亚热带4种林分土壤真菌群落结构及功能
2024-03-18文翕,党鹏
文 翕, 党 鹏
(1.中南林业科技大学林学院;2.中亚热带林学国家长期科研基地,湖南 长沙 410004)
亚热带地区的森林是全球森林的重要组成部分,对陆地生态系统的生物多样性、碳储量、生态系统稳定性以及木材生产具有重要影响[1]。中国亚热带地区是我国森林多样性最丰富的地区之一,主要由常绿阔叶林组成。此外,杉木(Cunninghamialanceolata)、马尾松(Pinusmassoniana)等速生针叶树种也广泛栽植于该地区。然而,杉木和马尾松作为我国南方种植面积较大的两种人工林,虽然缓解了木材需求压力,但也带来了一系列生态问题,如生物多样性下降和生产力下降。纯针叶林树种种植单一,导致凋落物分解缓慢,降低了养分循环速率,从而引起土壤退化[2]。次生针阔混交林是亚热带地区的重要地带性森林类型,其多样化的树种组成有利于改善土壤肥力和提高森林生态系统的稳定性[3]。土壤微生物是土壤中数量和种类较多的生物群落之一,其参与森林生态系统生物化学循环,具有重要的生态功能[4],与土壤理化特性相比,土壤微生物特别是真菌能更好地反映土壤质量[5-6]。真菌作为土壤微生物的重要组成部分,具有分解有机物[7]、促进土壤稳定和改善土壤结构的作用[8];同时,一些真菌病原体也能通过侵染植物组织来抑制宿主植物的生长和繁殖[9]。因此,研究真菌群落的结构、多样性和功能特性有助于评估森林土壤的健康状况和质量。
真菌群落的结构和多样性受到环境因素和植物的影响。环境因素可以通过改变土壤性质来影响真菌群落,例如土壤温度、水分、酸碱性和养分含量等[10]。不同树种的差异也会对土壤真菌群落产生相应的影响。不同的营林模式会影响土壤特性,从而对微生物的结构和多样性产生影响。例如,与青杄林相比,华北落叶松林的土壤真菌多样性更高[8]。在不同森林类型中,优势真菌门也存在差异,如蒙古栎林的土壤真菌群落以子囊菌门和担子菌门为主[11],而神农架山林的土壤中优势菌类属于接合菌门[12]。因此,植被类型对真菌群落的影响方式存在一定的不确定性。
土壤中的真菌群落具有不同的功能群。共生真菌(如外生菌根真菌和丛枝菌根真菌)与植物密切关联,形成互惠共生关系,以改善养分交换。腐生真菌是凋落物、木质碎片和死根的主要分解者,通过分解这些有机物获得能量,并促进其生长[13]。近年来,学者们对亚热带森林土壤中微生物的研究逐渐增多,主要集中在土壤理化性质[14]、土壤细菌功能群[15]以及土壤微生物量[16]的分析上,而对于亚热带典型林分的土壤真菌群落结构和功能特征差异的报道较少。
