嫁接对番茄植株根际微生物群落组成的影响
2024-03-13周信雁魏宇飞谢源源杨尚东庞师婵
朱 瑜,周信雁,魏宇飞,谢源源,韦 勋,杨尚东,庞师婵
(1.广西大学农学院/广西农产品安全重点实验室/植物科学国家级实验教学示范中心,南宁 530004;2.广西南亚热带农业科学研究所,广西 龙州 532415)
【研究意义】番茄是世界上消费量最大的蔬菜之一[1]。近年来,由于我国番茄主产区长期重茬栽培,连作障碍危害日趋严重,已成为阻碍番茄产业可持续发展的重要问题之一[2]。实际生产中,运用嫁接技术已显著提高了黄瓜、茄子、葡萄、苹果等作物的抗性、品质和产量等农艺性状,有效缓解了连作障碍[3]。同样,嫁接亦有助于减轻番茄连作障碍的危害,可增加番茄植株根系对养分的吸收、增强抗病性及提高番茄品质[4-5]。植物与土壤微生物之间存在密切关系,植物根系通过产生根系分泌物可以使土壤中微生物富集于根系周围[6],而根际微生物则可以通过多种代谢活动显著影响植物生长发育[7-9]。根际微生物主要包括细菌和真菌,其丰度占比可达60%~80%,两者之间不断进行相互作用,通过养分和生态位的竞争增加自身的种群优势[10]。其中,有益微生物可以通过自身代谢活动影响植物的生长,如调控土壤营养元素的循环、分泌激素类化合物和抑制病原菌侵害等;而有害微生物则会侵染植物,阻碍植物生长或导致其死亡[11]。因此,研究番茄实生和嫁接植株根际土壤微生物群落组成,揭示嫁接对番茄根际土壤微生物的影响,能为挖掘对番茄生长有益的微生物种类提供思路,对促进番茄产业可持续发展有重要意义。【前人研究进展】嫁接可诱导植物根际土壤微生物促进土壤碳、氮、磷元素的循环,进而提升植物对营养物质的吸收利用能力[11-13]。刘斯晗等[14]发现,番茄嫁接可以促使其根系产生邻苯二甲酸二甲酯(DP)和2,6-二叔丁基对甲苯酚(BHT)2种抗病性化合物,DP和BHT可影响番茄根际微生物的形成,从而显著提高番茄对青枯病的抵抗能力;覃仁柳等[15]发现,番茄嫁接显著提高根系内生微生物多样性和有益微生物数量,进而增强植株对环境的适应性;茄子[16]和西瓜[17]嫁接处理也可以显著增加其有益根际微生物数量,有效降低植株黄萎病和枯萎病发病率。嫁接不同种类砧木可以不同程度改变植株根系活力和根际、叶际微生物的数量和种类,可以提高植物对土壤病原体或不利环境的抵抗力[18-19]。根际土壤微生物通过生命活动具有分解土壤中有机质、产生植物激素类化合物、降解根系分泌物、塑造菌根等作用,从而可以改变土壤性质和根系结构,为植株生长创造更加有利的条件[20]。【本研究切入点】虽然前人对嫁接提高番茄植株抗性已开展了相关研究,但嫁接不同砧木对番茄植株根际微生物组成的影响仍鲜见报道。【拟解决的关键问题】本文选用不同砧木,基于高通量测序技术系统分析嫁接对番茄植株根际土壤微生物(细菌和真菌)群落组成的影响,进一步阐明嫁接提高番茄植株抗性的作用方式,为构建生态防控番茄土传病害的栽培技术体系提供参考。
1 材料与方法
1.1 试验材料
砧木为新砧1号和宝砧1号,其具有适应性广、抗病性强、根系生长发达等特性。接穗选用桃太郎番茄品种,其生长势强、产量高且营养品质优良。砧木和接穗均购于农友种苗(中国)有限公司。
1.2 试验方法
1.2.1 试验设计与样品采集 2020年8月至12月在广西大学农学院蔬菜生产基地(108°17′25″ E,22°51′02″ N)内进行盆栽试验,定植盆高50 cm、直径40 cm。试验点土壤类型为赤红壤土,pH 5.71,有机质21.2 g/kg,全氮1.05 g/kg,全磷0.49 g/kg,全钾7.4 g/kg,碱解氮131.7 mg/kg。设5个处理:新砧1号实生植株(简称:新砧)、新砧1号×桃太郎嫁接植株(简称:桃太郎/新砧)、宝砧1号实生植株(简称:宝砧)、宝砧1号×桃太郎嫁接植株(简称:桃太郎/宝砧)、桃太郎实生植株(简称:桃太郎)。当番茄幼苗长至3叶1心时采用插接法进行嫁接,植株成活后进行定植。每个试验处理设置3组重复,每组重复种植20株。番茄生长管理按常规方法进行,每间隔4 d浇透一次水。
