西北地区草地贪夜蛾种群遗传多样性分析及治理策略
2024-03-28张大为陈靖魏玉红惠娜娜郭致杰罗进仓
张大为 陈靖 魏玉红 惠娜娜 郭致杰 罗进仓

摘 要 旨在明确甘肃省草地贪夜蛾的入侵来源,并制定科学有效的防控对策。基于mtCOI基因分子标记分析中国草地贪夜蛾不同生态区8个省12个地理种群276个样品的遗传多样性指数、遗传分化系数及基因流等。结果表明,甘肃省草地贪夜蛾种群的单倍型多样性指数和平均核苷酸差异数分别为0.133~0.157与0.133~0.317,均低于中国周年繁殖区广东、广西、云南种群的0.157~0.819与1.033~7.705;所有种群的Tajimas D中性检验和Fus F检验结果均为负值,表明草地贪夜蛾入侵中国后经历了明显的种群扩张事件。四川种群与其他种群遗传分化显著,62个种群间存在中等程度以上的基因交流。陕西略阳、陕西宁强、甘肃徽县、甘肃成县种群的有效迁入个体数和有效迁出个体数之和分别为11 860.66、11 708.65、10 878.66和10 379.32,在中国草地贪夜蛾的基因交流过程中具有中继站的作用,表明陕南汉水谷地为中国草地贪夜蛾西线北迁入侵西北的主要通道。
关键词 mtCOI基因;草地贪夜蛾;遗传多样性;遗传分化;基因流
草地贪夜蛾[Spodoptera frugiperda(J.E.Smith)]是原产于美洲热带和亚热带地区的重大农业迁飞性害虫[1-2]。在美洲适生寄主多达350余种,但主要嗜食玉米、高粱、甘蔗等C4作物[3-4],可导致玉米减产70%以上[5]。强大的适应能力及飞行能力使其在侵入非洲后短期内席卷撒哈拉以南的44个国家[6],2018年侵入印度后联合国粮农组织对草地贪夜蛾作出了全球预警,但其仍快速扩散至整个亚洲及大洋洲[7]。2019年草地贪夜蛾经缅甸入侵中国后迅速在26个省(区)发生危害[8-9],并在南方地区完成了定殖。鉴于其为害严重性,农业农村部将其列为中国“一类农作物病虫害”之首[10],草地贪夜蛾北迁日期的提前、虫源基数的逐年增加将对中国粮食安全构成持续的严峻威胁[11-13]。
甘肃地域狭长,是连接中国西北和西南地区的战略要道。陇南地区地处青藏高原东麓,横断山脉和秦岭的交汇处,是草地贪夜蛾随夏季东南季风进入中国西北地区桥头堡和重要的“中转补给站”[13]。甘肃省区域特色作物玉米、小麦、杂粮、马铃薯[14]等均是草地贪夜蛾的嗜食寄主,其中,全省玉米种植面积连年稳定在100万hm2以上[15],河西走廊制种玉米产量占全国一半以上。研究显示入侵中国的草地贪夜蛾以“玉米型”为主[16],因此,遏制草地贪夜蛾经陇南地区进一步向西北扩散对于保障中国的粮食安全具有重要 意义。
根据草地贪夜蛾的发生规律及为害特点,中国划定了周年繁殖区,迁飞过渡区和重点防范区实施分区治理策略[17]。区域性和暴发性的为害特点使得化学防治成为控制草地贪夜蛾最主要的手段[18],不同区划的害虫治理策略存在差异,进而草地贪夜蛾对药剂的敏感程度也不尽相同。因此,明确入侵中国西北地区草地贪夜蛾的入侵来源及潜在的扩散路径对于该虫的科学治理具有重要的指导作用。线粒体DNA是分析昆虫种群遗传结构的常用分子标记之一,其细胞色素氧化酶I亚基(mtCOI)基因相对保守且多态性好,已被广泛用于多种外来入侵昆虫的物种鉴定[19-21]、种群遗传分化[22-25]、基因流[26-28]及系统发育[20,29]等方面的研究。