本研究以亚热带典型森林类型杉木人工林、马尾松人工林、甜槠林及针阔混交林为对象,采集林下土壤样品,利用高通量测序技术,通过Pearson相关性分析和冗余分析(redundancy analysis, RDA)比较土壤养分、土壤真菌多样性和群落组成、结构之间的差异。研究旨在探究土壤真菌不同功能群在这4种林分中的分布格局,并揭示影响土壤真菌群落结构和功能特征的主要驱动因素,为亚热带森林的培育和可持续经营提供依据。
1 材料与方法
1.1 研究区概况
野外样地布设于湖南省岳阳市平江县芦头试验林场(28°31′17″—28°38′00″N,113°51′52″—113°58′24″E),海拔124.5~1 272.5 m。年平均降水量约1 450 mm,年平均温度18.5 ℃,研究地点具有典型的湿润中亚热带季风气候特征,年无霜期260 d,年日照时数为1 930 h。地貌类型主要是中低山地形,山势陡峭,山脚土层极薄,部分地区有岩石裸露,而山中上部土层较为厚实,平均坡度为25°,海拔600 m以下主要为红壤地区。
芦头试验林场的林地经营总面积4 940 hm2,森林覆盖率92%,森林总蓄积量43.31×104m3。该地区的动植物资源丰富(1 600多种)。地带性植被类型为中亚热带常绿阔叶林。在林场内,森林植被类型多样,包括甜槠林、苦槠林、钩栲林、青冈林和石栎林等。这些森林都属于典型的常绿阔叶林,拥有丰富的生物多样性。本试验选择立地条件相近的杉木人工纯林、马尾松人工纯林、甜槠次生林和针阔混交次生林(主要树种为杉木、马尾松、甜槠和青冈等)进行土壤采集和植被调查(表1)。
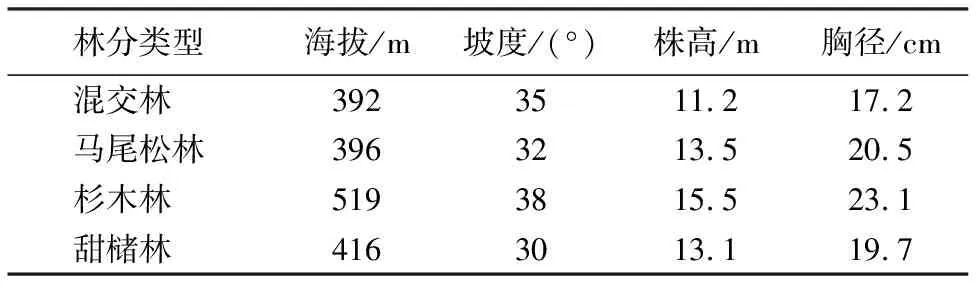
表1 样地概况Table 1 Conditions of sampling plots
1.2 样品采集与预处理
于2021年10月,在研究区选取马尾松林、杉木林、甜槠纯林及其混交林作为研究对象。每种林分设置4个20 m×20 m的样地,样地距离林缘至少10 m,样方间隔10 m以上。在每个样地中进行每木检尺以及灌草层植物的调查,包括植物种类、数量、覆盖度、株高、胸径和冠幅等参数。在每个样地中,按照S型取样方法,使用土钻采集5个0~20 cm的土壤样品,并将其充分混匀后作为混合土壤样品,每种林分4个混合样品,共计16个。迅速将采集完的土壤样品放置在冷藏保温箱中运回实验室。将土壤样品过2 mm筛后分成两部分:一部分置于阴凉干燥处自然风干,用于测定土壤理化指标;另一部分于-80 ℃冷冻保存,用于土壤微生物测定。