番茄植株定植80 d后,采集根际土壤样品。具体方法如下:以番茄植株为中心,直径20 cm左右,挖松植株周围土壤,然后手握植株茎底部,拔取整个植株,采用“S”形采样方法,3株混合作为一个样品,每个处理3次重复;同时采用抖根法[21]收集附着在根系表面的土壤置于样品袋中,放入冰盒带回实验室,于-80 ℃冰箱中保存备用。
1.2.2 总DNA提取、PCR扩增和序列测定分析 总DNA提取、PCR扩增和序列分析由上海美吉生物医药科技有限公司完成。具体操作步骤如下:总DNA抽提根据FastDNA Spin Kit for Soil 试剂盒(MP Biomedicals,美国)说明进行,DNA浓度和纯度使用NanoDrop2000分光光度计(Thermo Fisher Scientific,美国)检测,利用1%琼脂糖凝胶电泳检测DNA提取质量,其中细菌使用338F (5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3')、真菌使用ITS1F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和ITS2F(5'-GCTGCGTTCTTCATCATCGATFC-3')为引物对16SrRNA基因和ITS基因的V3~V4可变区进行PCR扩增,最后进行PCR产物鉴定、纯化及定量分析。llumina MiSeq测序:将同一样本的PCR产物混合后使用2%琼脂糖凝胶回收PCR产物,利用AxyPrep DNA Gel Extraction Kit (Axygen Biosciences,美国)进行回收产物纯化,2%琼脂糖凝胶电泳检测,并用Quantus TM Fluorometer (Promega Corporation,美国)对回收产物进行定量检测。使用NEXTflex Rapid DNA-Seq Kit进行建库,利用上海美吉生物医药科技有限公司MiSeq PE300平台进行测序。
1.3 统计分析
采用Excel 2019进行数据统计整理;利用IBM SPSS Statistics 26,采用Duncan’s新复极差法进行显著性分析。使用Mothur 1.30.2计算微生物群落的α多样性;选择相似度97%的操作分类单元(OTU)表进行微生物群落组成和维恩图分析,并使用R语言(3.3.1版本)工具进行统计、绘图和非度量多维尺度分析(NMDS);根据分类组成的不同分组条件,使用LEfSe进行线性判别分析(LDA),以识别各处理显著差异的物种。利用上海美吉生物医药科技有限公司的I-sanger云数据平台进行在线数据分析。
2 结果与分析
2.1 嫁接对番茄植株根际土壤细菌群落在不同分类水平上OTU数量的影响
由表1可知,番茄实生和嫁接植株根际土壤细菌群落OTU可归类为37门、97纲、267目、463科、905属、1754种。与新砧、宝砧、桃太郎处理植株相比,桃太郎/新砧和桃太郎/宝砧处理植株根际土壤细菌群落在不同分类水平上OTU数量发生明显变化。其中,与新砧和桃太郎处理植株相比,桃太郎/新砧处理植株根际土壤细菌群落在不同分类水平上OTU数量整体减少;与宝砧和桃太郎处理植株相比,桃太郎/宝砧处理植株根际土壤细菌群落在门、纲和科分类水平上OTU数量增加,在属分类水平上OTU数量下降。与砧木和接穗的实生植株相比,嫁接植株根际土壤细菌群落组成发生了变化,而且变化程度受砧木种类影响。
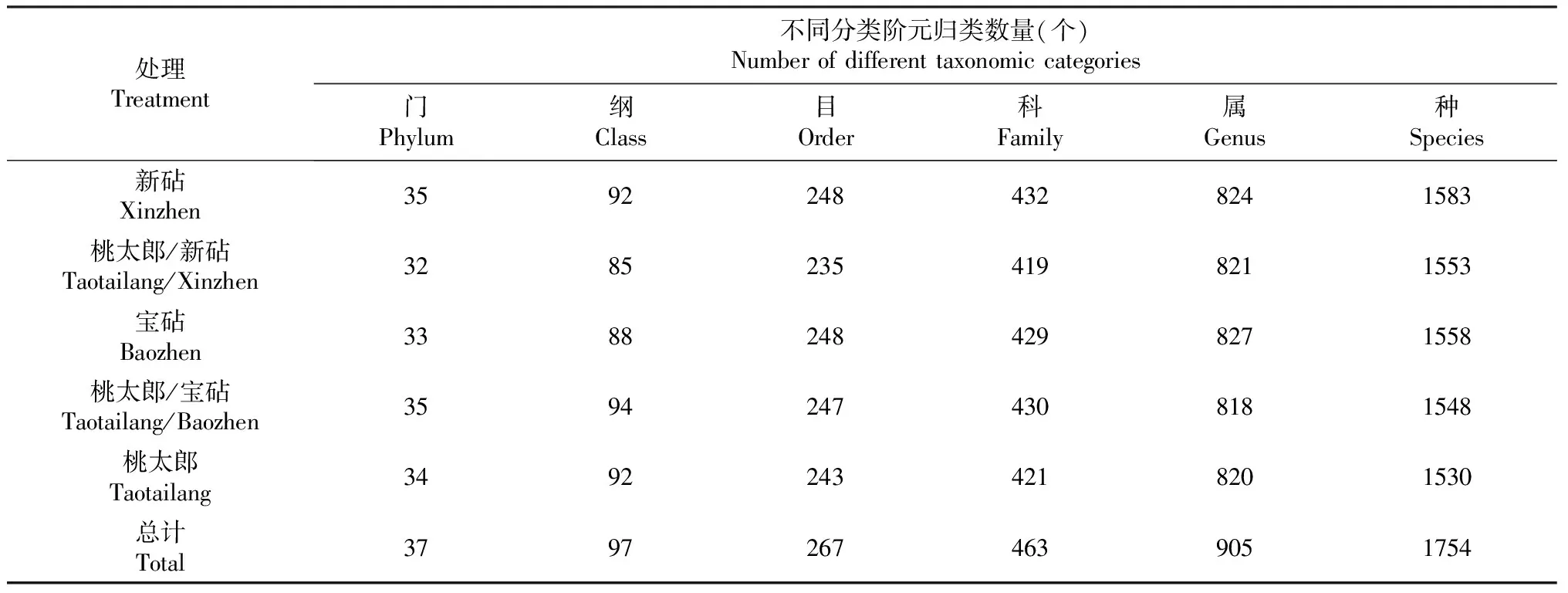
表1 嫁接对番茄植株根际土壤细菌在不同分类水平上OTU数量的影响
2.2 嫁接对番茄植株根际土壤细菌Alpha多样性的影响