国内有学者研究了草地贪夜蛾入侵初期中国周年繁殖区不同种群的遗传多样性[30-31],但定殖中国后西北地区的草地贪夜蛾种群遗传多样性研究未见报道。因此,本研究采集中国草地贪夜蛾周年繁殖区(海南省、广西省、广东省、云南省)、迁飞过渡区(四川省、陕西省、湖北省)及重点防范区(甘肃省)的12个地理种群共276个样本,基于mtCOI分子标记技术分析不同地理种群的遗传多样性指数、遗传分化系数及基因流,以期揭示草地贪夜蛾定殖中国后不同地理种群遗传结构变化,分析入侵西北地区草地贪夜蛾的可能虫源地及扩散路径,为制定科学有效的防控对策提供理论依据。
1 材料与方法
1.1 供试虫源
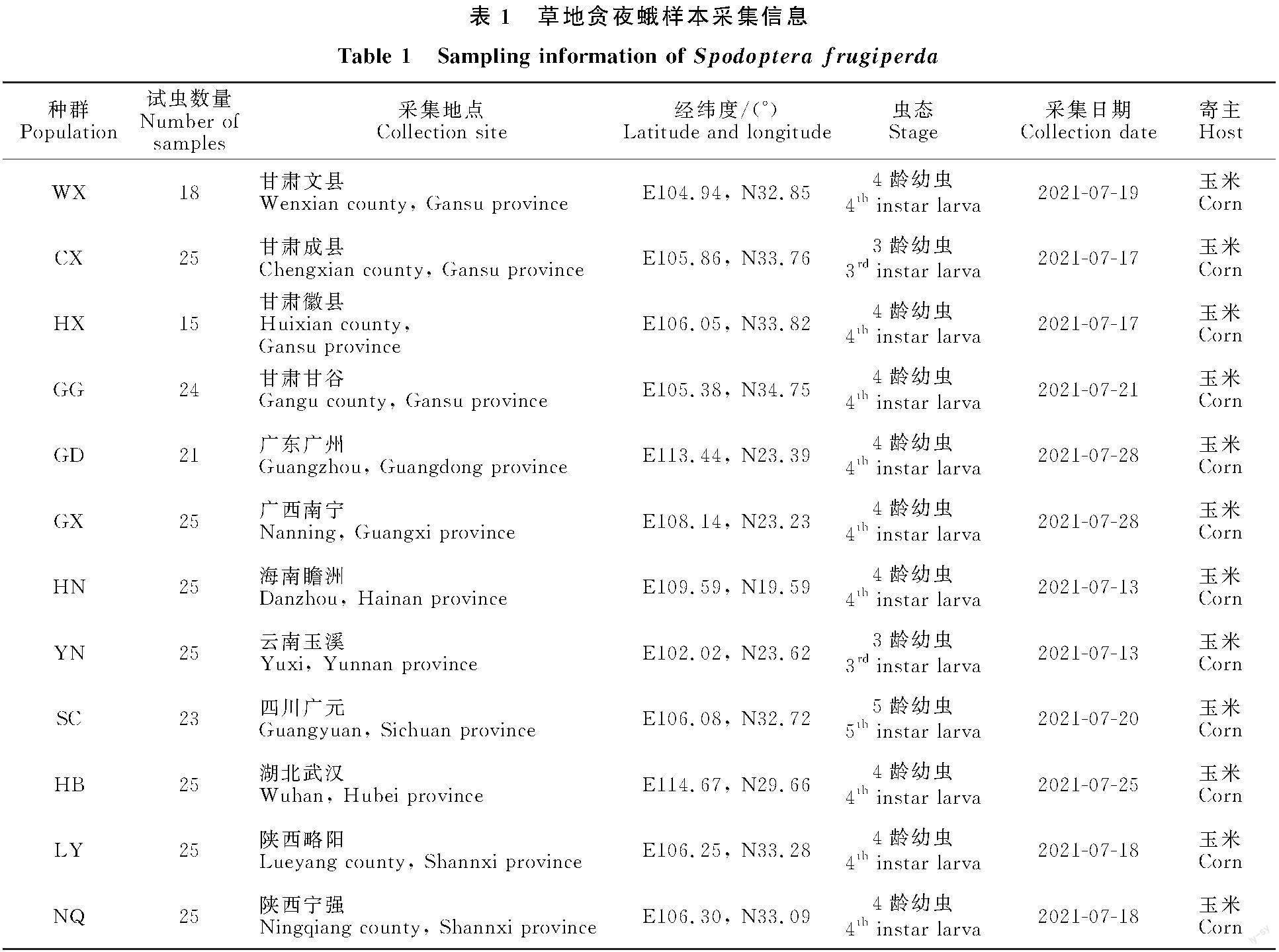
2021年5-7月在中国草地贪夜蛾周年繁殖区海南省、广东省、广西省、云南省,迁飞过渡区湖北省、陕西省、四川省和重点防范区甘肃省采集12个草地贪夜蛾种群共276个样本,具体采集点位信息见表1。各采样点选取草地贪夜蛾发生为害的典型地块,采用对角线五点取样法在玉米喇叭口期采集3~5龄幼虫,所有幼虫样本浸泡于95%的无水酒精,带回实验室后-20 ℃保存,备用。[FL)]
1.2 仪器和试剂
移液器,德国Eppendorf 公司;PCR仪,杭州博日科技有限公司,型号为XPcycle;水平电泳槽,北京君意东方电泳设备有限公司,JY-SPCT;台式高速冷冻离心机,美国BECKMAN公司,Allegra 21R;全自动数码凝胶图像分析系统,上海天能科技,Tanon-1600;電子天平,美国 AHOMS 公司,AR5120;DNA 提取试剂盒,天根生化科技(北京)有限公司,DP304;2×Taq PCR MasterMix,南京诺唯赞生物技术有限公司;AL2000 DNA Marker(2 000 bp),南京钟鼎生物技术有限公司,DM0101;离心管,美国Axygen公司,311-01-051;灭菌枪头,美国Axygen公司;其他试剂均为国产分析纯。
1.3 草地贪夜蛾基因组DNA的提取
参照DNA提取试剂盒说明书提取单头草地贪夜蛾幼虫样本的基因组DNA,提取后的DNA以20 μL的超纯水稀释至200 ng/μL,-20 ℃保存,备用。
1.4 草地贪夜蛾mtCOI基因序列的扩增
根据NCBI已公布的草地贪夜蛾线粒体DNA全长序列(GenBank登录号:KM362176.1),参考李向永等[31]设计的草地贪夜蛾mtCOI特异性引物,选取FAW-F:5′-CAACATTTATTTTGATTTTTTGG-3′和FAW-R:5′-CCATTACATATAATCTGCCATATTA-3′用于基因扩增,引物由南京集思慧远生物科技有限公司合成。
PCR反应体系(20 μL):2×Taq PCR MasterMix,10 μL;正向引物(10 μmol/L),1 μL;反向引物(10 μmol/L),1 μL;模板DNA,1 μL;ddH2O,7 μL。PCR反应程序如下:95 ℃预变性5 min;95 ℃变性30 s,57 ℃退火30 s,72 ℃延伸1 min,循环32次;最后72 ℃再延伸5 min。扩增结束后取20 μL扩增产物在1%琼脂糖凝胶电泳分离30 min,经凝胶成像系统分析后拍照保存。
1.5 数据分析
