利用腐生线虫加速替代疫木中松材线虫种群数量研究
2022-11-29叶建仁史丽娜
王 磊,叶建仁,史丽娜
(南京林业大学林学院,南方现代林业协同创新中心,江苏 南京 210037)
松材线虫病是由松材线虫(Bursaphelenchusxylophilus)引起的一种危险性森林病害,在我国乡土松种上是一种典型的病原主导性病害,大多数寄主植物一旦感病很快枯萎死亡[1-2]。1982年在南京首次发生后,已经陆续在我国19个省市传播和蔓延[3]。由于我国松树种类多、面积大且分布广,森林资源和生态环境遭受了巨大的破坏,经济损失达数千亿元[4-6]。
防控松材线虫病主要有检疫和疫情监测、疫木除治、媒介昆虫防治等。松材线虫病一旦被检疫或监测发现,疫木除治就成为防控的核心措施。此时病原线虫和媒介昆虫都集中在疫木中,只要彻底杀灭疫木中的天牛和线虫,病害就可以得到防控[6]。2018年以前,带有松材线虫或松褐天牛(松墨天牛,Monochamusalternatus)的松树砍伐后的处理手段有焚烧、熏蒸、热处理、微波处理、切片、制板等[7-9]。由于这些疫木处理措施监管漏洞多,一些实际未经处理的疫木常被人为传播到远近不等的地区,引发病害在更多的区域发生与蔓延[10]。2018年后国家进一步严格疫木处理,要求病死树必须在山上或山下就近就地尽快粉碎处理或烧毁[11]。此要求可有效减少疫木处理过程中的管控漏洞,但实际防控过程中出现了人力、财力、实际操作难以落实的状况。我国南北森林环境变化大,许多情况下机械或人力作用有限。因此,研究探索简易有效的松材线虫病疫木就地除害技术对于控制我国松材线虫病的蔓延扩散意义重大。
在松材线虫病疫木中除了松材线虫,往往还存在其他寄生性植物线虫、食真菌线虫和食腐性线虫。由于生存空间、食物竞争、释放抗生类物质等因素,线虫在种群间会发生争夺资源、产生竞争,抑制其他种群生长发育的现象[12-15]。腐生线虫是一类以细菌作用的糖和蛋白质分解产物为食料的线虫,不含口针但能靠取食腐殖质或某些微生物进行生长发育[16-17]。国内外只有个别学者对枯死松树体内的腐生线虫进行过初步研究,松材线虫病疫木在发病后期,通常会出现松材线虫数量不断减少,腐生线虫数量不断增加的现象[18-19]。虽然松材线虫病往往发生在冬季,腐生线虫的生长繁殖可能会受低温限制难以应用到实际生产上,但已有研究发现在自然条件下腐生线虫仍会对松材线虫具有一定抑制作用[20]。基于此,本研究进一步探讨不同种类腐生线虫对病疫木中松材线虫种群数量的影响,旨在筛选出能够加速松材线虫种群消亡的腐生线虫种类,探索利用高效的腐生线虫替代技术就地处理松材线虫病疫木,从源头阻断松材线虫病的传播与扩散。