由表2可知,各处理番茄植株根际土壤细菌群落测序结果的覆盖度均在96.0%以上,表明测序结果可以代表样本中微生物的真实情况。嫁接植株根际土壤细菌的多样性指数(Shannon指数)和丰富度指数(ACE和Chao l指数)与其对应砧木和接穗实生植株相比均差异不显著(P>0.05,下同),仅有新砧和宝砧2种实生砧木植株间根际土壤细菌的多样性指数差异显著(P<0.05,下同)。不同砧木植株根际土壤细菌群落多样性与丰富度存在显著差异,这可能亦是不同种类砧木之间抗性存在差异的重要原因之一。
2.3 嫁接对番茄植株根际土壤细菌群落组成影响的NMDS分析
由NMDS分析结果(图1)可知,在OTU水平上,桃太郎/新砧处理植株与新砧和桃太郎处理植株间在根际土壤细菌群落结构上差异显著,桃太郎/宝砧处理植株与宝砧和桃太郎处理植株在根际土壤细菌群落结构上差异显著,说明嫁接改变了番茄植株根际土壤的细菌群落结构。
2.4 嫁接对番茄植株根际土壤细菌群落组成的影响
2.4.1 门分类水平优势菌群分析 由图2可知,变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、绿弯菌门(Chloroflexi)、厚壁菌门(Firmicutes)、髌骨细菌门(Patescibacteria)、拟杆菌门(Bacteroidetes)、芽单胞菌门(Gemmatimonadetes)和酸杆菌门(Acidobacteria)细菌是番茄嫁接和实生植株根际土壤的共有优势细菌门类(丰度占比大于1.00%,下同)。
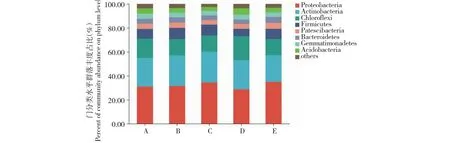
图2 番茄实生和嫁接植株根际土壤细菌的门分类水平组成Fig.2 Compositions of rhizosphere soil bacteria in tomato plants and grafted plants at phylum level
其中,在桃太郎/新砧处理植株根际土壤中,放线菌门和厚壁菌门细菌丰度占比均高于新砧和桃太郎处理植株;但绿弯菌门、髌骨细菌门、酸杆菌门、芽单胞菌门细菌丰度占比则低于桃太郎和新砧处理植株。在桃太郎/宝砧处理植株中,绿弯菌门、酸杆菌门、芽单胞菌门细菌丰度占比均高于宝砧和桃太郎处理植株;而桃太郎/宝砧处理植株根系中变形菌门、拟杆菌门和厚壁菌门细菌丰度占比分别为28.89%、3.59%、6.10%,均低于宝砧和桃太郎处理植株。综上所述,嫁接改变了番茄植株根际土壤细菌群落门分类水平组成的丰度占比,而且改变程度与砧木种类有关。
2.4.2 属分类水平优势菌群分析 由图3可知,番茄嫁接和实生植株根际土壤中,共有的优势细菌属分类数量为16个。其中,与新砧和桃太郎处理植株相比,桃太郎/新砧处理植株根际土壤中,鞘氨醇单胞菌属(Sphingomonas)、芽孢杆菌属(Bacillus)、链霉菌属(Streptomyces)、unclassified_f__Micrococcaceae、norank_f__norank_o__Gaiellales细菌属丰度占比提升;但norank_f__Gemmatimonadaceae、norank_f__Roseiflexaceae属细菌丰度占比下降;与桃太郎处理植株相比,桃太郎/新砧处理植株根际土壤中增加特有优势菌属栖大理石菌属(Marmoricola)。与宝砧和桃太郎处理植株相比,桃太郎/宝砧处理植株根际土壤中,norank_f__norank_o__norank_c__Subgroup_6、类诺卡氏菌、norank_f__norank_o__norank_c__Gitt-GS-136、norank_f__norank_o__norank_c__KD4-96、norank_f__JG30-KF-CM45属细菌丰度占比增加,同时拥有特有优势菌属norank_f__norank_o__norank_c__JG30-KF-CM66;但鞘氨醇单胞菌属、芽孢杆菌属、德沃斯氏菌属(Devosia)、norank_f__norank_o__Gaiellales、unclassified_f__Micrococcaceae细菌属丰度占比下降。