所有样品的PCR产物回收纯化后,委托南京集思慧远生物科技有限公司进行双向测序。使用Mafft软件(V7.427)对一代测序序列进行质量检测及多序列比对;采用DnaSP 6.0(V6)软件计算草地贪夜蛾12个地理种群的多态位点数、单倍型多样性、核苷酸多样性、Tajimas D中性检验和Fus F检验等;采用Arlequin(V3.5.2.2)計算12个草地贪夜蛾种群的遗传分化系数Fst,并根据Nm=(1-Fst)/2Fst公式计算种群间的基因流Nm;采用Migrate(V3.7.2)软件计算12个草地贪夜蛾种群的有效迁入数及迁出数。
2 结果与分析
2.1 草地贪夜蛾不同地理种群的遗传多样性 指数
完成了对草地贪夜蛾12个地理种群276个样本的DNA提取,所有样本DNA带型一致,纯度好且无降解现象发生,符合mtCOI分子标记测序的要求。经Mafft软件对DNA序列进行多序列比对后,供试的276个草地贪夜蛾样品共获得243个mtCOI基因序列,所有mtCOI基因序列均无插入或缺失,根据比对结果选取782 bp长度进行后续分析。
在12个草地贪夜蛾地理种群中,变异的多态性位点为27,单倍型多样性为0.273,核苷酸多样性为0.436,平均核苷酸差异数为3.184,种群间Tajimas D值为-2.567 3,(P< 0.001),Fus F值为-8.262 3,(P<0.01),差异极显著,表明草地贪夜蛾定殖中国后发生了全面、快速的种群扩张。周年繁殖区GD、GX和YN种群的单倍型多样性为0.157~0.819,核苷酸多样性为 0.137~1.041,平均核苷酸差异数为1.033~ 7.705;迁飞过渡区SC、HB、LY和NQ种群的单倍型多样性为0.080~0.672,核苷酸多样性为 0.021~ 1.646,平均核苷酸差异数为0.160~ 12.336;甘肃省CX和HX种群的单倍型多样性为0.133~ 0.157,核苷酸多样性为0.017~ 0.041,平均核苷酸差异数为0.133~0.317。GD种群的单倍型多样性和核酸多样性最高,最低为甘肃的HX种群(表2)。
中性检验显示,中国周年繁殖区GD、GX和YN种群的Tajimas D值为-2.293 2~ -0.170 8,Fus F值为-3.580 8~-0.305 5;迁飞过渡区SC、HB、LY和NQ种群的Tajimas D值为-2.574 1~-1.183 5,Fus F值为 -4.081 8~-2.300 3;甘肃CX和HX种群的Tajimas D值为 -1.685 0~-1.303 2,Fus F值在-2.313 4~-1.262 1。所有种群的Tajimas D和Fus F值均为负值,HB、NQ和YN种群表现极显著,甘肃的CX种群也表现显著,推测这4个种群近期经历了明显的种群扩张。
2.2 草地贪夜蛾不同地理种群的遗传分化
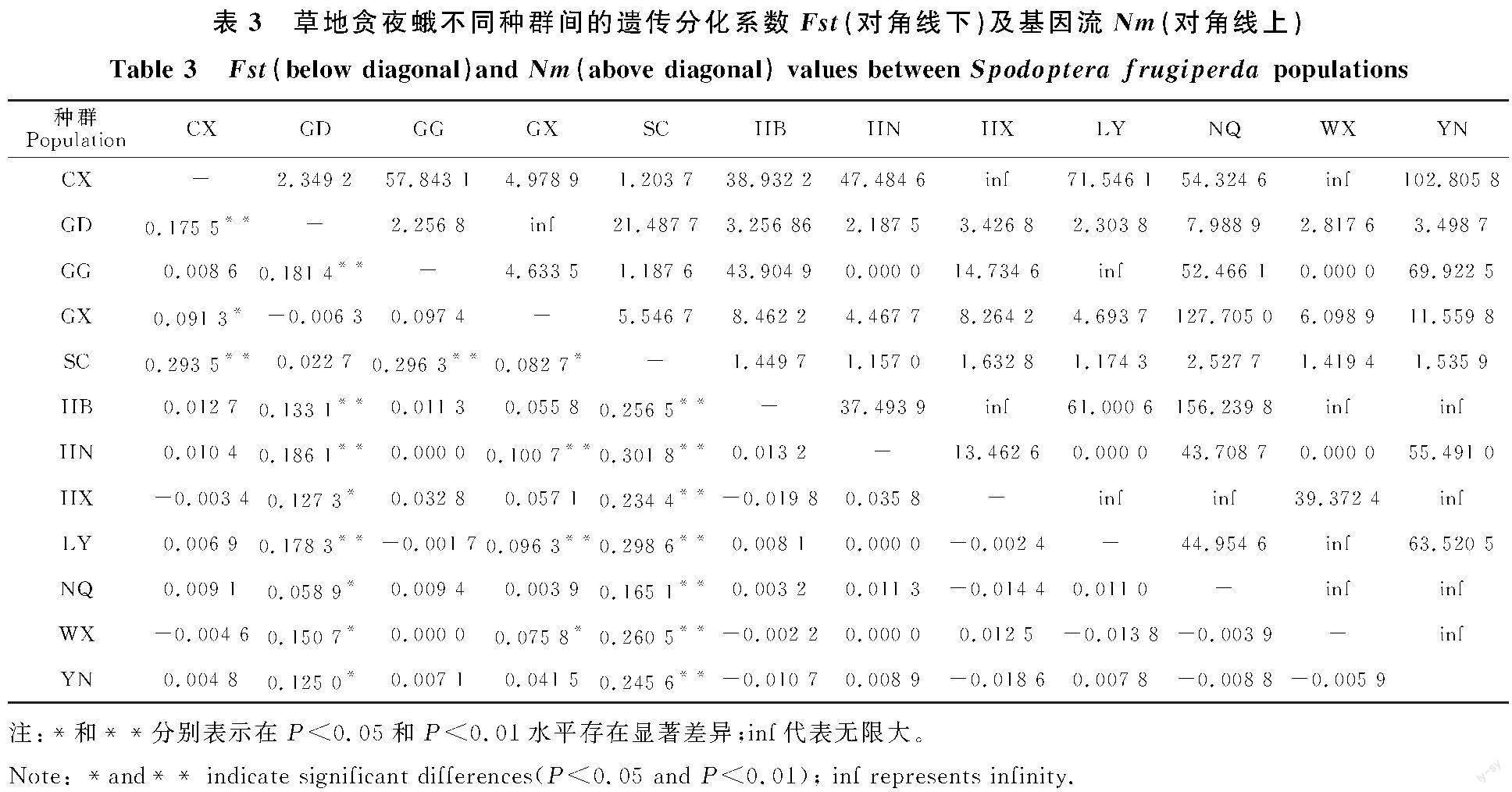
通过Arlequin软件计算草地贪夜蛾种群内的遗传分化系数Fst,并根据Roussed提出的标准分析不同种群由遗传结构引起的种群分化情况[32]。将12个种群两两组合分为66对(表3),有16对的Fst值极显著高于其他43对(P< 0.001),有7对的Fst值显著高于其他43对(P< 0.05)。在所有的12个地理种群中,SC分化尤为显著,与HN、YN、GX、NQ、LY以及甘肃的HX、WX、CX和GG种群之间出现高度或极度分化,Fst分别为0.301 8、0.245 6、0.082 7、0.165 1、 0.298 6、0.234 4、0.260 5、0.293 5和0.296 3,与广东种群的遗传分化较低(Fst值为0.022 7)。
