抗虫辣椒BLTY品系对斜纹夜蛾肠道细菌多样性的影响
2024-03-18贺韶韶庄浩楠袁格格
余 宙, 贺韶韶, 庄浩楠, 袁格格, 陈 功, 于 欢
(湖南农业大学植物病虫害生物学与防控湖南省重点实验室,湖南 长沙 410128)
斜纹夜蛾(SpodopteralituraFabricius)属鳞翅目(Lepidoptera)夜蛾科(Noctuidae),是世界性的农林业主要害虫,在亚洲热带和亚热带地区广泛分布[1-3]。其具有生殖能力强、寄主范围广且食量大的特点,常间歇性大暴发,给农林业生产造成严重危害[4-5]。目前,对斜纹夜蛾的防治仍以药剂防治为主。但是,化学农药的过度使用会导致该虫抗药性增强、人畜安全受到威胁及环境污染加重等问题[6-10]。
昆虫的中肠含有大量的微生物,直接或间接对宿主发挥重要作用,包括分解毒素和杀虫剂、抵御寄生虫和其他病原体、在个体之间传递信号以及生产营养物质和帮助消化等[11-15]。例如:蓝波妙[16]发现,斜纹夜蛾肠道菌群能够编码纤维素降解酶、木聚糖酶和果胶酶,由此说明斜纹夜蛾肠道细菌在宿主营养供给上起重要作用;Gadad et al[17]利用抗生素清除斜纹夜蛾的肠道菌群后发现,该虫对农药的抗性显著降低,揭示了肠道微生物能够介导斜纹夜蛾对农药的抗性。然而,在条件合适的情况下,共生细菌也可以成为致病菌。例如:发光光杆状菌(Photorhabdusluminescens)是共生菌还是病原体,取决于它是寄生在线虫的肠道中还是寄生在昆虫的血淋巴中[18];此外,肠道菌群的组成或位置的紊乱也会导致宿主出现病理状态或死亡现象[19-21]。这些研究均表明斜纹夜蛾肠道菌群相对丰度或结构的改变都可能影响宿主。
本研究团队在前期研究中筛选得到对斜纹夜蛾幼虫具有胃毒性的BLTY品系辣椒。斜纹夜蛾1~3龄幼虫在取食该品系辣椒叶片后出现大规模死亡,存活率不高于22.5%;同时,与取食没有胃毒性的FXBX辣椒叶片相比,幼虫发育历期显著延长,化蛹率和羽化率显著降低[22]。但是,具体的抗虫机制尚不明确。本研究利用基因测序的方法,对取食BLTY品系辣椒叶片后斜纹夜蛾肠道菌群多样性、结构及功能的变化进行探究,为进一步揭示BLTY品系辣椒对斜纹夜蛾的作用机理提供参考。
1 材料与方法
1.1 供试昆虫与植物