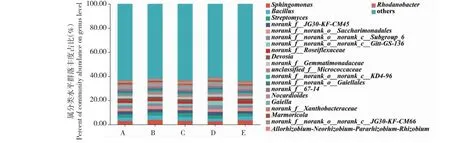
图3 番茄实生和嫁接植株根际土壤细菌的属分类水平组成Fig.3 Compositions of rhizosphere soil bacteria in tomato plants and grafted plants at genus level
2.5 嫁接和实生番茄植株根际土壤细菌LEfSe分析
如图4所示,通过LDA方法分析多级物种的差异,以期发现不同处理中差异显著物种,结合LDA效应值与P值比较各处理中细菌物种丰度对差异效果影响大小,筛选出差异物种(LDA score ≥ 2.0,P<0.05)。在新砧处理中发生显著变化的细菌差异物种有4个:Pseudonocardiaceae、Pseudonocardiales、Amycolatopsisg、Wandonia;在桃太郎/新砧处理中发生显著变化的细菌差异物种有9个:norank_f__Chitinophagaceae、Elsterales、Flavitalea、Gemmatirosa、Criblamydiaceae、norank_f__Criblamydiaceae、Longimycelium、unclassified_o__Elsterales、unclassified_o__Elsterales;在宝砧处理中发生显著变化的细菌差异物种有5个:Geodermatophilaceae、Iamiaceae、Iamia、unclassified_f__Rhodobacteraceae、Anaeromyxobacter;在桃太郎/宝砧处理中发生显著变化的细菌差异物种有6个:Solirubrobacteraceae、unclassified_f__Geodermatophilaceae、unclassified_f__Solirubrob acteraceae、JCM_18997、UTBCD1、Deinococcaceae;在桃太郎处理中发生显著变化的细菌差异物种有13个:Chitinophagales、Chitinophagaceae、Flavisolibacter、Hymenobacteraceae、Peredibacter、norank_f__norank_o__Chitinophagales、Bacteriovoracaceae、norank_o__Chitinophagales、norank_o__norank_c__Elusimicrobia、norank_c__Elusimicrobia、norank_f__Vermiphilaceae、Vermiphilaceae、norank_f__norank_o__norank_c__Elusimicrobia。表明,番茄嫁接处理可显著影响植株根际土壤细菌群落的组成。
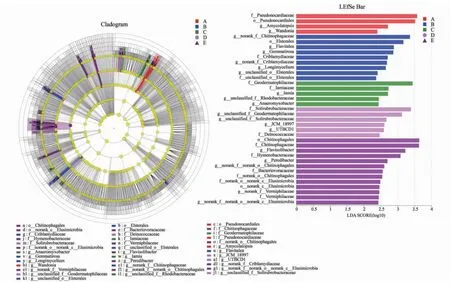
圆圈表示从门到属的系统发育水平,每个圆的直径与该组的丰度成正比;不同的前缀表示不同的级别(p:门; c:类;o:目; f:科; g:属,下同)。Circles indicated phylogenetic levels from phylum to genus, the diameter of each circle was proportional to the abundance of the group; Different prefixes indicated different levels different levels (p:phylum; c:class,o:Order; f:Family; g:Genus. The same as below).