2.3 不同草地贪夜蛾地理种群间的基因流
66对草地贪夜蛾不同地理种群间的基因流(Nm)为0~无限大(inf),其中,45对草地贪夜蛾种群间的Nm大于4,表明这些种群间的基因交流普遍发生;有17个种群间的基因流Nm为1~4,表明这些种群间有中等程度的基因交流;4个种群间的基因交流有限(表3)。甘肃省CX、HX、WX、GG种群与周年繁殖区GX和YN种群间基因流均大于4,推测入侵甘肃的草地贪夜蛾可能主要来源于云南、广西草地贪夜蛾的北迁西线。
Migrate软件分析显示,中国周年繁殖区GD种群、HN种群、GX种群和YN种群每世代的有效迁出数分别为4 195.66、5 650.99、4 565.66和3 565.67,每世代的有效迁入数分别为2 139.66、3 943.65、3 142.33和4 539.65。HN种群的有效迁入和迁出个体数之和较高,为中国草地贪夜蛾周年繁殖区的主要基因交流中继站,YN种群的有效迁入个体数较高,推测为草地贪夜蛾由缅甸经云南持续入侵中国的原因。迁飞过渡区HB种群、SC种群、NQ种群和LY种群每世代的有效迁出数为4 299.65、4 876.34、4 896.33和 5 137.66,每世代的有效迁入数为3 616.99、 2 755.66、6 812.32和6 723.00,陕西NQ种群和LY种群的有效迁入和迁出个体数和较高,推测陕南的汉水谷地为中国草地贪夜蛾由南向北入侵的重要中转基地。甘肃CX种群、GG种群、HX种群、WX种群每世代的有效迁出数为3 644.99,4 893.65、4 794.99和5 080.99,每世代的有效迁入数为6 734.33、4 334.99、6 083.67和4 776.33,其中CX种群和HX种群的有效迁入和迁出个体数总和较高,在甘肃省草地贪夜蛾进一步的北迁扩散中起着重要的作用(表4)。
3 结论与讨论
草地贪夜蛾2019年已经在中国南方地区完成定殖过程,并随季风往返于大部分省(区)发生为害,持续威胁中国的粮食生产安全[12]。2022年全国草地贪夜蛾的发生程度显著高于往年,且发生北界北移3个纬度以上[33],表明完成定殖后该虫入侵势头的加剧。分子标记技术可以深入的分析草地贪夜蛾的种群遗传分化及基因流,进而明确中国西北地区草地贪夜蛾的入侵来源及亲缘关系,为该虫的科学治理提供理论依据。本研究基于COI基因鉴定到12个草地贪夜蛾种群有多种单倍型,与陈冬平等的研究结果类似[34],与草地贪夜蛾入侵初期云南种群只有1种单倍型的研究不一致[16,31],原因可能与本研究选择的COI基因母系遗传特征有关,基于核基因组的Tpi基因能更准确地鉴定生物型,再者草地贪夜蛾在中国的持续入侵扩散进程中可能发生了遗传杂交现象。本研究中12个草地贪夜蛾种群的单倍型多样性指数为0.273,云南种群为0.230,显著低于其他周年繁殖区种群,李向永等[31]的研究也发现云南种群的单倍型多样性指数较低(0.047~0.214),可能由于云南为草地贪夜蛾经缅甸入侵中国的第一站,种群来源单一,种群遗传多样性不高,而入侵广东、广西的草地贪夜蛾种群数量多,来源广,并已经在当地形成稳定种群。
