基于SRAP分子标记构建云南旱冬瓜核心种质
2024-03-13邹广权王晓丽曹现富张新洛曹子林
邹广权,王晓丽,曹现富,李 艳,张新洛, 曹子林
(1.西南林业大学西南山地森林资源保育与利用教育部重点实验室,昆明 650224; 2.西南林业大学生态与环境学院,昆明 650224)
【研究意义】旱冬瓜(Alnusnepalensis)系桦木科桤木属落叶乔木,是典型的喜马拉雅植物区系树种,在中国主要分布于西南地区。作为云南重要的乡土树种,旱冬瓜生长迅速,适应性强,干型优良,用途广泛[1]。收集和获取优良的种质资源是发展高质人工用材林的基础,云南省目前已完成收集云南省16个州(市)的64个旱冬瓜种源,并建立旱冬瓜种质资源库,为旱冬瓜林木资源的高效利用奠定了基础。在云南省玉溪市玉白顶林场旱冬瓜林木种质资源库中,优树收集区收集了来自云南省内43 个不同地区的优树,从优树的命名来看,多个优树来自于一个乡(镇)的情况较普遍,可能不同的优树间存在遗传关系较近的情况。核心种质库是种质资源中的优异性状集合群,开展构建旱冬瓜核心种质的研究工作,对了解旱冬瓜种质库中优树的遗传情况、降低优树间的遗传重复、节约种质资源库保存成本及后期利用具有重要作用。【前人研究进展】国内外关于核心种质的研究广泛,构建核心种质最佳的策略与方法也各有异同,研究对象主要集中在生长周期较短的食用作物上,如番茄(Solanumlycopersicum)、山药(Dioscoreapolystachya)、苦瓜(Momordicacharantia)等[2-4],对生长周期相对较长的木本植物主要集中在经济林木,如荔枝(Litchichinensis)、板栗(Castaneamollissima)、核桃(Juglansregia)等[5-7],随着林木培育的发展,在灰楸(Catalpafargesii)、杉木(Cunninghamialanceolata)、云南松(Pinusyunnanensis)等[8-10]用材林树种也有相关核心种质的报道。目前鲜见旱冬瓜核心种质构建的相关研究报道。SRAP分子标记在生物遗传多样性和重要性状标记等方面广泛运用。