1 材料与方法
1.1 试验材料
分离腐生线虫的疫木来自国内5个发生松材线虫病的疫区,分别是安徽省池州市九华山风景区(117°8′E,30°5′N)、黄山市黄山风景区(118°1′E,30°1′N),四川省巴中市巴州区水宁寺镇(107°1′E,31°49′N),江苏省南京市中山陵(118°51′E,32°04′N)、宿迁市宿城区(118°17′E,33°58′N),均属于亚热带季风气候区,四季分明、温暖湿润、雨量充沛、光照充足,霜期较短,冬季平均气温为0℃以上。在这些疫区染病死亡的黑松(Pinusthunbergii)和马尾松(Pinusmassoniana)上采集木样,用于腐生线虫的分离。腐生线虫替代松材线虫试验的疫木来自江苏句容林场摇令口工区(119°12′E,32°05′N)当年感染松材线虫病的15年生黑松林。
1.2 试验方法
1.2.1 腐生线虫与松材线虫的主要形态区别
在光学显微镜下观察,腐生线虫的口腔内不含口针,松材线虫的口腔内含有口针;腐生线虫的食道类型通常是小杆型或双胃型,松材线虫的食道类型是滑刃型。
1.2.2 腐生线虫的纯化培养
将漏斗放入灭菌锅内,105 ℃灭菌10 min后,取每份木样的木质部,劈成细木条,用贝尔曼漏斗法[21]静置分离12 h后,再用15 mL离心管收集其滤液。在显微镜下观察分离得到的线虫液,用移液枪挑取25条以上形态相似的腐生线虫成虫,置于腐生线虫培养基的平板上,25 ℃下培养14 d后再次用贝尔曼漏斗法收集线虫,进行纯化培养。
腐生线虫培养基配方:黑松木屑30 g、葡萄糖20 g、琼脂30 g、水1 L,121 ℃灭菌25 min[22]。
1.2.3 腐生线虫在培养基上的繁殖力测定
将每种腐生线虫在腐生线虫培养基平板上各接种3皿,每皿接种200条,置于25 ℃恒温培养箱中培养14 d。采用贝尔曼漏斗法分离培养基中的腐生线虫,在光学显微镜下计数,最后统计各种腐生线虫在培养基上的扩增数量情况。
1.2.4 室内条件下腐生线虫与松材线虫在疫木块内种群竞争试验
将感病黑松主干均匀切割成3.5 cm厚的圆盘,从形态学上部开始由高到低依次对圆盘进行编号,带回南京林业大学森林病理实验室。根据相邻圆盘大小选择3个圆盘,将圆盘边材锯成边长2.5 cm的小木块,每个圆盘选择18个小木块,将这54个小木块作为每个温度每种虫株处理的接种样品。由于松材线虫数量在距离地面不同高度的圆盘的边材之间差异较大,并且每一圆盘的边材体积有限,为了尽量减少试验误差,对每一温度每种虫株都安排了相应的对照组。将木块分为9列,前3列为对照组,处理时间分别设置为0、15、30 d,后6列的处理时间分别为3个接种梯度的15、30 d,每列6个重复。