Tajimas D中性检验和Fus F统计检验对种群扩张很敏感,在种群发生扩张时通常显示为负值。本研究全部12个种群的Tajimas D中性检验和Fus F统计值均为负值,表明草地贪夜蛾自2019年侵入中国后经历了明显的种群扩张,且呈现持续的扩张态势。湖北种群、宁强种群和云南种群的表现尤为显著,原因可能是云南为草地贪夜蛾经缅甸入侵中国的第一站,湖北和陕西为草地贪夜蛾随季风在中国由南向北迁移过程的主要中转站。中性检验的结果也验证了吴秋琳等[13]对草地贪夜蛾在中国西北地区的迁飞路线的预测。
12个种群的遗传分化结构显示,四川种群与广东种群遗传分化较低,与其他种群出现高度或极度分化,推测四川种群最初来源于广东,遗传分化可能与当地独特的地理环境及农户粗放的管理习惯有关。不同地理种群间的基因流Nm显示,中国的绝大部分草地贪夜蛾种群存在中等程度以上的基因交流。此外,12个种群的有效迁移个数分析显示,周年繁殖区的海南种群、陕西宁强种群、略阳种群、甘肃成县种群和徽县种群的个体迁移总和较高,推测以上种群为中国草地贪夜蛾的基因交流中继站。基因流分析表明,陕南汉水谷地区域可能为中国草地贪夜蛾北迁的交汇中心,也是甘肃省草地贪夜蛾的主要入侵来源。甘肃陇南成县和徽县种群在草地贪夜蛾进一步向陇中、宁夏及内蒙的传播扩散中起着重要的作用。因此,本研究遗传分化结果也验证了吴秋琳等[13]对草地贪夜蛾在中国西北地区迁飞扩散路径的 预测。
本研究对草地贪夜蛾定殖中国后不同生态区划12个地理种群的遗传多样性进行了解析,初步验证了云南为中国草地贪夜蛾的最初入侵地,陕南的汉水河谷地区为草地贪夜蛾随东南季风北迁的主要通道,甘肅陇南地形复杂且寄主类型丰富,是草地贪夜蛾进一步入侵中国西北的重要“中转站”。但草地贪夜蛾在中国定殖时间尚短,本研究的采样点偏少,覆盖面不广,今后应继续加大采样密度和频次,更大尺度阐明中国草地贪夜蛾种群的遗传结构变化及迁飞扩散路径,为该虫的综合治理提供更为全面的科学数据。
作为典型的长距离迁飞性害虫,草地贪夜蛾定殖后将随季风“北迁南回”在中国各大粮食主产区发生为害。西北地区是中国重要的玉米、小麦、马铃薯及制种玉米生产基地,有效削减草地贪夜蛾的北迁基数对于中国粮食安全具有举足轻重的作用。本研究表明汉水河谷地区是中国草地贪夜蛾夏季北迁西线的重要中转地,鉴于此,今后西北地区草地贪夜蛾防控阻截的重点是在汉水河谷地区的陕南、甘肃陇南等地利用高空探照灯及性诱捕器等手段,加强对草地贪夜蛾迁飞成虫的动态监控,并在复种玉米等重点作物持续开展幼虫的田间监测,研发物理诱控与化学防治相结合的高效防治技术措施,做到早发现早消灭,最大限度压低进一步北迁的虫源基数。此外,基于中国日趋成熟的昆虫雷达应用技术,建议在周年繁殖区云南、海南等地的草地贪夜蛾迁飞路线布设监测雷达,第一时间掌握成虫的迁飞行为及迁飞方向。同时,在草地贪夜蛾周年繁殖区及迁飞过渡区持续开展田间种群的动态监测及抗药性监测研究,并深入开展生物药剂研发、生物天敌筛选、Bt玉米种植等生物、生态治理技术研究。在北方重点防范区集成研发草地贪夜蛾监测预警与应急防控技术,构建更为科学完善的全国草地贪夜蛾跨区联动防控体系,确保草地贪夜蛾定殖后国内粮食安全及生态安全。
参考文献 Reference:
[1] 江幸福,张 蕾,程云霞,等.草地贪夜蛾迁飞行为与监测技术研究进展[J].植物保护,2019,45(1):12-18.