在云南松、马铃薯(Solanumtuberosum)、油茶(Camelliaoleifera)、何首乌(Fallopiamultiflora)等物种的遗传多样性和核心种质库构建策略的研究中,SRAP 分子标记技术已然是重要的手段[10-13],目前旱冬瓜遗传多样性的 SRAP 标记分析鲜见报道。【本研究切入点】借鉴王晓丽等[10]和Wang等[14]在材用云南松核心种质构建策略探讨中的研究方法,综合考虑旱冬瓜优树子代的生长状况和现实保存情况。【拟解决的关键问题】以旱冬瓜优树半同胞子代为试材,以SRAP 分子标记数据为源数据,从核心种质构建策略、核心种质评价和检测、核心种质的确认等方面探讨旱冬瓜核心种质的构建方法,为旱冬瓜优树的进一步评价、保存和利用提供依据。
1 材料与方法
1.1 供试材料
试验涉及的28株优树的84株半同胞子代(10年生)全部源于云南省玉溪市玉白顶林场旱冬瓜种质资源库(表1)。在每样株生长健壮的枝条上采集当年生叶片10 g,放入盛有硅胶的自封袋中保存,用于提取DNA和后续SRAP分子标记使用。

表1 试验所用旱冬瓜优树信息
试验用的TaqDNA聚合酶、10×TaqBuffer(含Mg2+)、dNTPs、2000 bp DNA Ladder Marker 由北京博迈德生物有限公司提供,SRAP-PCR反应引物序列(表2)由北京博迈德生物有限公司合成。
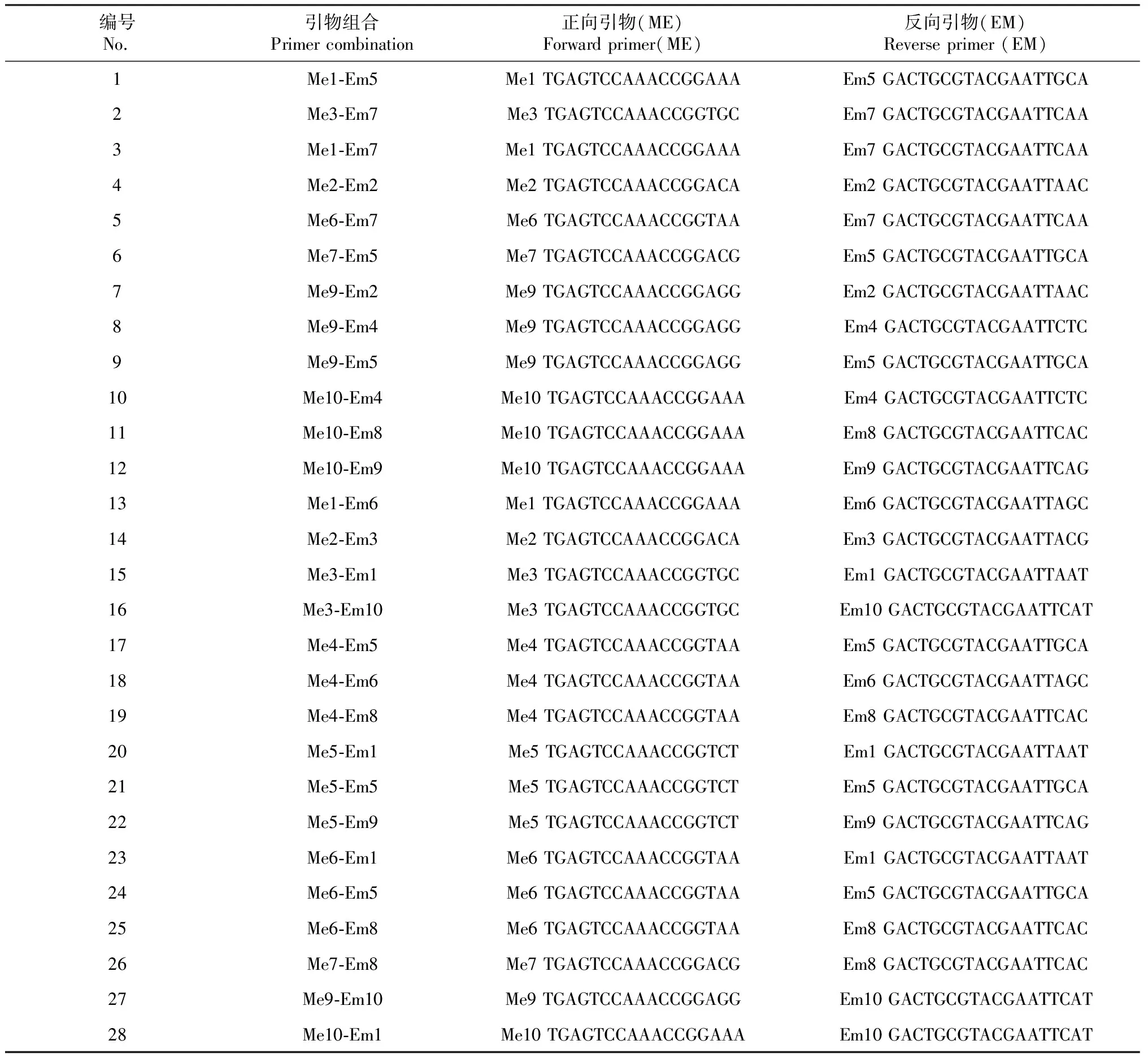
表2 SRAP -PCR扩增引物组合及其引物序列
1.2 试验方法
1.2.1 DNA提取及扩增 采用改良CTAB法提取旱冬瓜样株叶片DNA[15]。旱冬瓜SRAP-PCR反应体系和扩增程序参考前人的研究成果并加以改良[11-14],反应体系共20.0 μL,具体为DNA模板2.0 μL(50 ng/μL),上、下游引物(10 μmol/L)各1.5 μL,2×TaqPCR MasterMix 10.0 μL,ddH2O补齐至20 μL。扩展程序:94 ℃预变性5 min;94 ℃变性1 min,35 ℃退火1 min,72 ℃延伸1 min,共5个循环;94 ℃ 变性1 min,44 ℃退火1 min,72 ℃ 延伸1 min,共35个循环;最后72 ℃延伸7 min,4 ℃保存。PCR扩增产物经1%琼脂糖凝胶电泳中初检后,再采用8%聚丙烯酰胺凝胶电泳分离银染检测,拍照供分析。