2.6 嫁接和实生番茄植株根际土壤细菌Venn图分析
由图5可知,新砧、桃太郎和桃太郎/新砧处理植株根际土壤中分别获得4291、4096、4026个OTUs;其中,新砧、桃太郎和桃太郎/新砧处理植株根际土壤中特有OTU数量分别为253、174和156个。宝砧、桃太郎和桃太郎/宝砧处理植株根际土壤中分别获得4006、4096、4101个OTUs;其中,宝砧、桃太郎和桃太郎/宝砧处理植株根际土壤中特有的OTU数量分别为188、217、229个OTUs。分析可知,嫁接能改变番茄植株根际土壤中OTU数量,嫁接植株根际土壤中OTU数量比实生植株增加,且不同砧木作用不同,这是嫁接增强植株抗性的重要原因之一。
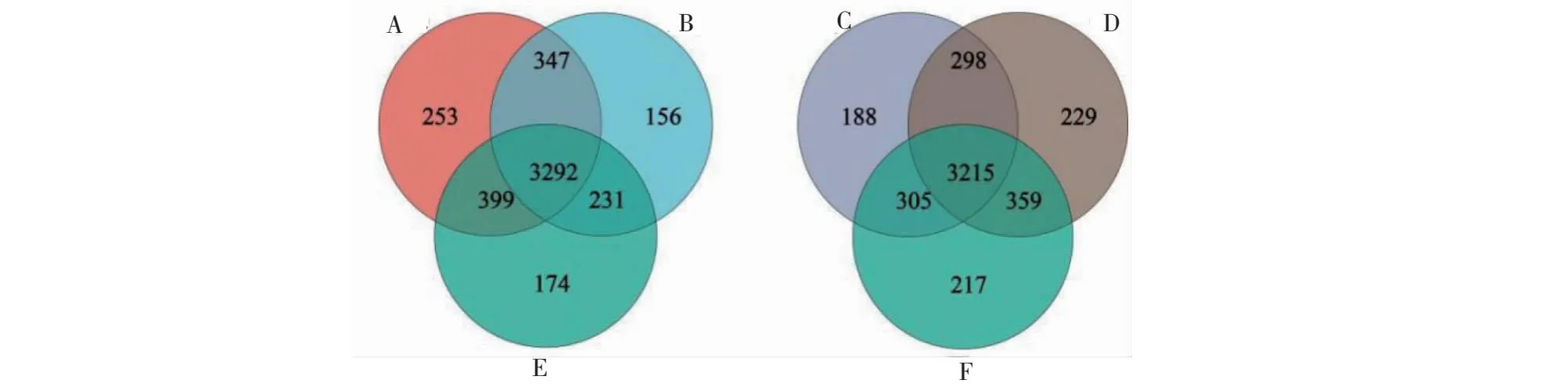
图5 番茄根际土壤细菌OTU分布的Venn图Fig.5 Venn diagram of OTU distribution of rhizosphere soil bacteria in tomatoes
2.7 嫁接对番茄植株根际土壤真菌群落在不同分类水平上OTU数量的影响
由表3可知,番茄实生和嫁接植株根际土壤真菌群落OTU可归类到12门、30纲、62目、127科、203属、304种。与新砧、宝砧、桃太郎处理植株相比,桃太郎/新砧和桃太郎/宝砧处理植株根际土壤在不同分类水平上的真菌OTU数量发生明显变化。其中,与新砧和桃太郎处理植株相比,桃太郎/新砧处理植株根际土壤真菌在科、属和种分类水平上OTU数量整体增加,而在纲分类水平上OTU数量减少;与宝砧和桃太郎处理植株相比,桃太郎/宝砧处理植株根际土壤在目、科、属和种分类水平上OTU数量整体减少。表明,与砧木或接穗实生植株相比,嫁接同样改变了植株根际土壤真菌群落组成,变化程度受砧木种类影响。