1.3 土壤理化性质测定
1.4 土壤真菌DNA提取、扩增和测序
采用MagPure Soil DNA LQ Kit(MagPure®Soil DNA LQ Kit, D 6356-03)试剂盒,从0.5 g土壤中提取土壤核酸,并进行0.8%琼脂糖凝胶电泳;采用利用紫外定量设备(NC2000, Thermo Scientific)对DNA进行定量。利用特异性引物ITS5(5′-GGAAGTAAAAGTCGTAACAAGG-3′) ITS2(5′-GCTGCGTTCTTCATCGATGC-3′)对真菌ITS进行扩增。PCR扩增体系共25 μL,包含:DNA模板2 μL,上下游引物各1 μL(10 μmol·L-1),缓冲液5 μL,Q5高保真缓冲液0.25 μL,保真DNA聚合酶5 μL,dNTP(2.5 mmol·L-1) 2 μL,超纯水(ddH2O) 8.75 μL。PCR扩增条件为:预变性(98 ℃,30 s)、变性(98 ℃,15 s)、退火(50 ℃,30 s)、延伸(72 ℃,30 s),共25个循环,最后在72 ℃延伸5 min,4 ℃保存。对扩增结果进行2%琼脂糖凝胶电泳,切取目的片段并回收。利用Quant-iT PicoGreen dsDNA Assay Kit在Microplate reader (FLx800, BioTek)上对PCR产物进行定量,并混样。由上海派森诺生物科技有限公司使用NovaSeq 6000进行2×250 bp的双端测序。
1.5 数据分析
使用Qiime 2切除序列的引物片段,调用DADA 2进行质控、去噪、拼接和去除嵌合体。去除singletons ASVs,合并ASVs的特征序列和ASV表格。最后,通过UNITE数据库对获得的代表性序列进行物种注释。
使用Qiime平台计算群落的α多样性,并使用Excel 2019计算物种的相对丰度。然后,在微科盟生物云和联川生物平台上作图。使用R语言基于Bray-Curtis矩阵计算群落的β多样性,并利用非量度多维尺度(non-metric multidimensional scaling,NMDS)进行可视化分析。