接种前先记录木块鲜质量,然后用电钻在木块中央钻取1个直径0.5 cm、深1.0 cm的小孔,将灭菌的湿棉花置于接种孔底部,再在孔内滴加无菌水和腐生线虫悬浮液,滴加液体的总体积为600~700 μL,以对照组疫木块中松材线虫数量的平均值作为基础值,将腐生线虫按照1、2、4倍(接种体积分别为100、200、400 μL)于该基础值数量接种至处理木块中。接种腐生线虫在湿棉花上后,立刻再用湿棉花覆盖接种孔,并用封口膜包裹接种孔。本试验共接种8种腐生线虫,分别置15和25 ℃恒温培养箱中培养15和30 d后,使用贝尔曼漏斗法分离线虫并统计松材线虫和腐生线虫的数量。按如下公式计算木块中松材线虫减少率:
式中:x1表示试样木块的松材线虫减少率,%;n0表示试样木块0 d时(对照组)的初始松材线虫数量,条/g;n1表示试样木块在接种后15或30 d时的松材线虫数量,条/g。
1.2.5 自然条件下腐生线虫种群与松材线虫种群间竞争试验
于2019年12月在江苏省句容林场进行野外接种试验。用油锯将当年染病的松材线虫病疫木主干分成约75 cm长的木段并逐一编号。每个木段设置4个接种孔,每个孔深度为50 mm、直径35 mm。将湿棉花置于每个孔的底部,将腐生线虫悬浮液接种至棉花上,并在孔上方覆盖一层湿棉花后用保鲜膜包裹住接种口。试验设置8种腐生线虫处理,每处理12个重复,每重复的接种量为2.0×105~2.5×105条,以接种无菌水的木段作为对照。2020年 4月,即120 d后取样,每个木段设置5个取样点,位于接种孔附近,用电钻钻取木屑,装入信封带回实验室,使用贝尔曼漏斗法分离。回收漏斗内的木屑装入信封,放入105 ℃烘箱中烘干至质量恒定。在天牛羽化前,即150 d后,将木段分成两部分。一部分用于收集天牛蛹室周围0.5~1.0 cm木样,另一部分用铁丝网罩住,等待天牛羽化后捕获天牛。蛹室周围木样和天牛均带回实验室使用贝尔曼漏斗法分离并统计线虫数量。
1.3 数据处理
利用 Excel 2010和SPSS 24.0处理和分析数据,Prism 6.0软件作图。
2 结果与分析
2.1 疫木中分离的腐生线虫及在培养基中的繁殖速率
采用贝尔曼漏斗分离法,共分离到8种腐生线虫,具体虫株及其整体形态特征见图1。
