TMEM206基因敲除小鼠小脑组织中差异表达基因的筛选及生物学功能分析
2024-04-21陆盈孙顺昌
陆盈,孙顺昌,2
1 上海交通大学医学院附属瑞金医院检验科,上海 200025;2 上海交通大学医学院附属瑞金医院无锡分院检验科
质子活化型Cl-(proton-activated Cl-, PAC)通道是新发现的一种Cl-通道,由3分子跨膜蛋白206(transmembrane protein 206, TMEM206)单体组成,可将Cl-转运至细胞内,但其在细胞中的功能尚未完全清楚[1]。体外研究[2]显示,PAC通道在pH值降至6.2时开始激活,PAC通道在酸性条件下被激活后,将Cl-从内质网或分泌囊泡内排出,防止内体腔过酸,病理条件下如缺血性卒中和肿瘤组织内,pH值较低,PAC通道也可被活化,可将Cl-转运至细胞内,细胞容积增大,甚至肿胀死亡[3]。TMEM206蛋白氨基酸序列在脊椎动物中高度保守,且在人体各组织中广泛分布,以脑组织中相对含量最高,提示其或许有类似管家基因的功能。TMEM206蛋白含2个跨膜螺旋和1个富含β折叠的胞外结构域,TMEM206蛋白的N-端和C-端位于胞内侧[4]。我们的前期研究发现,ataxin-1蛋白与TMEM206蛋白存在相互作用,小脑组织浦肯野细胞中ataxin-1蛋白的编码基因ATXN1基因突变是1型脊髓小脑型共济失调(spinocerebellar ataxia type 1, SCA1)发病的分子基础,我们推测TMEM206蛋白可能与SCA1的发病有关,因此有必要探讨TMEM206基因的功能。2022年10月—2023年5月,我们通过CRISPR/Cas9基因编辑技术敲除小鼠的TMEM206基因,筛选TMEM206基因敲除小鼠小脑组织中的差异表达基因并进行生物学功能分析,探讨TMEM206基因的功能。
1 材料与方法
1.1 C57BL/6系TMEM206基因敲除纯合小鼠的制备 TMEM206基因敲除鼠采用C57BL/6系小鼠制备,基因敲除鼠及其组织制备符合实验动物福利与伦理原则和标准。基因敲除使用CRISPR/Cas9基因编辑技术,敲除TMEM206基因的外显子2~外显子5片段(见图1)。敲除路径:向单细胞期受精卵同时注射Cas9蛋白和靶向外显子2和外显子5的单向导RNA。单向导RNA序列如下,gRNA1:5'-CTAGCTTCT-GCTGATTACCC-3';gRNA2:5'-CCTTAAAGATGTACCCCAAG-3';gRNA3:5'-CAAGAAGGCTCACGGACCAC-3';gRNA4:5'-AGTGAAGCCTGAAGCTCGGC-3'。TMEM206基因敲除小鼠委托江苏集萃药康生物科技股份有限公司制备,TMEM206基因敲除突变通过PCR扩增和Sanger测序验证,TMEM206基因敲除纯合鼠通过TMEM206基因敲除杂合鼠配殖获得。
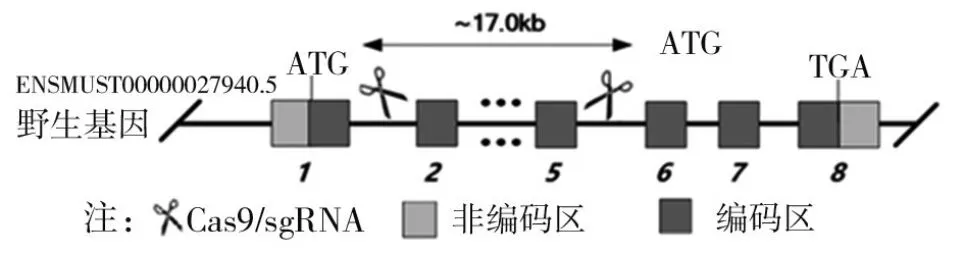
图1 TMEM206基因敲除示意图
1.2 TMEM206基因敲除纯合小鼠和同窝野生小鼠的饲养、体质量变化 挑选TMEM206基因敲除纯合小鼠和同窝野生小鼠雌、雄各5只,常规饲养,在第49~52周称取小鼠体质量,比较TMEM206基因敲除纯合小鼠和同窝野生小鼠的体质量变化,结果显示TMEM206基因敲除纯合小鼠和同窝野生小鼠体质量差异无统计学意义(P均>0.05),提示TMEM206基因敲除不影响小鼠的生长发育。