采用SPSS 26进行单因素方差分析和Pearson相关性分析,使用Canoco 5.0进行冗余分析。使用FUNGuild对真菌的营养类型和功能类群进行分类,并将Qiime2输出的ASV信息与物种信息合并,附加在ASV表格中。利用R语言中的“FUNGuildR”包和“funguild”数据库进行对比和分析,仅保留置信度为“极可能”和“很可能”两个等级[18]。将被鉴定为复合营养型的群落归并为“其他真菌”,将被鉴定为复合多功能方式的群落统一归类为该营养类型下的“其他病理/腐生真菌”。
2 结果与分析
2.1 土壤的理化性质差异
土壤理化性质分析结果(表2)表明,土壤pH、全碳、全氮、硝态氮、铵态氮、速效钾均在不同林分间差异显著(P<0.05)。其中,混交林中的土壤全碳、全氮、全磷含量均高于其他3种林分。马尾松林的土壤硝态氮、铵态氮含量显著高于其他林分(P<0.05),其他林分之间无显著差异;速效钾则在杉木林中含量最高,显著高于混交林、马尾松林及甜槠林(P<0.05)。土壤碳氮比在4种林分间无显著差异。
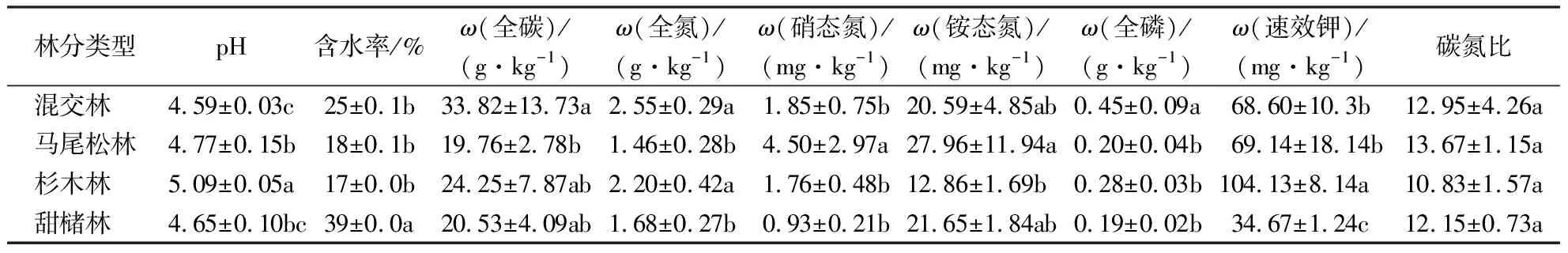
表2 不同林分土壤理化性质(平均值±标准差)1)Table 2 Soil physiochemical properties under different stands (mean±SD)
2.2 土壤的真菌多样性及组成
4种林分土壤样本通过高通量测序,获得1 836 564条高质量序列,共检测到3 141个真菌ASVs,涉及9门19纲44目72科105属。分析结果(图1)表明,杉木林的Shannon指数(6.17)、Simpson指数(0.92)、Chao1指数(593.41)及ASV数量(592.10)均高于其他3种林分,说明杉木林真菌群落丰富度和物种多样性高于马尾松林、混交林及甜槠林。其中:杉木林与马尾松林之间的Shannon指数和Simpson指数没有显著差异,而马尾松林与混交林、甜槠林之间存在显著差异(P<0.05);Chao1指数和ASV数量在杉木林与其他林分之间存在显著差异(P<0.05)。总体上,4种造林模式的真菌群落α多样性表现为:杉木林>马尾松林>混交林>甜槠林。
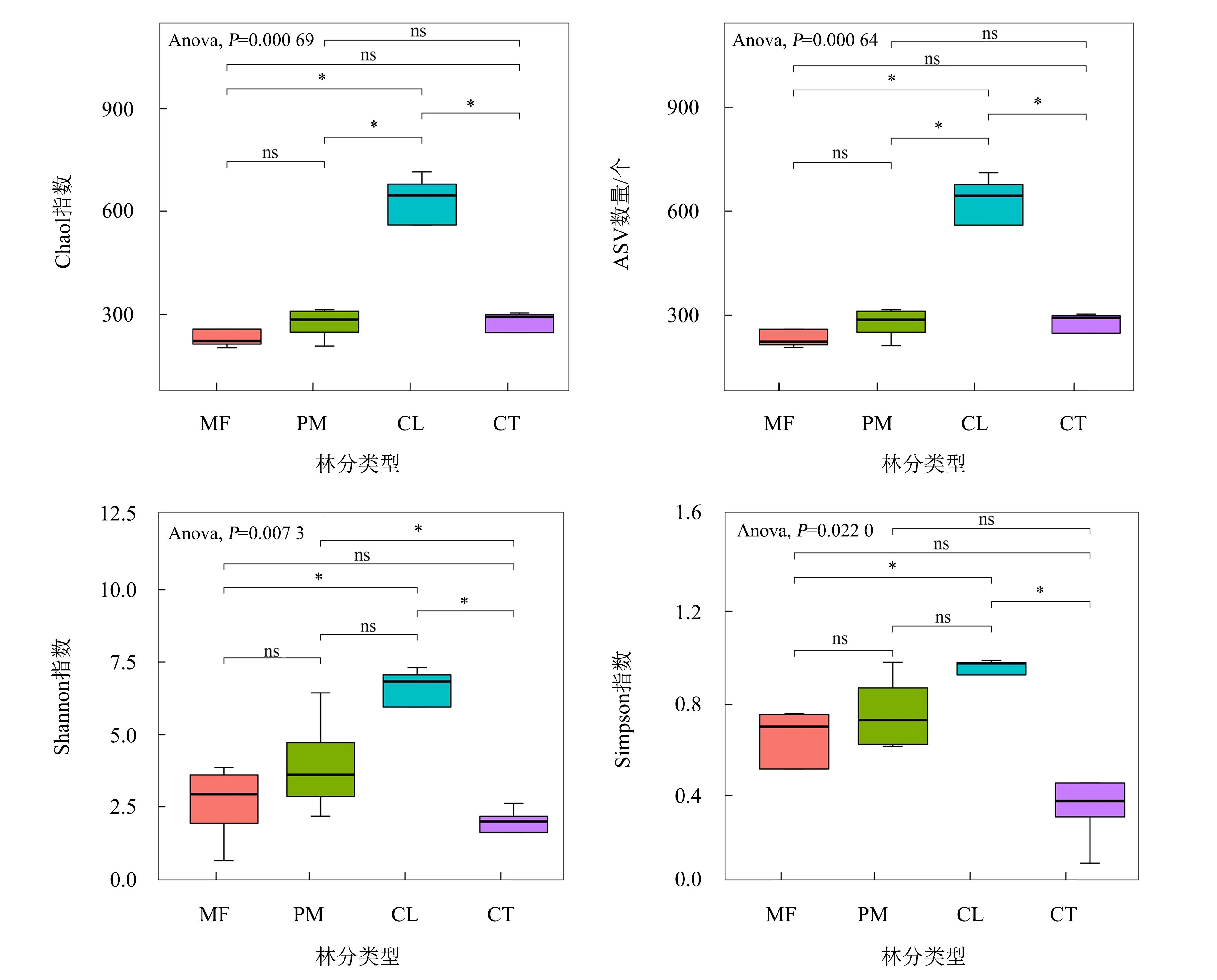
MF:混交林;PM:马尾松人工林;CL:杉木人工林;CT:甜槠林。