JIANG X F,ZHANG L,CHENG Y X,et al. Advances in migration and monitoring techniques of the fall armyworm,Spodoptera frugiperda (J.E.Smith)[J].Plant Protection,2019,45(1):12-18.
[2] 郭井菲,何康来,王振营.草地贪夜蛾的生物学特性、发展趋势及防控对策[J].应用昆虫学报,2019,56(3):361-369.
GUO J F,HE K L,WANG ZH Y.Biological characteristics,trend of fall armyworm Spodoptera frugiperda,and the strategy for management of the pest[J].Chinese Journal of Applied Entomology,2019,56(3):361-369.
[3] MONTEZANO D G,SPECHT A,SOSA-GMEZ D R, et al. Host plants of Spodoptera frugiperda (Lepidoptera:Noctuidae) in the Americas[J].African Entomology,2018,26(2):286-300.
[4] HARDKE J T,LORENZ III G M,LEONARD B R.Fall armyworm (Lepidoptera:Noctuidae) ecology in southeastern cotton[J].Journal of Integrated Pest Management,2015,6(1):DOI:10.1093/jipm/pmv009.
[5] MURA G,MOLINA-OCHOA J,COVIELLA C.Population dynamics of the fall armyworm,Spodoptera frugiperda (Lepidoptera:Noctuidae) and its parasitoids in northwestern Argentina[J].Florida Entomologist,2006,89(2):175-183.
[6] EARLY R,GONZALEZ-MORENO P,MURPHY S T, et al.Forecasting the global extent of invasion of the cereal pest Spodopterafrugiperda,the fall armyworm[J].NeoBiota,2018,40:25-50.
[7] JING D P,GUO J F,JIANG Y Y,et al.Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques[J].Insect Science, 2020,7(4):780-790.
[8] 刘 杰,姜玉英,吴秋琳,等.我国草地贪夜蛾冬春季发生为害特点及下半年发生趋势分析[J].中国植保导刊,2019,39(7):36-38,49.
LIU J,JIANG Y Y,WU Q L,et al. Occurrence and damage characteristics of Spodoptera frugiperda in winter to spring and its occurrence trend in the second half year in China[J].China Plant Protection,2019,39(7):36-38,49.
[9] 姜玉英,劉 杰,谢茂昌,等.2019年我国草地贪夜蛾扩散为害规律观测[J].植物保护,2019,45(6):10-19.
JIANG Y Y,LIU J,XIE M CH,et al. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019[J].Plant Protection,2019,45(6):10-19.
[10] 中华人民共和国农业农村部.中华人民共和国农业农村部公告第333号:一类农作物病虫害名录[EB/OL].[2020-03-15].http:∥www.moa.gov.cn/govpublic/ZZYGLS/202009/t20200917_6352227.htm.
Ministry of Agriculture and Rural Affairs of the Peoples Republic of China.Announcement No.333 of the Ministry of Agriculture and Rural Affairs of the Peoples Republic of China:List of category I of crop diseases and pests [EB/OL].[2022-03-15].http:∥www.moa.gov.cn/govpublic/ZZYGLS/202009/t20200917_6352227.htm.
[11] 姜玉英,刘 杰,吴秋琳,等.我国草地贪夜蛾冬繁区和越冬区调查[J].植物保护,2021,47(1):212-217。
JIANG Y Y,LIU J,WU Q L,et al. Investigation on winter breeding and overwintering areas of Spodoptera frugiperda in China[J].Plant Protection,2021,47(1):212-217.
[12] 吴孔明.中国草地贪夜蛾的防控策略[J].植物保护,2020,46(2):1-5.
WU K M.Management strategies of fall armyworm (Spodoptera frugiperda) in China[J].Plant Protection,2020,46(2):1-5.