斜纹夜蛾采自湖南农业大学耘园实验基地,于人工气候养虫室中用人工饲料[23]饲养4代以上,温度(27±0.5) ℃,相对湿度(70±5)%,光暗比14 h∶10 h,每天定时更换饲料。将待羽化的蛹成对放入养虫笼内,成虫羽化后以10%(体积分数)的蜂蜜水饲养。试虫为3龄幼虫。
供试辣椒品种BLTY2、BLTY2父本(♂BLTY2)、BLTY2母本(♀BLTY2)、FXBX种子均由湖南省蔬菜研究所提供。种子经过消毒待发芽后,移至盛有育苗营养土的32穴育苗盘中。待4叶期时,将幼苗移栽至花盆中,于植物病虫害生物学与防控湖南省重点实验室的温室[温度(25±5) ℃、相对湿度(70±10)%、光暗比12 h∶12 h]中生长。用于试验的所有植物均在温室中生长7周,并具有7片完全展开的真叶,整个栽培过程中不使用任何化学农药。
1.2 样品处理
取同一批次斜纹夜蛾3龄幼虫,分别用BLTY2、♂BLTY2、♀BLTY2、FXBX辣椒叶片以及人工饲料[23]饲喂,48 h后,每组随机选取60头于超净工作台上解剖。解剖工具先经75%(体积分数)酒精清洗灭菌,再经紫外光照射30 min。使用75%(体积分数)酒精和5%(质量分数)次氯酸钠对斜纹夜蛾体表消毒200 s,后迅速用0.15 mol·L-1氯化钠漂洗3次,于冰块上解剖斜纹夜蛾幼虫,取出肠道,将肠道内容物放入1.5 mL离心管中与0.05 mol·L-1磷酸缓冲液混匀,保藏于-80 ℃冰箱冷冻。
1.3 总DNA提取
使用艾科瑞细菌基因组DNA提取试剂盒提取各组样品的肠道总DNA,使用DNA产物纯化试剂盒(DP204,北京天根生化科技有限公司)与酶标仪(Infinite M200 PRO, TECAN)检验DNA的质量和浓度,之后保存于-20 ℃冰箱待用。
1.4 文库构建和测序
采用通用引物27F(AGAGTTTGATCCTGGCTCAG)和1492R(GGTTACCTTGTTACGACTT),在96孔梯度PCR仪(莫纳生物科技有限公司)上对基因片段进行聚合酶链式反应(polymerase chain reaction, PCR)扩增。反应总体系25 μL(PCR buffer 2.50 μL,dNTP 0.50 μL,上、下游引物各1.00 μL,TaqDNA polymerase 0.15 μL,模板DNA 2.00 μL,ddH2O 17.85 μL)。反应程序:94 ℃预变性5.0 min,变性50 s;在45~60 ℃下退火50 s、72 ℃延伸10 min,27个循环,最后产物于4 ℃保存。用2%(质量分数)琼脂糖凝胶回收2 μL的PCR产物,利用琼脂糖凝胶纯化回收试剂盒(Aidlab公司)切胶纯化PCR产物。根据纯化PCR产物的条带亮度和浓度计算混样体积并建库,每个样品的PCR产物加入量为30 ng,使用Agilent 2100 Bioanalyzer对文库的片段范围和浓度进行检测。