基于Bray-Curtis距离矩阵的NMDS分析结果(图2)表明,4种林分下的土壤真菌群落存在一定的差异(Stress=0.090 5)。说明林分组成对森林土壤真菌群落的分布产生了影响。在NMDS1轴上,杉木林与其他3种林分的土壤真菌群落明显分离,表明土壤真菌群落具有明显的特异性。混交林与甜槠林、马尾松林在一定程度上重叠,表明它们具有一些共同的真菌类型。
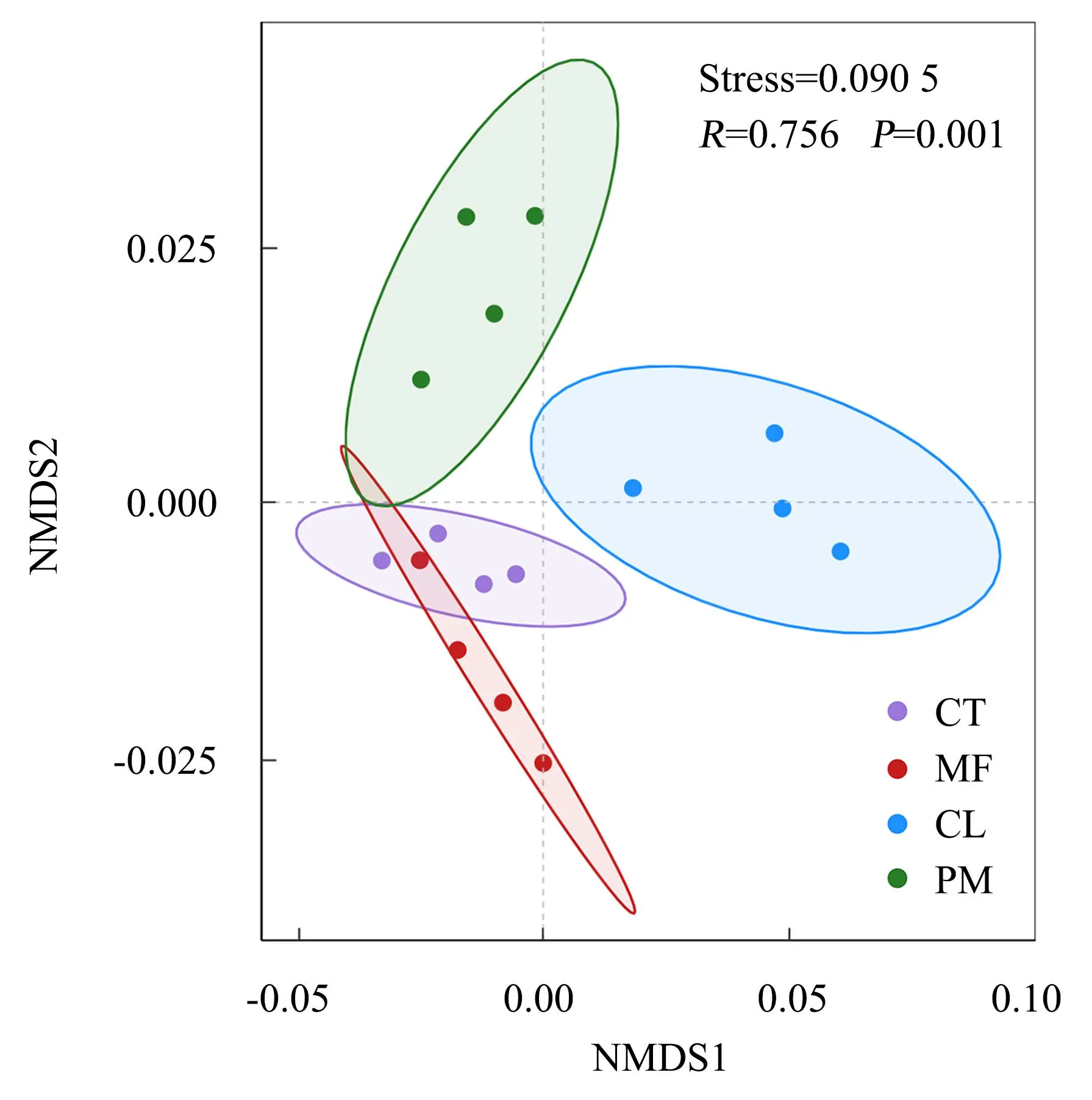
MF:混交林;PM:马尾松人工林;CL:杉木人工林;CT:甜槠林。
在门水平上,4种林分土壤真菌的优势菌门主要是担子菌门Basidiomycota(33.69%~86.56%)和子囊菌门Ascomycota(13.25%~91.87%),其次是被孢霉门Mortierellomycota(0.13%~0.80%)和毛霉菌门Mucoromycota(0.11%~0.66%),其他门的相对丰度很低(<0.10%)(图3A)。担子菌门在混交林中的相对丰度显著高于杉木林,而杉木林中的子囊菌门相对丰度显著高于混交林和马尾松林。在科水平上,平均相对丰度大于1%的科共有10个(图3B),优势真菌科依次为红菇科Russulaceae(0.14%~67.52%),蜡伞科Hygrophoraceae(0.04%~20.97%)、毛霉科Elaphomycetaceae(0%~20.00%)、古根霉菌科Archaeorhizomycetaceae(0.57%~12.75%)、角担菌科Ceratobasidiaceae(0.11%~12.01%)。其中,红菇科在甜槠林和马尾松林中的相对丰度较高,分别占67.52%和56.91%,显著高于杉木林中红菇科的相对丰度0.14%。其次是混交林中的蜡伞科(20.97%)和甜槠林中的毛霉科(20.00%)。而杉木林中的古根霉菌科(12.75%)和角担菌科(12.01%)的相对丰度显著高于其他3种林分。
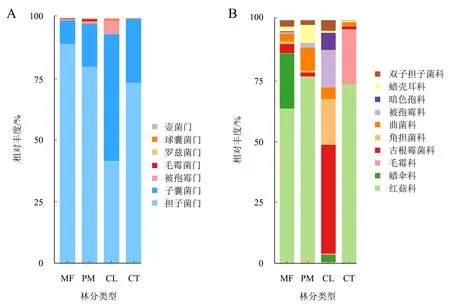
MF:混交林;PM:马尾松人工林;CL:杉木人工林;CT:甜槠林。
2.3 土壤真菌群落功能预测
在4种林分中,土壤真菌的营养类型主要以共生营养型为主,其次是腐生营养型(图4A)。对这些营养类型进行进一步分类,并划分为12个主要功能类群,包括外生菌根真菌(ectomycorrhizal fungi)、丛枝菌根真菌(arbuscular mycorrhizal fungi)、内生真菌(endophyte)、附生真菌(epiphyte)、地衣寄生菌(lichenized fungi)、兰花菌根真菌(orchid