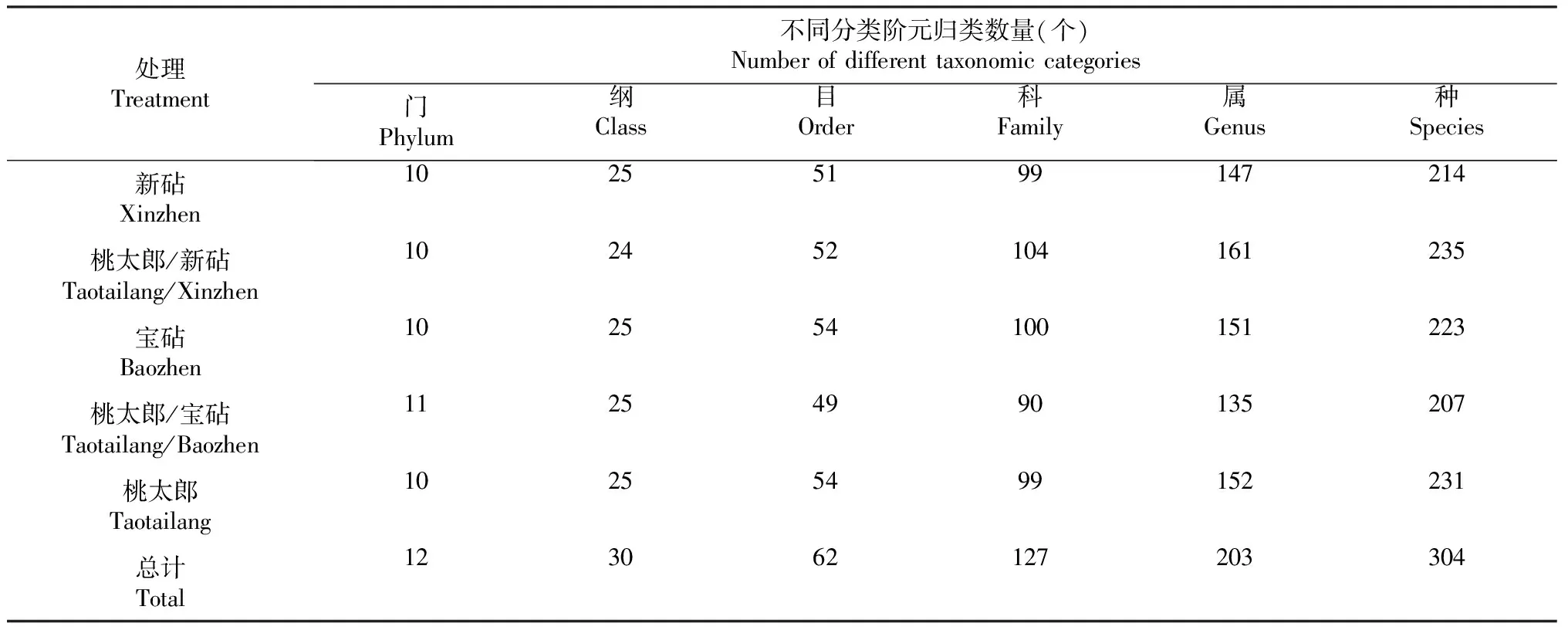
表3 嫁接对番茄植株根际土壤真菌群落在不同分类水平上OTU数量的影响
2.8 嫁接对番茄植株根际土壤真菌Alpha多样性的影响
由表4可知,番茄各处理植株根系的覆盖率均在99.00%以上,表明测序结果可以代表样本中真菌微生物的真实情况。番茄实生和嫁接植株根际土壤之间真菌的Shannon、Chaol、Heip和ACE指数均无显著差异,表明与砧木或接穗实生植株相比,嫁接对番茄植株根际土壤真菌的多样性和丰富度无显著影响。
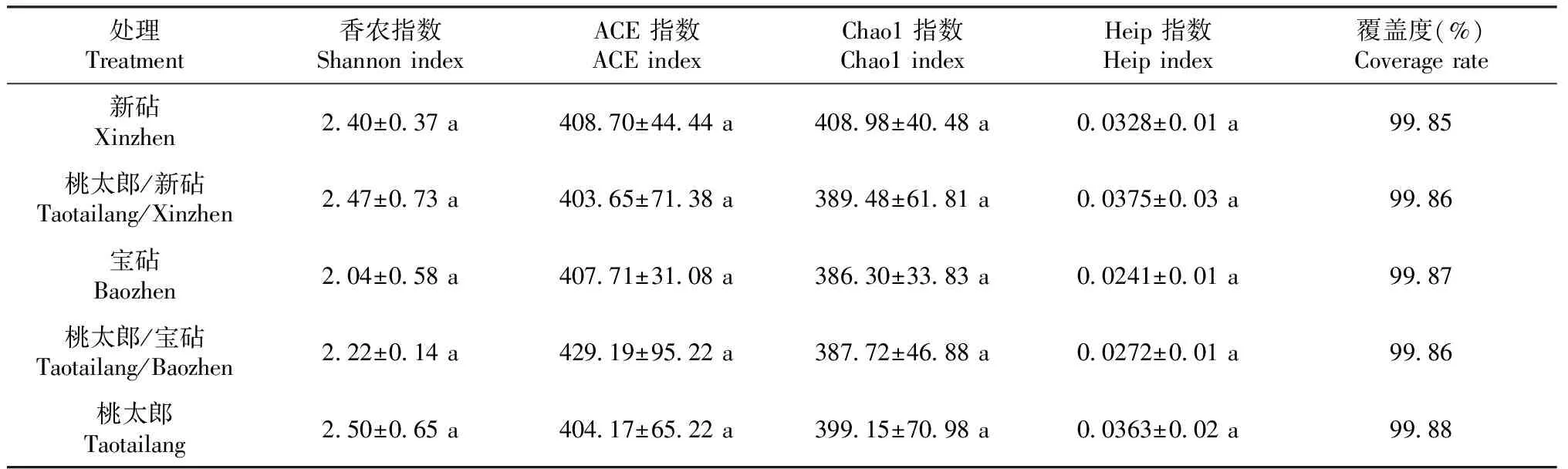
表4 番茄不同处理根际土壤真菌的多样性指数和丰富度指数比较
2.9 嫁接对番茄植株根际土壤真菌群落组成影响的NMDS分析
由NMDS分析结果(图6)可知,在OTU水平上,桃太郎/新砧处理植株根际土壤真菌群落与新砧和桃太郎处理植株根际土壤真菌群落有差异,但差异不显著;而桃太郎/宝砧处理植株根际土壤真菌群落与宝砧和桃太郎处理植株根际土壤真菌群落存在显著差异。表明嫁接同样改变了植株根际土壤的真菌群落结构,且变化程度与砧木种类有关。
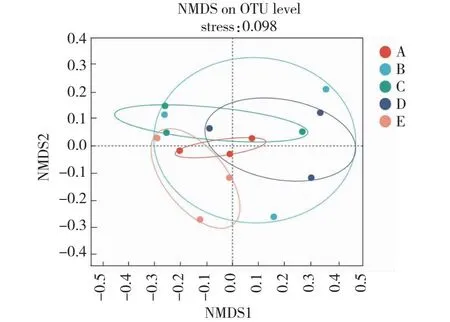
图6 番茄实生和嫁接植株根际土壤真菌的群落组成Fig.6 Composition of rhizosphere soil fungi communities between the roots of live and grafted tomato plants
2.10 嫁接对番茄植株根际土壤真菌群落组成的影响