1.5 肠道样本测序数据处理及物种组成分析
将合格文库送往深圳华大基因科技有限公司进行16S rRNA测序。主要步骤如下:将HiSeq平台测序所得的原始reads进行过滤得到clean data;利用软件FLASH(Fast Length Adjustment of Short Reads, v1.2.11)将reads进行拼接,利用重叠关系将双末端测序得到的成对reads组装成一条序列,得到高变区的tags[24];使用软件USEARCH(v7.0.1090_i86linux32)将拼接好的tags按照序列相似性聚类生成操作分类单元(operational taxonomic units, OTU)[25];得到OTU代表序列后,利用RDP classifer(1.9.1)软件通过比对OTU代表序列与RDP数据库进行物种注释(置信度阈值0.6)。
1.6 肠道微生物多样性分析
基于Illumination HiSeq平台(https://www.illumina.com.cn)进行微生物多样性分析。从样本中随机抽取序列预测不同测序深度下的物种丰富度,从而对斜纹夜蛾肠道菌群样本数量进行评估,参考岳雪华等[26]的方法绘制物种稀释性曲线。应用软件Venny 2.1基于OTU数目作韦恩图,得到各样本特有和共有的OTU数目。利用R语言v3.4.1软件分析斜纹夜蛾肠道细菌的Alpha多样性,包括覆盖度指数、Shannon指数和Chao指数[27]。分别利用unweighted unifrac算法和binary-jaccard算法计算肠道细菌的Beta多样性,利用主坐标分析降维并进行可视化,包括PCoA主成分分析、菌群分型分析和PCA主成分回归分析[28]。利用QIIME 1.9.1软件(http://qiime.org/install/index.html)生成门水平和属水平的相对丰度图。通过LEfSe聚类分析对科水平和目水平的菌群结构进行统计检验和差异分析[29]。利用PICRUSt2软件对测序结果进行KEGG功能预测分析[30]。
2 结果与分析
2.1 肠道菌群样本数量评估