1.2.2 核心种质的构建 利用各样株SRAP-PCR分子标记信息构建0-1矩阵数据,利用软件POPGENE32计算遗传相似矩阵,依据遗传相似矩阵,聚类软件为NTSYSpc2.10e,遗传距离采用Nei’s遗传距离,聚类方法采用不加权类平均法[10],取样方法采用改进的最小距离逐步取样法,设10%、15%、25%、35%、45%和55%共6个抽样比例,构建基于SRAP分子标记数据的种质子集。
1.2.3 核心种质评价 利用软件POPGENE32计算遗传多样性指数来评价种质子集[10,14],通过软件SPSS 23.0 和Excel 2022进行种质子集遗传多样性指数的t/F检验和相关系数分析,最后依据软件NTSYSpc2.10e的聚类结果来评价种质子集的代表性和实用性。
2 结果与分析
2.1 旱冬瓜原种质集SRAP-PCR扩增结果
采用28对SRAP引物对28个旱冬瓜优树的84株半同胞子代种质进行PCR扩增,共扩增出201个多态位点,用于构建旱冬瓜核心种质。如图 1所示,1~21号样品扩增充分分离、条带清晰明了,表明试验提取的DNA、筛选出SRAP 引物、采用的扩增体系、电泳体系和银染体系适合旱冬瓜种质研究。
2.2 旱冬瓜原种质集与不同抽样比例种质子集比较
2.2.1 遗传多样性比较 由表3可见,多态位点数、多态位点百分率、观测等位基因数和等位基因保留率在55%、45%、35%和25%抽样比例下与原种质集一致,而在15%和10%抽样比例下低于原种质集,说明当抽样比例过低时,原种质集的某些性状就会遗失,其余4个遗传多样性指数均大于原种质集,呈现出随抽样比例的降低而先增后减趋势,在25%抽样比例下,除有效等位基因数外,其余3个指标均达到最大值,说明基于优树子代构建的种质子集不仅能降低原种质集的遗传累赘,还能提高遗传多样性,且25%抽样比例优于其他抽样比例,其构建的种质子集在遗传多样性上更能代表原种质集。