2.10.1 门分类水平优势菌群分析 由图7可知,子囊菌门(Ascomycota)、壶菌门(Chytridiomycota)、unclassified_k__Fungi门真菌是嫁接和实生番茄植株根际土壤中共有的优势真菌门类。与番茄实生植株相比,嫁接植株根际土壤中壶菌门真菌丰度占比提升,而子囊菌门真菌丰度占比下降。
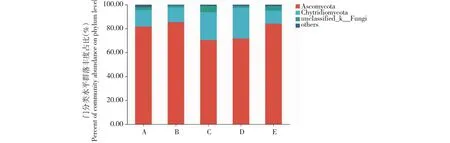
图7 番茄实生和嫁接植株根际土壤真菌门的分类水平组成Fig.7 Compositions of rhizosphere soil fungi in tomato plants and grafted plants at phylum level
2.10.2 属分类水平优势菌群分析 由图8可知,毛壳菌属(unclassified_f__Chaetomiaceae)、根囊壶菌属(Rhizophlyctis)、unclassified_c__Sordariomycetes、unclassified_k__Fungi、青霉属(Penicillium)是番茄嫁接植株和实生植株根际土壤中共有的5个优势真菌属。与桃太郎和新砧处理植株相比,桃太郎/新砧处理植株根际土壤中,根囊壶菌属、unclassified_k__Fungi、分枝孢子霉属(Cladosporium)真菌的丰度占比增加,而毛壳菌属真菌丰度占比下降,此外,桃太郎/新砧嫁接植株根际土壤缺失木霉属(Trichoderma)、曲霉菌属(Aspergillus)、unclassified_o__Eurotiales、unclassified_o__Sordariales等菌属,但与新砧处理植株相比,增加了镰刀菌属(Fusarium)。