从物种稀释性曲线(图1)可知,随着样本测序数量的增大,曲线由最开始的急增逐步趋于平缓,表明斜纹夜蛾3龄幼虫肠道的菌群信息绝大多数被测得。随着测序数量的持续增大,检测到的物种数不再增加,说明本次测序量充足,能够反映所测细菌的物种广度,可用于后续分析。
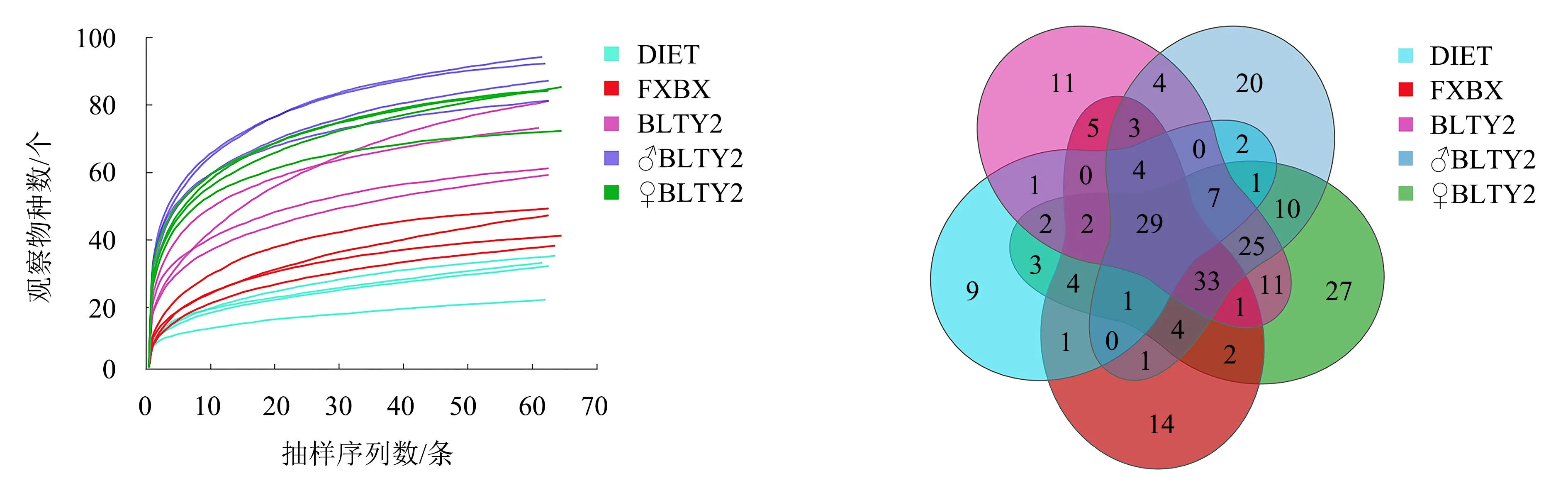
DIET为人工饲料,FXBX为无毒性辣椒叶片,其余为胃毒性辣椒叶片。
2.2 肠道细菌多样性分析
2.2.1 OTU数韦恩图 按照97%序列相似性聚类共得到237个OTU。其中:5种处理的斜纹夜蛾幼虫肠道细菌共有OTU数为29个,占细菌OTU总数的12.24%;取食BLTY2、♂BLTY2、♀BLTY2、FXBX辣椒叶片以及人工饲料的斜纹夜蛾幼虫肠道细菌特有OTU数分别为11、20、27、14、9个,分别占细菌OTU总数的4.64%、8.44%、11.39%、5.91%、3.80%(图2)。可见,取食辣椒叶片后的特有OTU数多于取食人工饲料的OTU数。
2.2.2 Alpha多样性 取食不同食物的斜纹夜蛾肠道细菌覆盖度指数介于0.999 7~0.100 0之间(图3A),表明样本中序列被测出的概率很高,测序结果能代表样本中菌群的真实情况。取食BLTY品系辣椒叶片的斜纹夜蛾幼虫肠道细菌群落的Shannon指数(图3B)与Chao指数(图3C)均显著高于取食FXBX辣椒叶片和人工饲料的样品,说明取食BLTY品系辣椒叶片后斜纹夜蛾肠道细菌的多样性和丰度均显著上升。
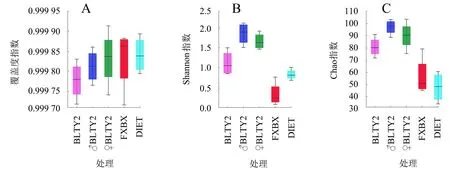
A.覆盖度指数箱线图;B.Shannon指数箱线图;C.Chao指数箱线图。DIET为人工饲料,FXBX为无毒性辣椒叶片,其余为胃毒性辣椒叶片。
2.2.3 Beta多样性 通过R语言统计分析和作图,基于unweighted unifrac距离矩阵的PCoA分析表明,取食3个BLTY品系辣椒叶片的斜纹夜蛾幼虫肠道菌群组成较相似(图4A),且与取食FXBX辣椒叶片和人工饲料的差异较大。菌群分型分析将取食BLTY品系的3组肠道菌群聚合在一起(图4B),表明3组样品的菌群组成相似,与PCoA分析结果相符。 PCA分析表明,取食BLTY品系辣椒叶片的3组样品的肠道菌群组成与取食人工饲料的样品发生了两个方向的偏移(图4C),且FXBX组与饲料组之间也存在偏移,这与PCoA分析结果一致。