1~21:引物组合Me2+ Em2对1~21号样株的扩增结果;M:标记。1-21:Amplification results of primer combination Me2+Em2 of sample strains 1-21;M:Marker.
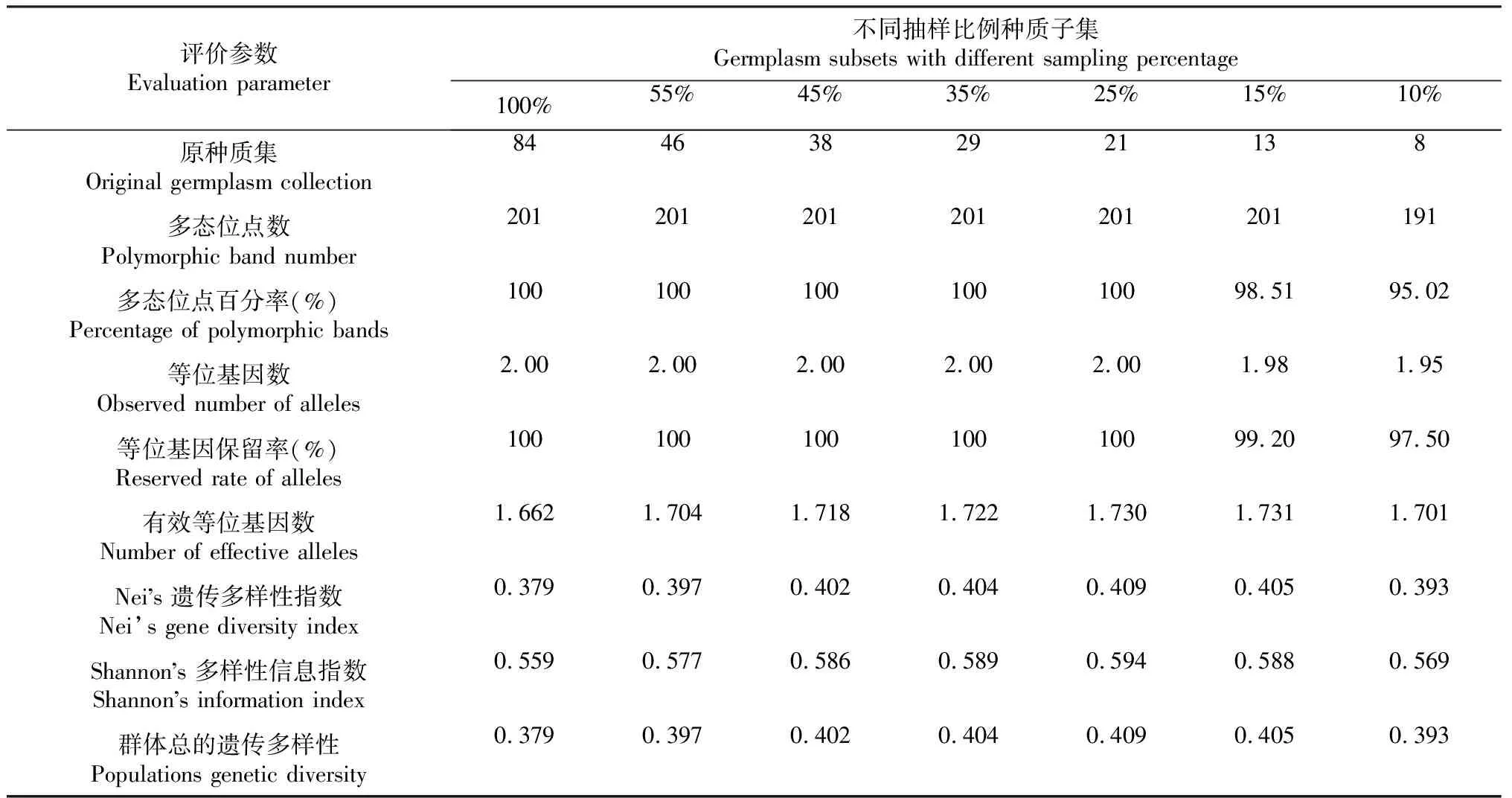
表3 旱冬瓜种质集遗传多样性指数
2.2.2 遗传多样性指标的均值t检验和方差F检验 由表4可见,在55%抽样比例构建的种质子集各指标的检验中,仅Shannon’s多样性信息指数F检验存在显著差异,其余指标的t/F检验均没有显著差异;在45%、35%、25%抽样比例构建的种质子集中,除等位基因数外,其余4个评价参数的方差均小于原种质集、均值皆大于原种质集,且在t/F检验中,这4个评价参数均与原种质集差异显著;在15%和10%种质子集的t/F检验中,除等位基因数与原种质集差异显著外,其余5个种质子集的等位基因数与原种质集无显著差异,就等位基因数而言,认为其余4个种质子集更有代表性,15%抽样比例种质子集的有效等位基因数在F检验存在显著差异;此外,随着抽样比例的降低,除等位基因数外的4个遗传评价参数的方差随差异比例降低而逐渐减小,在25%种质子集达到最小值后又随差异比例降低而逐渐增大,说明随着抽样比例的降低,各种质子集与原种质集的差异也先减后增,说明25%抽样比例构建的种质子集相较其他比例构建的种质子集,与原种质集差异最小。

表4 旱冬瓜种质集遗传多样性指标t/F检验
2.2.3 相关系数比较 如图2所示,种质子集与原种质集各遗传指数的均值相关系数接近或等于1,表现为强相关性,就均值而言,6个种质子集能代表原种质集;各遗传指数方差的相关系数只有10%抽样比例的种质子集与原种质集呈中度相关,从相关性来看,55%、45%、35%、25%及15%这5个种质子集均可代表原种质集。
综上,通过各种质集合的遗传多样性比较、t/F检验和相关系数比较,认为25%抽样比例种质子集能代表原种质集。
2.3 核心种质的确认