根据采样疫区名称将8种腐生线虫分别编号为虫株JHS1、JHS2、JHS4、HS2、ZSL1、SN1(♀)、SQ1(♀)、SQ3(♀)。腐生线虫JHS1、JHS2、JHS4、HS2和ZSL1均含有雌雄虫,雄虫的体长均稍小于雌虫。SN1、SQ1、SQ3均未发现有雄虫,可能存在两性同体生殖或孤雌生殖,即卵和精子都由同一母体产生,通过自体受精而繁殖;也可能存在性成熟的雄虫在和雌虫交配后很快死亡的情况,所以未检出雄虫。在具体形态上,腐生线虫的后口腔壁上可能带有2~3个疣突或杆状的小齿;食道为小杆型或双胃型,未见食道腺;尾部均为圆锥形,但尾型在不同种的虫株间往往差异明显;雌虫的生殖腺单个或1对;雄虫的尾部有的有交合伞,有的不具备,但都具备6~9对生殖乳突。腐生线虫培养14 d的繁殖力情况见表1,由表1可知,在木屑培养基平板上培养14 d时,虫株SQ3和ZSL1的扩增倍数明显高于其他处理组,分别为239.9和136.3倍;其次是虫株HS2和SQ1,分别为52.9和34.8倍。而虫株JHS1、JHS2、JHS4和SN1在培养基上的繁殖力情况表现都一般,扩增倍数分别为12.5、11.7、10.5和17.3倍,相互之间不存在显著差异。可见,木屑培养基适合多数腐生线虫的前期快速扩繁工作。
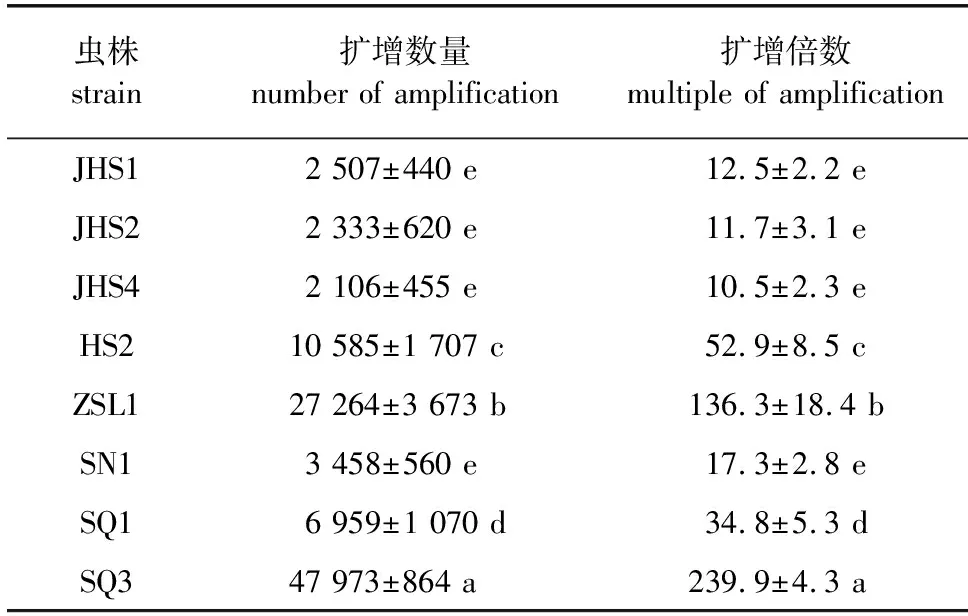
表1 腐生线虫培养14 d的繁殖力情况
2.2 室内条件下腐生线虫与松材线虫在疫木中的竞争
2.2.1 在15 ℃条件下腐生线虫对松材线虫的抑制作用
在15 ℃条件下培养15 和30 d后,8种腐生线虫在不同接种量的疫木块中均能被分离到。分析结果(图2)表明,接种量越大的疫木块中腐生线虫初期定殖的密度就越大,但随着时间延长8种腐生线虫数量均逐渐降低,有的降低很快,有的降低到一定程度后又有上升。
其中虫株ZSL1和JHS2在30 d时仍能保持较高的种群数量。而且发现在30 d时,接种量为400 μL 的虫株JHS2处理与对照相比,松材线虫种群数量显著减少(P<0.05),松材线虫减少率为89.27%,与对照相比减少率提高19.05%;接种量为100 μL的虫株JHS4处理组的木块中松材线虫减少率为90.93%,与对照相比提高14.08%(图3)。而在虫株SN1和SQ3处理中出现松材线虫减少率为负值的情况,可能是由于这两处理的接种木块在取材时处于松材线虫感染中后期,此时松材线虫繁殖旺盛、具有明显的种群优势。
2.2.2 在25 ℃条件下腐生线虫对松材线虫的抑制作用
在25 ℃条件下培养15 和30 d后,8种腐生线虫在不同接种量下均能从木块中分离得到,初期疫木中腐生线虫数量也与接种量相关,但很快就逐渐减少,其中6种虫株在30 d时,已经降到很低的数量,只有虫株SQ3和JHS4依然能够保持一定种群数量(图4)。
