]Li Y, Li J Z, Li S S, et al. Effects of different composting methods onEnteromorpha: Maturity, nutrients, and organic carbon transformation[J]. Journal of Cleaner Production, 2022, 380: 135073.
[29]Wei Y Q, Zhao Y, Fan Y Y, et al. Impact of phosphate-solubilizingbacteria inoculation methods on phosphorus transformation andlong–term utilization in composting[J]. Bioresource Technology,2017, 241: 134−141.
[30]García C, Hernández T, Costa F, Pascual J A. Phytotoxicity due tothe agricultural use of urban wastes. Germination experiments[J].Journal of the Science of Food and Agriculture, 1992, 59(3): 313−319.
[31]Guo R, Li G X, Jiang T, et al. Effect of aeration rate, C/N ratio andmoisture content on the stability and maturity of compost[J].Bioresource Technology, 2012, 112: 171−178.
[32]李季, 王禄山. 堆肥微生物: 过去, 现在和未来[J]. 生物技术通报,2022, 38(5): 1−3.
Li J, Wang L S. Composting microbes: Past, present and future[J].Biotechnology Bulletin, 2022, 38(5): 1−3.
[33]张国言. 高效纤维素降解菌株筛选及其复合微生物菌剂在堆肥中的应用研究[D]. 山东泰安: 山东农业大学硕士学位论文, 2023.
Zhang G Y. Screening of high efficient cellulose degrading strainsand application of its compound microbial agents in composting[D].Tai'an, Shandong: MS Thesis of Shandong Agricultural University,2023.
[34]王义祥, 高凌飞, 叶菁, 等. 菌渣垫料堆肥过程碳素物质转化规律[J]. 农业工程学报, 2016, 32(增刊2): 292−296.
Wang Y X, Gao L F, Ye J, et al. Change of carbon substancecharacteristics during composting of waste packing and funguschaff[J]. Transactions of the Chinese Society of AgriculturalEngineering, 2016, 32(S2): 292−296.
[35]He J, Zhu N M, Xu Y S, et al. The microbial mechanismsof enhanced humification by inoculation with Phanerochaetechrysosporium and Trichoderma longibrachiatum during biogasresidues composting[J]. Bioresource Technology, 2022, 351: 126973.
[36]Wu J Q, Zhao Y, Qi H S, et al. Identifying the key factors that affectthe formation of humic substance during different materialscomposting[J]. Bioresource Technology., 2017, 244: 1193−1196.
[37]Yu H Y, Xie B T, Khan R, et al. The changes in functional markergenes associated with nitrogen biological transformation duringorganic-inorganic co-composting[J]. Bioresource Technology, 2020,295: 122197.
[38]许淑妮, 冯晓璐, 陈延玲, 李赟. 添加玉米秸秆对胡萝卜尾菜堆肥过程的影响[J]. 农业工程学报, 2023, 39(11): 230−237.
Xu S N, Feng X L, Chen Y L, Li Y. Effects of adding maize stoveron the process of carrot waste composting[J]. Transactions of theChinese Society of Agricultural Engineering, 2023, 39(11): 230−237.
[39]Villarreal-Chiu J, Quinn J, Mcgrath J. The genes and enzymes ofphosphonate metabolism by bacteria, and their distribution in themarine environment[J]. Frontiers in Microbiology, 2012, 3: 19.
[40] Marinari S, Masciandaro G, Ceccanti B, Grego S. Influence of organic and mineral fertilisers on soil biological and physicalproperties[J]. Bioresource Technology, 2000, 72(1): 9−17.
[41]Wan W J, Wang Y, Tan J D, et al. Alkaline phosphatase-harboringbacterial community and multiple enzyme activity contribute tophosphorus transformation during vegetable waste and chickenmanure composting[J]. Bioresource Technology, 2020, 297: 122406.
[42]Sun Q H, Wu D, Zhang Z, et al. Effect of cold-adapted microbialagent inoculation on enzyme activities during composting start-up atlow temperature[J]. Bioresource Technology, 2017, 244: 635−640.
[43]Ukalska–Jaruga A, Smreczak B, Klimkowicz-Pawlas A. Soil organicmatter composition as a factor affecting the accumulation of polycyclicaromatic hydrocarbons[J]. Journal of Soils and Sediments, 2019,19(4): 1890−1900.
[44]Hammel K E. Extracellular free radical biochemistry of ligninolyticfungi[J]. New Journal of Chemistry, 1996, 20: 195−198.
[45]吕药灵, 吴敏, 贾玉蕾, 等. 堆肥过程中理化性质和酶活性的演变[J]. 世界生态学, 2016, 5(1): 8−16.
Lü Y L, Wu M, Jia Y L, et al. Evolution of physico-chemical properties and enzymatic activity during composting[J]. InternationalJournal of Ecology, 2016, 5(1): 8−16.
[46]Wang N Y, Ren L H, Zhang J C, et al. Activities of functionalenzymes involved in C, N, and P conversion and their stoichiometryduring agricultural waste composting with biochar and biogas residueamendments[J]. Bioresource Technology, 2021, 345: 126489.
[47]白玲, 李倩, 邓芸, 等. 沼渣与餐厨废弃物、牛粪联合堆肥的腐殖化进程研究[J]. 农业机械学报, 2019, 50(8): 331−338.
Bai L, Li Q, Deng Y, et al. Humification process of biogas residuecombined with food waste and cattlemanure co-composting[J].Transactions of the Chinese Society for Agricultural Machinery,2019, 50(8): 331−338.
[48]齐熙平. 不同添加剂对含沙牛粪好氧堆肥腐殖化的影响机制[D].陕西杨凌: 西北农林科技大学硕士学位论文, 2023.
Qi X P. The influence mechanism of different additives on thehumification of aerobic composting of sand containing cow manure[D]. Yangling, Shaanxi: MS Thesis of North West Agriculture andForestry University, 2023.
基金项目:高校科研创新平台重大培育项目(2024CXPT-07);甘肃省科技厅技术创新引导计划项目(23CXNE0029) ;甘肃农业大学科研启动经费(GAU-KYQD-2022-01)。