1.3 TMEM206基因敲除小鼠小脑组织RNA测序文库的构建 分别取3只8周龄雌性TMEM206基因敲除纯合小鼠和3只同窝雌性野生小鼠,腹腔注射戊巴比妥钠液麻醉,用预冷切片液心脏灌流,迅速断头,剪开颅骨取出全脑,再分离小脑组织。3只TMEM206基因敲除纯合鼠分别取等量小脑组织混合为1管,3只野生小鼠也同样取等量小脑组织混合为1管。每管组织分别用Invitrogen公司的TRIzol试剂提取总RNA,经测定浓度和纯度后,构建测序文库,用于转录组测序。
1.4 TMEM206基因敲除小鼠小脑组织中差异表达基因的筛选 小脑组织转录组测序委托苏州金唯智生物科技有限公司完成。转录组测序数据经过滤后显示,TMEM206基因敲除纯合鼠小脑组织测序平均长度为148.25 bp,产生89495253测序reads,总碱基数为13268086226;野生C57BL/6J小鼠小脑组织测序平均长度为148.29 bp,产生82118098测序reads,总碱基数为12196692913。测序数据质量评估采用FastQC(v0.10.1)软件分析,结果显示TMEM206基因敲除纯合鼠和野生鼠小脑测序碱基质量分数均在35以上,即碱基位置测序正确率在99.9%以上;两个测序样本绝大多数碱基序列的平均Phred碱基质量分数峰值均>30,提示测序序列质量较好。基因差异分析使用Cuffdiff(v2.2.1)软件,依据测序数据counts值,通过EBseq算法筛选差异表达基因,差异倍数>2.0或<0.5,同时错误发现率<0.05时,视为差异表达基因。
1.5 TMEM206基因敲除小鼠小脑组织中差异表达基因的生物信息学分析 使用DAVID数据库(https://david.ncifcrf.gov/)对差异表达基因进行基因属性(gene ontology, GO)和京都基因和基因组百科全书(kyoto encyclopedia of genes and genomes,KEGG)通路富集分析。
2 结果
2.1 TMEM206基因敲除小鼠小脑组织中差异表达基因的筛选结果 共筛选出474个差异表达基因,与正常小脑组织相比,TMEM206基因敲除小鼠小脑组织中上调的基因265个、下调的基因209个。其中表达量上调最高的10个基因由高到低依次为Acp2、Gzma、Tfe3、Zfp518a、Ppp1r26、Top3b、Mki67、Zranb1、Top2a、Chd9,表达量下调最高的10个基因由高到低依次为Ppp1r26、Gak、Slc15a2、Coch、Gm49325、Resf1、Mid1-ps1、Ptpre、Myo7a、Ntrk2。以上结果提示TMEM206基因敲除影响了小鼠小脑组织的基因表达谱。
2.2 TMEM206基因敲除小鼠小脑组织中差异表达基因的生物学功能分析结果 GO功能分析结果显示,差异表达基因的表达产物定位于细胞外膜和主要组织相容性复合体Ⅰ类蛋白(major histocompatibility complex Ⅰ, MHC Ⅰ)较多,这些基因表达产物功能属性主要归类于结合转录因子、脂类分子及抗原分子等;差异表达基因参与的生物学过程众多,如DNA损伤的细胞反应、细胞分化选择、骨质矿化、BMP信号通路负调控、内皮细胞增殖负调控、血管收缩正调控等。KEGG通路富集分析结果显示,差异表达基因在分子属性上多为细胞黏附分子和肿瘤坏死因子路径的分子,功能上主要与细胞吞噬和衰老有关,此外还与人乳头瘤病毒、Ⅰ型人类T淋巴细胞白血病病毒、单纯疱疹病毒、ⅤⅢ型人疱疹病毒及EB病毒等感染有关。
3 讨论