培养15 和30 d后,对木块中松材线虫的种群数量进行分析,结果表明(图5),25 ℃条件下培养15 和30 d后,与对照相比,在8个处理中虫株JHS4表现最为突出(P<0.05)。在15 d时,虫株JHS4处理组的3种不同接种量的木块中松材线虫减少率分别为75.30%、63.69% 和78.40%,均显著高于对照组的39.88%(P<0.05)。在培养30 d时,接种量为400 μL的虫株JHS4处理中松材线虫减少率为44.05%,与对照相比差异显著(P<0.05)。30 d时虫株JHS4处理疫木块中松材线虫的数量与15 d时相比明显增加,可能是因为前15天时间内虫株JHS4在疫木块中的存活数量相对较多,能显著抑制松材线虫生长繁殖,后15天内虫株JHS4在疫木块中的种群数量虽然还保持在一定水平,但总体数量已经不如刚接种时的存活数量,所以对松材线虫繁殖的抑制效果略有减弱。
2.3 野外条件下腐生线虫与松材线虫在疫木中的竞争
2.3.1 接种腐生线虫对木段内松材线虫种群数量的影响
林间野外试验结果(图6)表明,接种试验120 d后,所有处理组包括对照中松材线虫的数量均在减少。但是,接种虫株JHS1、JHS2、JHS4、HS2和SN1处理组的木段内松材线虫减少率均高于对照,其中接种虫株JHS4的木段内松材线虫的减少率为96%,与对照(74%)相比差异显著(P<0.05)。
2.3.2 接种腐生线虫对羽化天牛及其蛹室周围携带松材线虫数量的影响
从2020年5月底到7月初,从林间放置的网罩中抓取羽化后的松褐天牛(松墨天牛),分离携带的松材线虫并统计数量,结果如表2所示。用不同的腐生线虫处理木段后,羽化天牛携带的松材线虫数量差异较大。对照组内每头天牛平均携带986条松材线虫,在接种虫株JHS4处理中天牛携带的松材线虫数量最少,平均每头天牛携带的松材线虫数量为132条,为对照的13.39%。其次为虫株ZSL1和JHS2处理,平均每头天牛携带的松材线虫数量分别为150和262条,为对照的15.21%和26.57%。
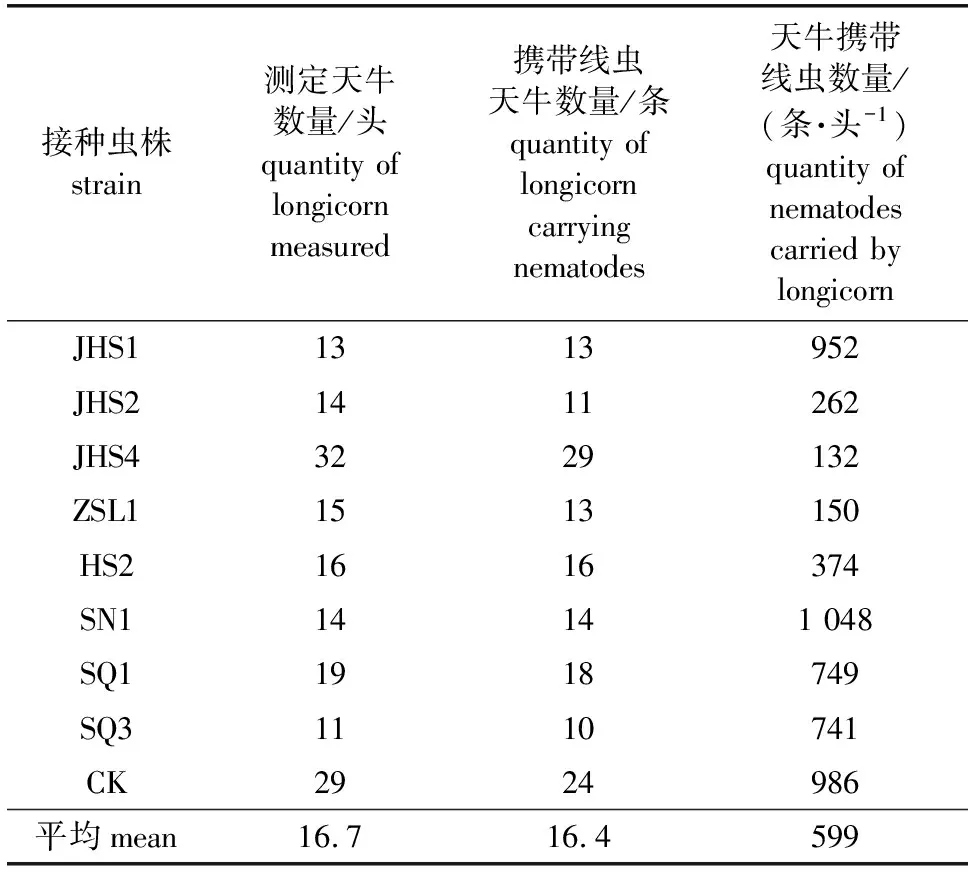
表2 接种不同腐生线虫对羽化松褐天牛携带松材线虫种群数量的影响
2020年5月上旬在天牛羽化前,收集天牛蛹室周围的木样, 分离其中松材线虫数量。结果如表3所示。由表3可知,所有处理组蛹室周围木样所携带的松材线虫数量均低于对照。其中,接种虫株JHS4处理组蛹室周围木样携带的松材线虫数量最低,为20条/g,是对照的25.64%;其次为虫株ZSL1处理组,携带松材线虫的数量为24条/g,是对照的30.77%。在镜检时发现虫株JHS2、JHS4、ZSL1、HS2和SQ3这5种腐生线虫普遍存在于相应处理组的蛹室中。
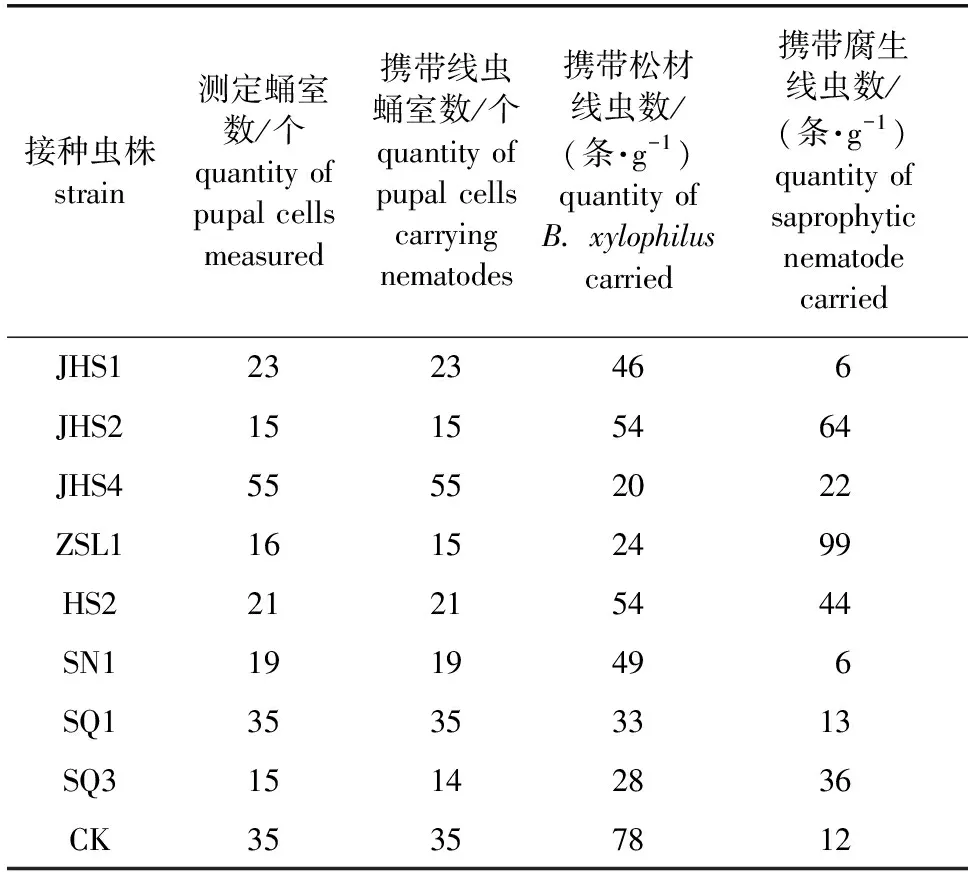