由表5可见,与原种质集的遗传距离相比,25%抽样比例种质子集的最大距离与原种质集相同,平均距离和最小距离大于原种质集,说明25%抽样比例种质子集保留了原种质的优异性状外,还有效地去除了原种质中的冗余材料,与遗传多样性指数分析结果一致。如图3所示,25%抽样比例种质子集遗传距离聚类图在0.55水平上分为两大类,聚类无有明显的地理趋势,不同种源的材料混杂的聚在一起,说明构建出的核心种质的代表性较好,遗传多样性较为丰富。认为25%抽样比例下的,以21个样株构建的种质子集能充分代表原种质集。
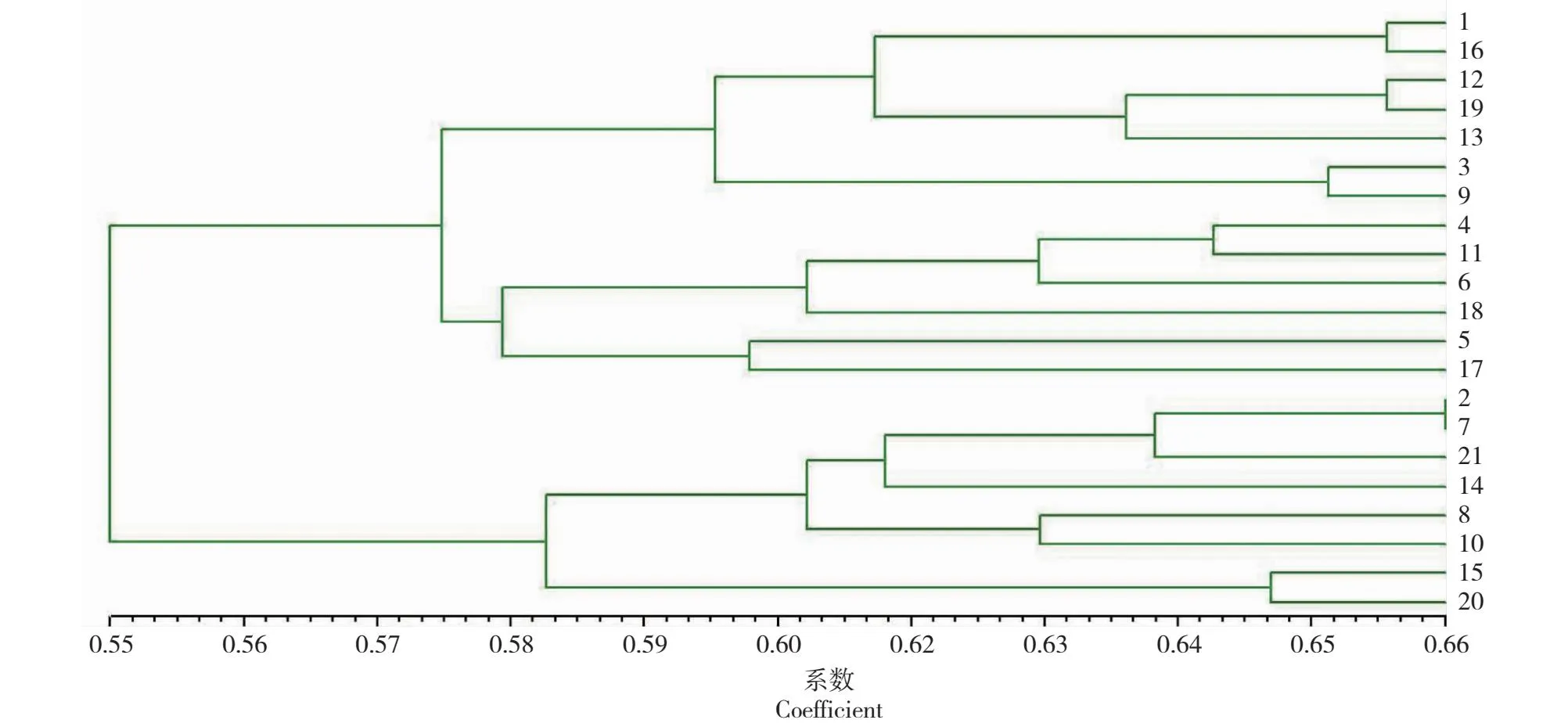
图3 25%抽样比例种质子集UPGMA聚类图Fig.3 UPGMA clustering diagram of germplasm subset with 25% sampling ratio
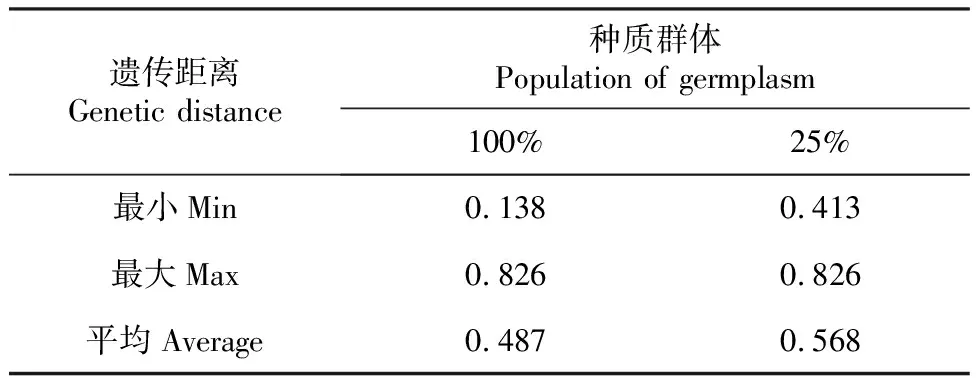
表5 遗传距离的比较
3 讨 论
本研究采用28对SRAP引物进行扩增,共扩增出201条多态性谱带,与前人研究结果相似[16-17]。核心种质库的大小取决于抽样比例的设置和原种质集大小。大量研究发现,5%~30%抽样比例构建的核心种质和种质资源保存库即可重复代表原种质群体[18],对不同的种质资源要考虑核心种质组成始终处于动态交流和调整之中,且随着种质资源的不断收集,还会出现新的变异类型,依据构建不同目的进行实用性检验是对核心种质有效性最直接和最具说服力的检验途径,王晓丽等[10]以30%的抽样比例构建的材用云南松种质保存库,可以提高原种质群体52.16%的遗传距离,刘遵春等[19]以新疆野苹果初级核心种质中的60份种质构建了包含45份的核心种质,其抽样比例高达70%,为原始种质的14%,保留了93%以上的农艺性状,郑福顺等[2]以15%抽样比例构建包含72份宁夏地区的番茄核心种质。本研究认为25%抽样比例构建的核心种质,符合前人研究的抽样比例规律,所构建的核心种质能充分保留原种质集遗传性质,提高种质间的遗传距离。