mycorrhizal fungi)、植物腐生菌(plant saprotroph)、木质腐生菌(wood saprotroph)、未定义腐生菌(undefined saprotroph)、动物病原菌(animal pathogen)、植物病原菌(plant pathogen)和寄生真菌(fungal parasite)(图4B)。在这些功能类群中,内生真菌、木质腐生菌和未定义腐生菌在4种林分之间存在显著差异(P<0.05)。内生真菌在混交林和甜槠林中的相对丰度分别为12.52%和11.60%,显著高于其在杉木林和马尾松林中的相对丰度(4.95%和8.74%)。杉木林中腐生营养型的相对丰度高于其他林分。
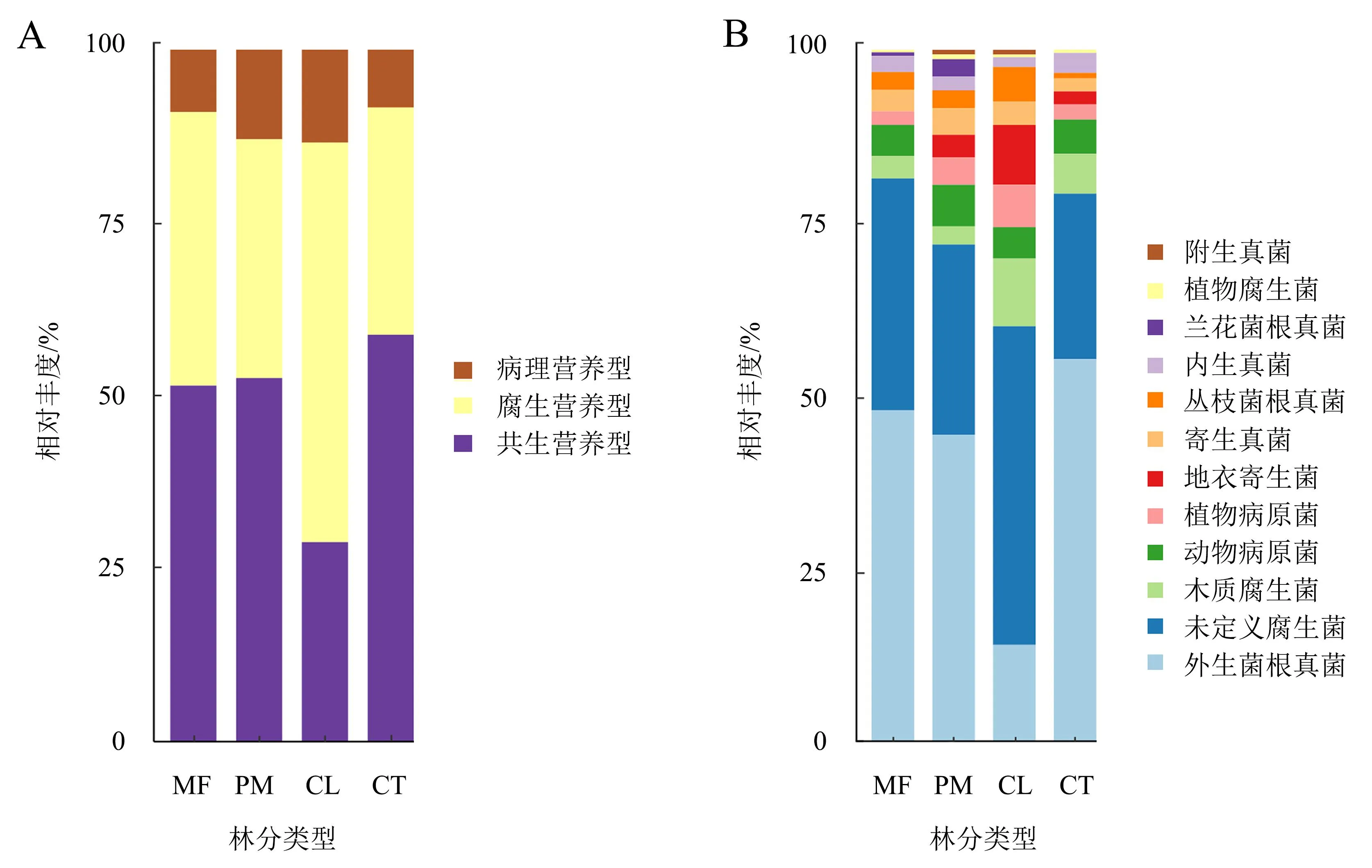
MF:混交林;PM:马尾松林;CL:杉木人工林;CT:甜槠林。
2.4 环境因子与真菌多样性及功能类群的相关性分析
门水平上的冗余分析结果(图5A)显示,土壤pH是影响真菌群落的主要环境因子,其解释率为39.7%。担子菌门与土壤pH呈负相关。

SWC:土壤水分含量;SOC:土壤有机碳;TN:全氮;AK:速效钾;TP:全磷;TN:全氮;铵态氮;硝态氮。
科水平上的冗余分析结果(图5B)表明,第1排序轴(RDA1)和第2排序轴(RDA2)解释量分别为51.26%和9.19%,两者共解释了土壤真菌群落结构变化的60.45%。在环境因子中,只有土壤pH与土壤真菌组成呈显著相关(P=0.002),其解释率为31.5%。受土壤pH影响的菌科包括暗色孢科(Dermateaceae)、角担菌科、被孢霉科(Mortierellaceae)、古根霉菌科。

表3 土壤理化性质与真菌群落多样性及功能类群的相关系数1)Table 3 Correlation coefficients between soil physicochemical properties and diversity and functional groups of fungal communities
3 讨论
3.1 土壤真菌的群落比较