脊髓小脑型共济失调(spinocerebellar ataxia,SCA)是一类遗传性共济失调,病变主要累及小脑、脑干及脊髓[5]。目前已发现的SCA类型有30多型,其中SCA1、SCA2、SCA3、SCA6、SCA7等型均由致病基因编码区CAG(编码谷氨酰胺)三核苷酸重复序列的重复数增加所致,因此又称为多聚谷氨酰胺病或poly Q病[6-7]。SCA1的致病基因为ATXN1,基因表达产物为ataxin-1,但发病机制仍不清楚[8]。前期通过稳定同位素标记氨基酸免疫沉淀分析和蛋白免疫共沉淀,我们发现TMEM206与ataxin-1存在相互作用,提示TMEM206或许参与SCA1的发病[9]。因此,有必要探讨TMEM206基因在小脑组织中的功能。研究[10]显示,PAC通道失活时,可保护酸诱导的神经元死亡,活化时可增强神经元细胞的酸毒性。敲除TMEM206基因不影响小鼠的生殖和发育,且能改善酸诱导的神经元细胞死亡,降低缺血性中风时脑梗死面积,抑制缺血性中风时脑损伤程度[11]。深圳华大基因研究院研究[12]发现,TMEM206可保护低氧环境导致脑损伤。因此,PAC通道在神经元中具有重要功能。
本研究中,敲除TMEM206基因的纯合小鼠均能正常生长发育,且成年鼠体质量与野生小鼠体质量无显著性差异。这证实了TMEM206基因虽广泛表达,但并非管家基因,敲除它不影响小鼠的生殖和发育。通过敲除野生C57BL/6J鼠的TMEM206基因,本研究发现小脑组织中265个基因表达量上调、209个基因表达量下调。GO功能分析显示,差异表达基因在结构上主要是细胞外膜和MHC Ⅰ。TMEM206蛋白本身是一种膜蛋白,细胞膜缺乏TMEM206蛋白时,其他膜蛋白基因表达可能发生改变,以维持细胞膜的结构与功能[13]。MHC Ⅰ类基因在中枢神经系统中均有表达,通常定位于轴突和树突表面,其在神经系统除发挥经典的免疫学功能外,还具有非免疫学功能,如调控神经系统的发育和突触可塑性[14]。有研究[15]表明,经典MHC Ⅰ类分子的异常表达与神经退行性疾病如帕金森病、阿尔茨海默症及肌萎缩性脊髓侧索硬化症等发病有关。SCA1是ATXN1基因突变引起的一种神经退行性疾病,ataxin-1与TMEM206蛋白存在相互作用,敲除TMEM206基因可引起MHC Ⅰ类分子表达异常,我们推测SCA1发病时,可能存在MHC Ⅰ类分子表达异常。
细胞黏附分子是一类介导细胞间或细胞与细胞外基质间相互接触和结合的分子。细胞黏附分子参与细胞的识别、活化、信号转导、增殖、分化,是免疫应答、炎症、凝血、肿瘤转移等重要生理和病理过程的分子基础[16]。对敲除小鼠TMEM206基因后小脑组织表达量发生改变的474个基因经KEGG分析,发现差异表达基因在功能上多为细胞黏附分子和肿瘤坏死因子路径分子,主要与细胞吞噬和衰老有关。TMEM206蛋白是一种离子通道蛋白,敲除TMEM206基因引起细胞黏附分子类的一些基因表达量发生改变,提示TMEM206蛋白功能或许与细胞识别、活化、增殖、分化及信号转导等有关。
本研究还发现,敲除TMEM206基因会引起一些与人乳头瘤病毒、Ⅰ型人类T淋巴细胞白血病病毒、单纯疱疹病毒、ⅤⅢ型人疱疹病毒及EB病毒等感染有关的基因出现表达量改变。与病毒感染的细胞结构一般为细胞表面受体,本研究显示敲除TMEM206基因,可导致一些细胞受体基因表达异常。
综上所述,通过敲除C57BL/6J鼠的TMEM206基因,本研究发现小脑组织有474个基因表达量发生改变,其中265个基因上调、209个基因下调,这些差异表达基因参与维持细胞外膜结构稳定和细胞黏附,并与病毒感染相关,提示作为离子通道蛋白,TMEM206蛋白在体内可能有众多的功能,其参与SCA1发病的机制有待进一步研究。