表3 接种不同腐生线虫对天牛蛹室携带松材线虫种群数量的影响
3 讨 论
通过接种腐生线虫至含有松材线虫的病疫木中,在室内及野外条件下进行腐生线虫与松材线虫的种间竞争试验,筛选能够加速松材线虫种群消亡的腐生线虫虫株。研究发现虫株JHS2、JSH4、ZSL1可在不同的条件下抑制松材线虫的种群数量,具有一定的生物防治应用潜力。其中虫株JSH2和JSH4在室内及野外条件下对松材线虫种群数量均有抑制作用;虫株ZSL1仅在野外接种试验中表现出了明显抑制松材线虫繁殖的效果,这可能与松材线虫和接种的腐生线虫种群间相互作用的时间长短有关。
生物防治是一种可持续性的防治方法,不仅可以在一定程度上替代化学农药,而且无残毒,对环境没有污染。除植物寄生线虫外[23],线虫中有许多种类是农林生态系统中的有效生物防治种[24-26]。本研究发现在环境温度较低(15 ℃)时,虫株JHS2在高接种量下可发挥对松材线虫的抑制作用,而虫株JSH4在低接种量下才可明显抑制松材线虫的种群数量,表明这两种腐生线虫发挥作用的有效接种量不同。Aalten[27]和Smitley等[28]分别使用小卷蛾斯氏线虫(Steinernemacarpocapsae)和嗜菌异小杆线虫(Heterorhabditisbacteriophora)抑制植物寄生线虫生长繁殖的相关研究表明,线虫种类不同,有效作用的施用量存在差异。温度是影响线虫生长发育的重要因子,低温可能限制了腐生线虫发挥竞争作用。史丽娜[20]使用腐生线虫培养基接种腐生线虫并置于不同温度下进行培养,结果表明在15 ℃时,腐生线虫能够正常进行生长繁殖,最适宜生长温度范围为20~25 ℃。本研究中在适生温度 25 ℃的培养后期(30 d时),仅高接种量的虫株JHS4发挥了抑制效果,表明有效的抑制作用是由接种量和时间相互作用共同决定的。这种相互作用与PÉREZ等[29-30]的研究结果相同。但虫株JHS4的抑制效果是否随着接种量的增加或时间的延长而提高还需要进一步的研究。
松材线虫传播到一个新的区域,要能成功定殖、建群和扩展,必须具备感病的寄主植物、携带线虫的媒介昆虫和适宜的环境条件[31-32]。天牛是松材线虫病的传播媒介,通过抑制媒介昆虫携带线虫的数量能够在一定程度上减少松材线虫病的发生。蒙海勤等[33]发现木腐真菌可阻断松材线虫向蛹室聚集。本研究结果表明腐生线虫能减少天牛及其蛹室周围的松材线虫数量,这可能与疫木内松材线虫整体数量减少直接相关。综上所述,松材线虫病疫木中存在某些腐生线虫种群能够加快松材线虫种群消亡的速度,将其应用于松材线虫病的疫木除治具有广阔前景,值得进一步深入开发研究。