土壤真菌多样性是反映真菌群落总体动态变化的敏感指标,可以作为土壤质量变化的指标[19]。基于高通量测序的分析结果显示,4种林分的土壤真菌群落表现出明显的差异,杉木林的土壤微生物多样性和群落结构与混交林、马尾松林和甜槠林存在显著差异。真菌α多样性的分析结果(图1)显示,杉木林的土壤真菌Shannon指数显著高于混交林和甜槠林,略高于马尾松林,表明杉木林的土壤真菌多样性较高且分布较均匀,而混交林的土壤真菌多样性指数较低。这可能是因为混交林的土壤养分较为充足,降低了植物对菌根的依赖性,从而减少了植物供给共生真菌的营养,导致真菌多样性降低[20]。另外,林下植物组成的差异也会显著影响土壤微生物群落的多样性。NMDS分析结果(图2)显示,土壤真菌群落的分布受到林分组成的影响,说明土壤真菌群落结构与植被林型具有相关性,能够积极响应森林植被类型的演替和变化。
此外,杉木人工林的真菌群落组成与马尾松人工林、甜槠林和混交林不同。这是因为人工林中的真菌群落受到土壤性质、植物群落和小气候等多种因素的影响。不同的植被类型会产生不同的凋落物残体和根系分泌物,而土壤微生物主要利用植物凋落物和根系分泌物作为碳源。因此,不同林分的土壤微生物群落多样性和结构存在差异。在4种林分的土壤中,担子菌门真菌和子囊菌门真菌是优势菌门,这与大多数研究结果[21-23]一致。这两类真菌在土壤中扮演着重要的角色:子囊菌多为腐生真菌,参与有机物质的分解和养分循环[24],通常在酸性土壤中丰富存在;而担子菌可以与植物共生形成菌根,也可以分解土壤中的木质纤维素[25]。此外,林地中的土壤真菌群落组成和结构对树种组成变化的响应较为积极。混交林中的树种组成可以直接影响凋落物的数量和质量,从而对土壤真菌群落的组成和结构产生影响。红菇科作为混交林、马尾松林和甜槠林的优势菌群,在杉木林和其他林分中的相对丰度存在差异。红菇科菌群具有庞大的菌丝系统与寄主植物的根系共生,多活跃于富含有机质的酸性土壤,对植物的生长发育起到促进作用[26]。
森林生态系统中的土壤真菌群落受土壤环境因子影响。在本研究中,担子菌在土壤酸碱度变化时表现出积极的响应,而子囊菌在较高pH的土壤中更为丰富。在特定环境下,大多数土壤真菌更喜欢湿润且呈弱酸性的土壤环境,因此它们对土壤酸碱度变化的响应较为积极。这个结果与Liu et al[27]的研究结果一致。本研究证实了土壤理化性质是影响真菌物种丰度和多样性的主要因素之一。土壤pH和养分条件是土壤真菌群落的关键限制因子。
3.2 土壤真菌的功能群分析