基于离散型的分子标记数据推测林木间的遗传距离和遗传多样性,采用的遗传距离和聚类法依林木种类而不同,最常见的是Nei’s遗传距离,而聚类方法多数采用不加权类平均法,构建核心种质的研究中也常见关于遗传距离、聚类方法、抽样方法及比例等构建策略的研究[10,19-20]。评价指数常见的有标记频率差异百分率、平均期望杂合度、多态位点百分率、Shannon-Weaver多样性指数、平均有效等位基因数和平均多态信息含量[10,19-21],常通过t/F检验分析种质子集的代表性。本研究依据前人研究的种质子集评价指数,结合均值t/F检验、相关系数分析、聚类分析等分析方法来分析种质子集对原种质集的代表性,结果表明,构建的核心种质尽可能保留等位基因数和遗传多样性,提高原种质集的遗传距离,减少冗余,这与前人的研究结果一致,其分析结果在后续基于SRAP分子标记数据构建林木核心种质的研究中具有一定的借鉴参考价值。但本研究的样品总数较少,因此在今后研究中还需补充并收集新的种质资源。
大量研究表明,生物的优良家系及优树的平均遗传增益和经济效益显著高于普通树种[22-23],林木种质资源库的建设对林木资源的保护和利用具有重要意义。但林木树体高大、生长周期长,且种质资源收集工作是不断动态变化的,数量会不断增加,导致其种质资源保存费力、费时,而在种质资源库的基础上构建核心种质是解决上述问题的一个有效途径。不同于以往林木种质构建核心种质研究中的大范围取样,种质资源库中的优树样株本身已经经过一次选优过程,构建核心种质更加便捷。曾宪君等[24]以欧洲黑杨基因库中的优树无性系子代,构建了欧洲黑杨优质核心种质;彭婵等[25]以湖北南方型美洲黑杨种质资源库中的优树无性系,构建了美洲黑杨优质核心种质;张良波[26]以光皮树种质资源库中的211株光皮树优树构建光皮树优树核心种质。本研究以旱冬瓜种质资源库中的优树子代(10年生)为试材,进一步探索构建林木核心种质的方法。研究表明,基于优树半同胞子代构建旱冬瓜核心种质的方法与前人研究结果类似,为优质旱冬瓜种质资源的长期保存和高效利用提供理论依据,也为其他林木核心种质构建提供参考。
4 结 论
在25%抽样比例下构建的旱冬瓜种质子集能充分代表原种质的遗传多样性,可作为该原种质的核心种质。利用上述核心种质构建策略,从84株样株组成的原种质中,筛选出21株样株组成旱冬瓜核心种质。