2.3 肠道细菌群落物种组成分析
在门水平上,相对丰度前10的菌门如图5A所示,厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)为5组斜纹夜蛾肠道细菌的共有优势菌门。在取食BLTY2、♂BLTY2、♀BLTY2、FXBX和人工饲料的斜纹夜蛾肠道细菌中,厚壁菌门的相对丰度依次为78.23%、59.12%、69.18%、92.07%和84.10%,变形菌门的相对丰度分别为17.96%、31.41%、25.20%、4.10%和15.80%。与取食人工饲料相比,取食BLTY品系辣椒叶片后斜纹夜蛾幼虫肠道细菌中厚壁菌门的相对丰度下降,而变形菌门、放线菌门(Actinobacteria)和拟杆菌门(Bacteroidetes)的相对丰度显著上升。
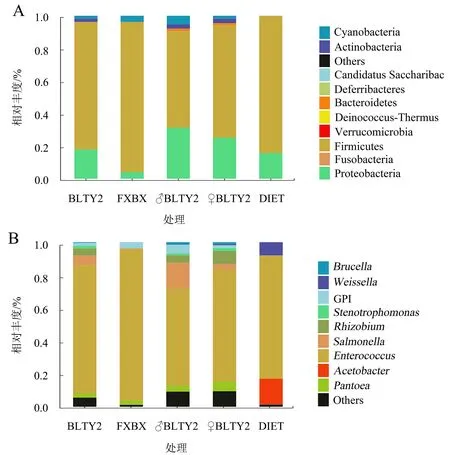
DIET为人工饲料,FXBX为无毒性辣椒叶片,其余为胃毒性辣椒叶片。
在属水平上,5组肠道细菌样本共获得131个菌属。取食BLTY2、♂BLTY2、♀BLTY2、FXBX辣椒叶片和人工饲料的斜纹夜蛾肠道细菌的菌属分别有88、89、98、53、39个,可见,取食BLTY品系辣椒叶片后的菌属多样性明显丰富。其中,相对丰度前10的菌属如图5B所示,与取食FXBX辣椒叶片(91.99%)和人工饲料(78.06%)相比,取食BLTY品系辣椒叶片的斜纹夜蛾肠道细菌中肠球菌属(Enterococcus)相对丰度(74.92%、58.93%、68.24%)下降,但沙门氏菌属(Salmonella)、寡养单胞菌属(Stenotrophomonas)和布鲁氏菌属(Brucella)相对丰度显著上升,而这3种菌属中多存在致病菌。综上,取食BLTY品系辣椒叶片后斜纹夜蛾幼虫的肠道菌群发生了显著变化。
2.4 LEfSe聚类分析
通过LEfSe聚类分析(图6)发现,与取食FXBX辣椒叶片和人工饲料相比,取食BLTY品系辣椒叶片的斜纹夜蛾幼虫肠道菌群中,放线菌目(Actinomycetales)、黄杆菌目(Flavobacteriales)、鞘氨醇杆菌目(Sphingobacteriales)、肠杆菌目(Enterobacteriales)、根瘤菌目(Rhizobiales)、伯克氏菌目(Burkholderiales)、黄单胞菌目(Xanthomonadales)代表的biomarker节点显著增大。