研究区的马尾松林、甜槠林和混交林的真菌群落以共生营养型为主,而杉木林的真菌群落以腐生营养型为优势。这可能与真菌的定殖策略密切相关。然而,作为最早定植于植物残体的菌群,腐生真菌容易受到植被的影响。当腐生和寄生真菌的比例高于共生真菌时,林木的抗逆性会持续减弱,从而引发病害的发生[28]。相反,丰富的共生营养型真菌可以有效提高植物对氮和磷的吸收效率[29],通过拮抗作用对抗植物病原菌,减少病理营养型真菌的产生,抑制病原菌的定殖,从而减轻植物的病害程度[30]。此外,菌根真菌(外生菌根真菌)通过吸收土壤中的营养物质来抑制腐生真菌和其他微生物分解者[31]。因此,建立多树种混交林、引入外来和乡土阔叶树种,增加共生营养型真菌的比例,有助于提升土壤质量和增强林木树种的抗性。
在本研究中,不同林分中的外生菌根真菌的相对丰度存在显著差异。马尾松作为典型的外生菌根树种,其土壤中的外生菌根真菌数量高于以丛枝菌根真菌为主的杉木林。作为与植物根系共生的真菌,外生菌根真菌在提高植物的抗逆能力、调节根际微环境、稳定植物群落等方面发挥着重要作用[20]。当外生菌根真菌的丰度达到一定程度时,它们可以通过改变与植物根系共存的植物组成来推动森林的次生演替过程[32]。
相关性分析显示,部分土壤真菌多样性指数及功能类群的相对丰度与AK密切相关,而其他土壤理化性质与真菌功能类群之间并没有显著相关性。这可能是因为植物根系与根内真菌之间存在相对稳定的供养和反馈关系[33]。根系微生态环境容易受到过滤作用的影响,改变土壤养分性质对真菌组成的影响,从而难以确定土壤理化性质和真菌功能类型组成之间的因果关系。另外,亚热带地区的植被组成复杂,环境因子多样化,影响土壤真菌群落的因素较多。真菌自身的生理活动也会影响土壤生态系统的健康和稳定性。因此,通过调整树种组成、优化林分结构、培育混交林等方法,可以增加土壤真菌功能的多样性,改善森林土壤的生态服务功能。
4 结论
在亚热带地区,不同类型的森林土壤的理化性质存在显著差异(P<0.05)。土壤有机质、全氮和全磷的含量表现为:混交林>杉木人工林>甜槠林>马尾松人工林。杉木林的土壤真菌群落趋于多样化且分布均匀。不同林分的土壤真菌群落优势菌门也存在差异,杉木林以子囊菌门为主,马尾松林、甜槠林和混交林以担子菌门为主。随着森林类型从针叶林向针阔叶混交林转变,真菌的共生营养型与腐生营养型的比例相对减少。土壤pH、速效钾和铵态氮是影响4种林分土壤真菌群落结构的主要土壤因子。土壤理化性质与土壤真菌物种丰度和多样性之间存在密切关系。然而,土壤真菌群落功能与养分的耦合机制仍不清楚,需要通过宏基因组、代谢组等方法对土壤微生物功能与生态系统服务功能之间的关系进行更高效和精确的研究。本研究证实,与纯针叶林相比,针阔叶混交林和阔叶林更有利于该地区土壤养分的积累。