[13] 吴秋琳,姜玉英,刘 媛,等.草地贪夜蛾在中国西北地区的迁飞路径[J].中国农业科学,2022,55(10):1949-1960.
WU Q L,JIANG Y Y,LIU Y,et al.Migration pathway of Spodoptera frugiperda in Northwestern China[J].Scientia Agriculture Sinica,2022,55(10):1949-1960.
[14] 龚大鑫.甘肃省区域特色农业竞争力研究[D].兰州:甘肃农业大学,2012.
GONG D X.A study on the competitiveness of regional specialized agriculture in Gansu province[D].Lanzhou:Gansu Agricultural University,2012.
[15] 甘肃省统计局.2021甘肃发展年鉴[M].北京.中国统计出版社,2021.
Gansu Provincial Bureau of Statistics.Gansu Development Yearbook of 2021[M].Beijing.China Statistics Press,2021.
[16] 张 磊,靳明辉,张丹丹,等.入侵云南草地贪夜蛾的分子鉴定[J].植物保护,2019,45(2):19-24
ZHANG L,JIN M H,ZHANG D D,et al.Molecular identification of invasive fall armyworm Spodoptera frugiperda in Yunnan province[J].Plant Protection,2019,45(2):19-24.
[17] 中华人民共和国农业农村部.农业农村部关于印发《2020年全国草地贪夜蛾防控预案》的通知[Z].北京:中华人民共和国农业农村部,2020:39-43.
Ministry of Agriculture and Rural Affairs of the PeopleS Republic of China.Notice of the Ministry of Agriculture and Rural Affairs on printing and distributing the national plan for the prevention and control of the fall armyworm in 2020[Z].Beijing:the Ministry of Agriculture and Rural Affairs of the Peoples Republic of China,2020:39-43.
[18] 吳孔明,杨现明,赵胜园,等.草地贪夜蛾防控手册[M]. 北京:中国农业科学技术出版社,2020.
WU K M,YANG X M,ZHAO SH Y,et al.Handbook on the Prevention and Control of the Fall Armyworm Spodoptera frugiperda[M].Beijing:China Agricultural Science and Technology Press,2020.
[19] 周青松.DNA条形码在寄生蜂鉴定中(昆虫纲:膜翅目)的应用研究[D].合肥:安徽大学,2014.
ZHOU Q S.Application of DNA barcoding studies in Hymenopteran parasitoids(Insecta:Hymenoptera)[D].Hefei:Anhui University,2014.
[20] 许丽丽,蔡 力,沈伟江,等.中国部分地区烟粉虱生物型种类及系统发育关系分析[J].应用生态学报,2014,25(4):1137-1144.
XU L L,CAI L,SHEN W J,et al.Biotypes and phylogenetic analysis of Bemisia tabaci (Hemiptera:Aleyrodidae) in China[J].Chinese Journal of Applied Ecology,2014,25(4):1137-1144.
[21] 唐运林,顾偌铖,吴燕燕,等.入侵重庆地区的草地贪夜蛾种群生物型鉴定[J].西南大学学报(自然科学版),2019,41(7):1-7.
TANG Y L,GU R CH,WU Y Y,et al.Biotype identification of the population of Spodoptera frugiperda that migrated to Chongqing area[J].Journal of Southwest Univerrsity(Natural Science Edition),2019,41(7):1-7.
[22] 周宁宁,王梦馨,崔 林,等.基于COI基因全长序列的假眼小绿叶蝉地理种群遗传分化研究[J].生态学报,2014,34(23):6879-6889.
ZHOU N N,WANG M X,CUI L,et al.Genetic variation of Empoasca vitis (Gthe) (Hemiptera:Cicadellidae) among different geographical populations based on mtDNA COI complete sequence[J].Acta Ecologica Sinica,2014,34(23):6879-6889.
[23] 郑 燕.基于微卫星和线粒体标记的梨小食心虫种群遗传多样性和遗传结构研究[D].陕西杨凌:西北农林科技大学,2014.
ZHENG Y.Genetic diversity and genetic structure of oriental fruit moth,Grapholita molesta (L.) analyzed by microsatellite markers and DNA gene sequences[D].Yangling Shaanxi:Northwest A&F University,2014.
[24] 王 蒙,徐 浪,张润杰,等.基于线粒体COI基因的桔小实蝇种群遗传分化研究[J].昆虫学报,2014,57(12):1424-1438.
WANG M,XU L,ZHANG R J,et al.Population genetic differentiation analysis of the oriental fruit fly,Bactrocera dorsalis (Diptera:Tephritidae) based on mtDNA COI gene[J].Acta Entomologica Sinica,2014,57(12):1424-1438.
[25] 王佳琪.基于线粒体COI基因的玉米黄呆蓟马不同地理种群遗传多样性研究[D].长春:吉林大学,2021.
WANG J Q.Genetic diversity of different geographical populations of Anaphothrips obscurusbased on mitochondrial COI gene[D].Changchun:Jilin University,2021.
[26] 李 菁.基于分子标记和Wolbachia感染检测的亚洲玉米螟种群遗传分化与基因流研究[D].北京:中国农业科学院,2010.