与取食FXBX相比,取食人工饲料后斜纹夜蛾幼虫肠道菌群中明串球菌科(Leuconostocaceae)和乳杆菌科(Lactobacillaceae)代表的节点显著增大,且红螺菌目(Rhodospirillales)为其特有菌目;而与取食人工饲料相比,取食FXBX的斜纹夜蛾肠道菌群中肠球菌科(Enterococcaceae)代表的节点显著增大。可见,取食BLTY品系辣椒叶片后斜纹夜蛾幼虫肠道菌群在目水平上发生了较大变化。
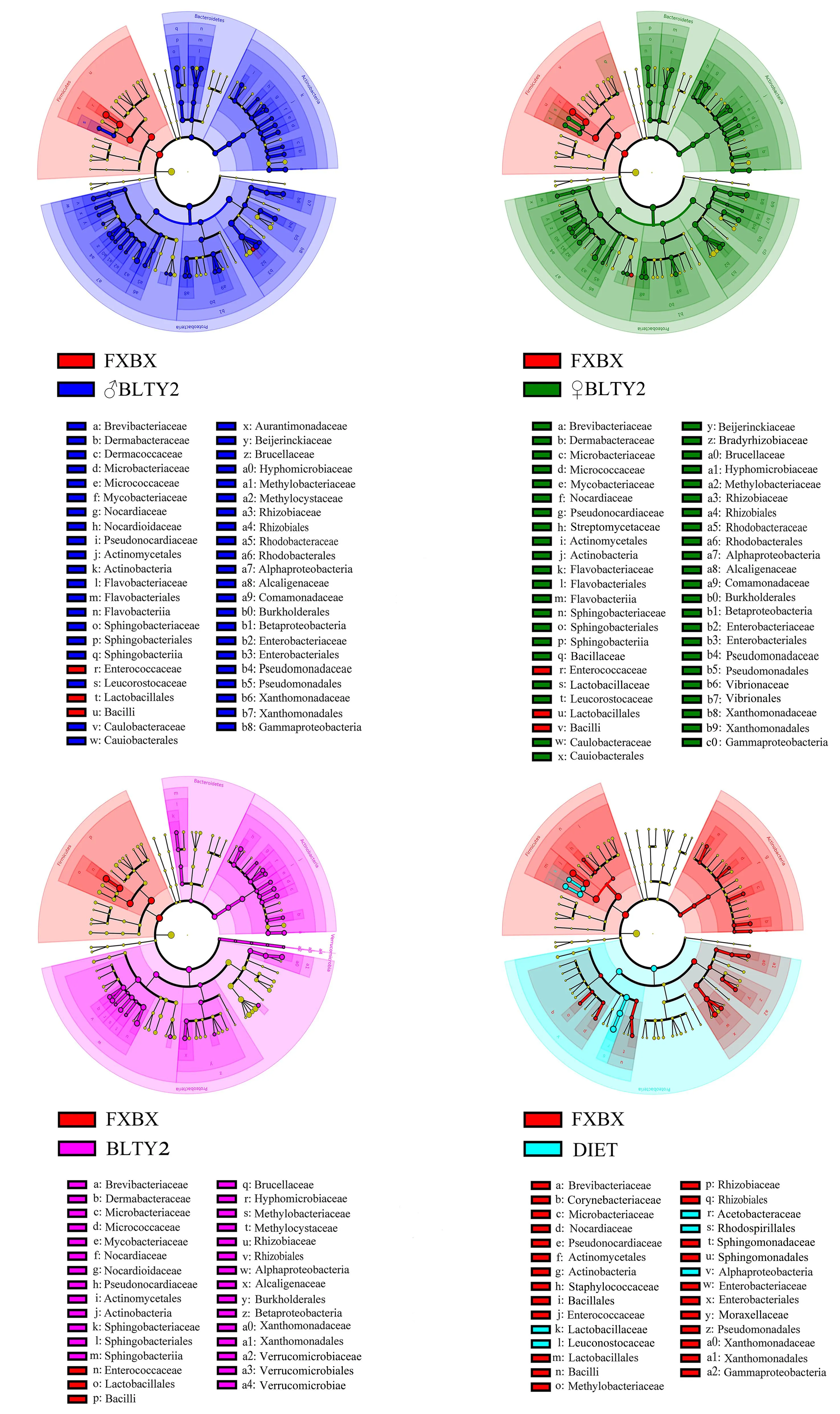
DIET为人工饲料,FXBX为无毒性辣椒叶片,其余为胃毒性辣椒叶片。
2.5 肠道细菌功能预测
KEGG功能预测分析显示,取食BLTY品系辣椒后斜纹夜蛾肠道细菌的一级分类水平代谢通路包括新陈代谢、遗传信息处理、环境信息处理、细胞转化、人类疾病和生物系统,与取食FXBX辣椒叶片和人工饲料相比,取食BLTY品系辣椒叶片的斜纹夜蛾幼虫肠道细菌在细胞转化和新陈代谢通路的相对丰度明显升高(图7A)。进一步对肠道细菌代谢通路的二级分类水平分析显示,与取食人工饲料相比,取食BLTY品系辣椒叶片后,斜纹夜蛾幼虫肠道细菌在细胞运动、运输和分解代谢、传染病、信号传导、外源化合物降解和代谢以及内分泌系统通路的相对丰度显著上升(图7B)。
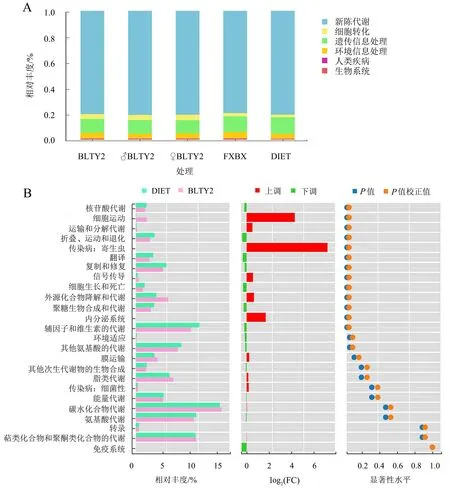
DIET为人工饲料,FXBX为无毒性辣椒叶片,其余为胃毒性辣椒叶片。
3 讨论
昆虫肠道细菌具有丰富的多样性,并与昆虫的健康状况密切相关,且受宿主所处的环境条件、饮食方式和结构、年龄和遗传关系等诸多因素影响[30-35]。本研究发现,取食具有胃毒性的BLTY品系辣椒叶片、无毒性的FXBX辣椒叶片以及人工饲料的斜纹夜蛾幼虫肠道细菌的OTU数目、菌属丰富度、多样性指数和功能均存在差异,其中,取食3个BLTY品系辣椒叶片的菌群组成较相似,并且比取食FXBX辣椒叶片和人工饲料的菌群组成更复杂多样。类似的情况也存在于胃毒性农药处理后的鳞翅目昆虫中:张靳宜等[36]发现,甲氨基阿维菌素苯甲酸盐会对斜纹夜蛾幼虫肠道菌群的结构和代谢能力产生明显影响,使原先不占据优势的棒状杆菌属(Corynebacteriu)和甲醇杆菌属(Methylobacterium)丰度明显上升,由杆菌代谢产物的积累促进毒性反应;姜笑维等[37]对棉铃虫(Helicoverpaarmigera)饲喂含拟除虫菊酯类杀虫剂的饲料后,发现致病菌群相对丰度显著增加;Zeng et al[38]发现,阿维菌素胁迫显著增加了舞毒蛾(Lymantriadispar)幼虫肠道中的肠杆菌属(Enterobacter)、念珠菌属(Candida)和链格孢菌属(Alternaria)3种条件致病菌群的丰度。