LI J.Studies on genetic differentiation and gene flow of Ostrinia furnacalis populations based on multiple molecular markers and Wolbachia infection detection[D].Beijing:Chinese Academy of Agricultural Sciences,2010.
[27] 王兴亚,许国庆.中国甜菜夜蛾地理种群的遗传分化与基因流[J].昆虫学报,2014,57(9):1061-1074.
WANG X Y,XU G Q.Genetic differentiation and gene flow among geographic populations of Spodoptera exigua (Lepidoptera:Noctuidae) in China[J].Acta Entomologica Sinica,2014,57(9):1061-1074.
[28] 孙 嵬,张柱亭,董 辉,等.基于线粒体COI基因序列的黄胫小车蝗不同地理种群的遗传分化及基因流分析[J].昆虫学报,2013,56(8):907-916.
SUN W,ZHANG ZH T,DONG H,et al.Analysis of genetic differentiation and gene flow among different geographic populations of Oedaleus infernalis (Orthoptera:Acrididae) based on mtDNA COI gene sequences[J].Acta Entomologica Sinica,2013,56(8):907-916.
[29] 褚 棟,张友军,徐宝云,等.烟粉虱不同地理种群的mtDNA COI基因序列分析及其系统发育[J].中国农业科学,2005,38(1):76-85.
CHU D,ZHANG Y J,XU B Y,et al.Sequence analysis of mtDNA COI gene and molecular phylogeny of different geographical populations of Bemisia tabaci (Gennadius)[J].Scientia Agriculture Sinica,2005,28(1):76-85.
[30] 王佳丽,韦加奇,孙志秀,等.入侵我国中南三省(区)草地贪夜蛾的单倍体型和群体遗传结构分析[J].华南农业大学学报,2020,41(1):9-16.
WANG J L,WEI J Q,SUN ZH X,et al.Haplotype and genetic diversity analysis of Spodoptera frugiperda invading in three provinces of central and southern China[J].Journal of South China Agricultural University,2020, 41(1):9-16.
[31] 李向永,尹艳琼,KHIN T N,等.基于mtCOI基因序列的我国云南省和缅甸、柬埔寨草地贪夜蛾种群遗传多样性分析[J].植物保护学报,2020,47(4):789-796.
LI X Y,YIN Y Q,KHIN T N,et al.Analysis on population genetic diversity of fall armyworm Spodoptera frugiperda in Yunnan Province,China,Myanmar and Cambodia based on mtCOI gene sequence[J].Journal of Plant Protection,2020,47(4):789-796.
[32] ROUSSET F.Genetic differentiation and estimation of gene flow from f-statistics under isolation by distance[J].Genetics,1997,145(4):1219-1228.
[33] 中華人民共和国农业农村部.农业农村部部署草地贪夜蛾防控工作[EB/OL].(2022-07-29)[2022-10-31].http:∥www.moa.gov.cn/xw/zwdt/202207t20220729_6405815.htm.
Ministry of Agriculture and Rural Affairs of the Peoples Republic of China.The Ministry of Agriculture and Rural Affairs deployed the prevention and control of the fall armyworm [EB/OL].(2022-07-29)[2022-10-31].http:∥www.moa.gov.cn/xw/zwdt/202207/t20220729_6405815.htm.
[34] 陈冬平,陈志廷,徐汉虹,等.应用不同分子标记鉴别入侵广州草地贪夜蛾的寄主型[J].华南农业大学学报,2020,41(1):17-21.
CHEN D P,CHEN ZH T,XU H H,et al.Host type identification of Spodoptera frugiperda invading in Guangzhou by different molecular markers[J].Journal of South China Agricultural University,2020,41(1):17-21.
Genetic Diversity Analysis and Governance Strategy of Spodoptera frugiperda Populations in Northwest China
Abstract This study aims to elucidate the invasion source and develop scientifically sound and effective prevention and control measures for Spodoptera frugiperda in Gansu province.Using molecular markers based on mtCOI gene,we analyzed the genetic diversity index,genetic differentiation coefficient and gene flow of S.frugiperda across 12 geographical populations in 8 provinces of China.The results showed that the haplotype diversity and average nucleotide difference in Gansu populations ranged from 0.133 to 0.157 and 0.133 to 0.317,respectively.These values were lower than thoose observed in Guangdong,Guangxi,and Yunnan populations within the annual breeding areas of China,which ranged from 0.157 to 0.819 and 1.033 to 7.705 respectively.Both Tajimas D neutral test and Fus F test yielded negative values across all populations,suggesting a significant population expansion event for S.frugiperda following its invasion of China.There was genetic differentiation between Sichuan population and others,while gene flow among 62 populations were found to be at an above-medium level.The cumulative effective emigration and immigration values for the Lueyang,Ningqiang,Huixian county and Chengxian county populations were 11 860.66,11 708.65,10 878.66 and 10 379.32,respectively.These play the role as relaying station in the process of gene exchange of S.frugiperda in China.The Hanshui Valley is an important corridor for the westward migration of S.frugiperda into the northwest in China.
Key words
Spodoptera frugiperda; mtCOI gene; Population genetic diversity; Genetic differentiation; Gene flow