由此推测BLTY品系辣椒作为抗虫品种,叶片中可能含有特殊的次生代谢物,破坏了斜纹夜蛾幼虫肠道菌群原有的稳态平衡和动态调控机制,引起代谢通路的显著变化,从而对斜纹夜蛾幼虫产生胃毒作用。
本研究还发现,与取食FXBX辣椒叶片和人工饲料相比,取食BLTY品系辣椒叶片后斜纹夜蛾3龄幼虫肠道中肠球菌属相对丰度下降,但沙门氏菌属、寡养单胞菌属和布鲁氏菌属相对丰度显著上升。肠球菌属是昆虫体内一类常见的细菌,一般不会对宿主产生致病性[39-40]。刘语涵[41]研究发现,肠球菌属大量存在于家蚕(Bombyxmori)肠道内,可产生淀粉、脂肪、纤维素等多种酶,促进家蚕对桑叶营养物质的吸收利用;肠球菌属在维持肠道菌群微生态稳定中也发挥着重要功能,特别是对微孢子虫的孢子萌发有明显的抑制效果[42]。沙门氏菌属是导致人类急性食源性疾病的重要人畜共患病原菌[43];宿主免疫功能低下时,可能会由于沙门氏菌进入血液引发严重的菌血症而死亡[44]。寡养单胞菌属为条件致病菌,在宿主免疫力低下时易引起呼吸系统疾病,常与其他条件致病菌混合感染[45-46]。布鲁氏菌属侵入机体后会使机体出现黏膜损伤、消化道感染等症状并引发炎症[47-48]。由此可见,取食BLTY品系辣椒叶片使斜纹夜蛾肠道有益菌的相对丰度下降,而致病菌的相对丰度增加,这可能是其重要的胃毒机制之一。此外,有研究发现,肠道菌群可以与植物代谢物共同作用介导宿主的病理状态。如Mason et al[49]研究发现,玉米中的凝集素几丁质酶可与草地贪夜蛾(Spodopterafrugiperda)幼虫肠道内的肠杆菌共同作用,诱导发生肠道渗漏综合症而致该虫死亡。本研究中,取食BLTY品系辣椒叶片后斜纹夜蛾肠道菌群发生了显著变化,这些变化的菌群与BLTY品系辣椒叶片中的特殊代谢物共同作用,可能也是引起斜纹夜蛾幼虫死亡的因素之一。
综上,BLTY品系辣椒可改变斜纹夜蛾幼虫肠道菌群的结构,主要使有益菌的相对丰度降低和致病菌的相对丰度升高,这可能是BLTY品系辣椒对斜纹夜蛾幼虫的胃毒机制,但该机制与BLTY品系辣椒中的何种代谢物质相关还需要进一步研究。