与桃太郎和宝砧处理植株相比,桃太郎/宝砧处理植株根际土壤中,根囊壶菌属、青霉属丰度占比增加,而unclassified_k__Fungi、unclassified_c__Sordariomycetes属真菌丰度占比下降;而桃太郎/宝砧处理植株根际土壤缺失分枝孢子霉属、木霉属、曲霉菌属、镰刀菌属、unclassified_o__Eurotiales、蛛网霉属(Arachniotus)等菌属。
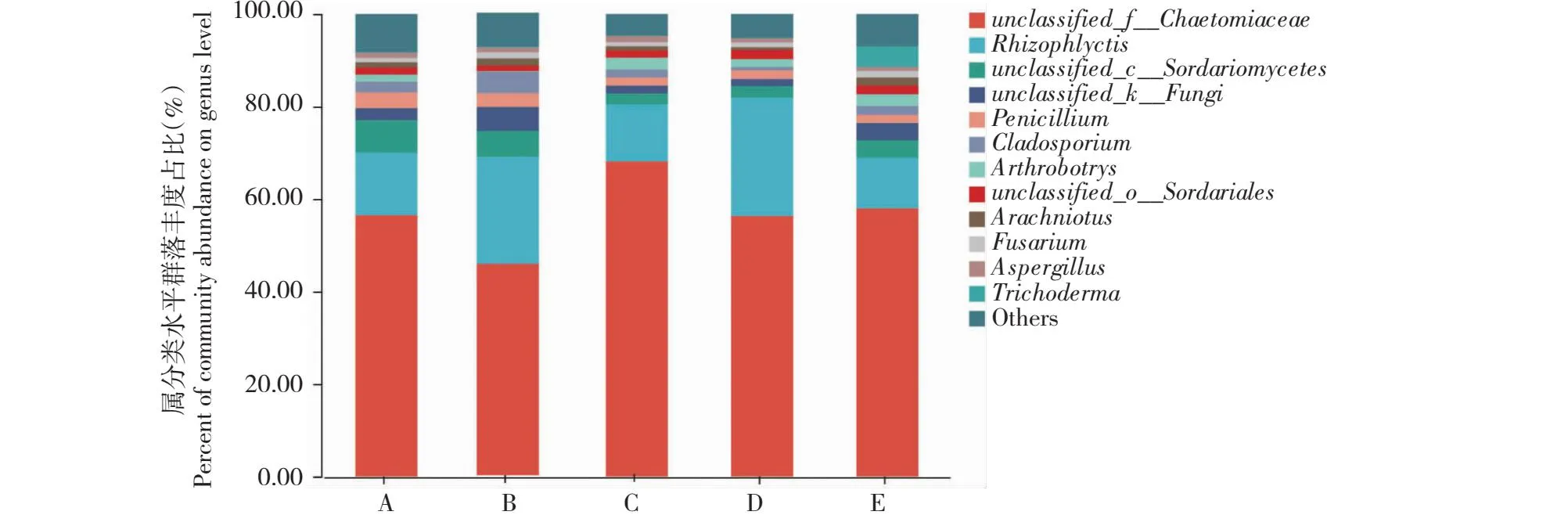
图8 番茄实生和嫁接植株根际土壤真菌的属分类水平组成Fig.8 Compositions of rhizosphere soil fungi in tomato plants and grafted plants at genus level
2.11 番茄实生和嫁接植株根际土壤真菌LEfSe分析
如图9所示,通过LDA方法分析多级物种的差异,以期发现不同处理中差异显著物种,结合LDA效应值与P值比较各处理中真菌物种丰度对差异效果影响大小,筛选出差异物种(LDA score≥3.0,P<0.05)。其中,在真菌分类水平上只有新砧、桃太郎/新砧和桃太郎处理植株根际土壤中分别拥有1、6、2种显著变化的真菌种类,而宝砧和桃太郎/宝砧处理则无显著变化真菌种类。在新砧处理植株根际土壤中发生显著变化的真菌差异物种为:Pleosporales;在桃太郎/新砧处理植株根际土壤中发生显著变化的真菌差异物种包括:Dothideomycetes、Cladosporiaceae、Capnodiales、Cladosporium、Pleosporaceae、Curvularia;在桃太郎处理植株根际土壤中发生显著变化的真菌差异物种包括:Cortinariaceae、Gymnopi-lus。结果表明,不同种类砧木与接穗嫁接处理对植株根际土壤真菌群落的影响有差异。
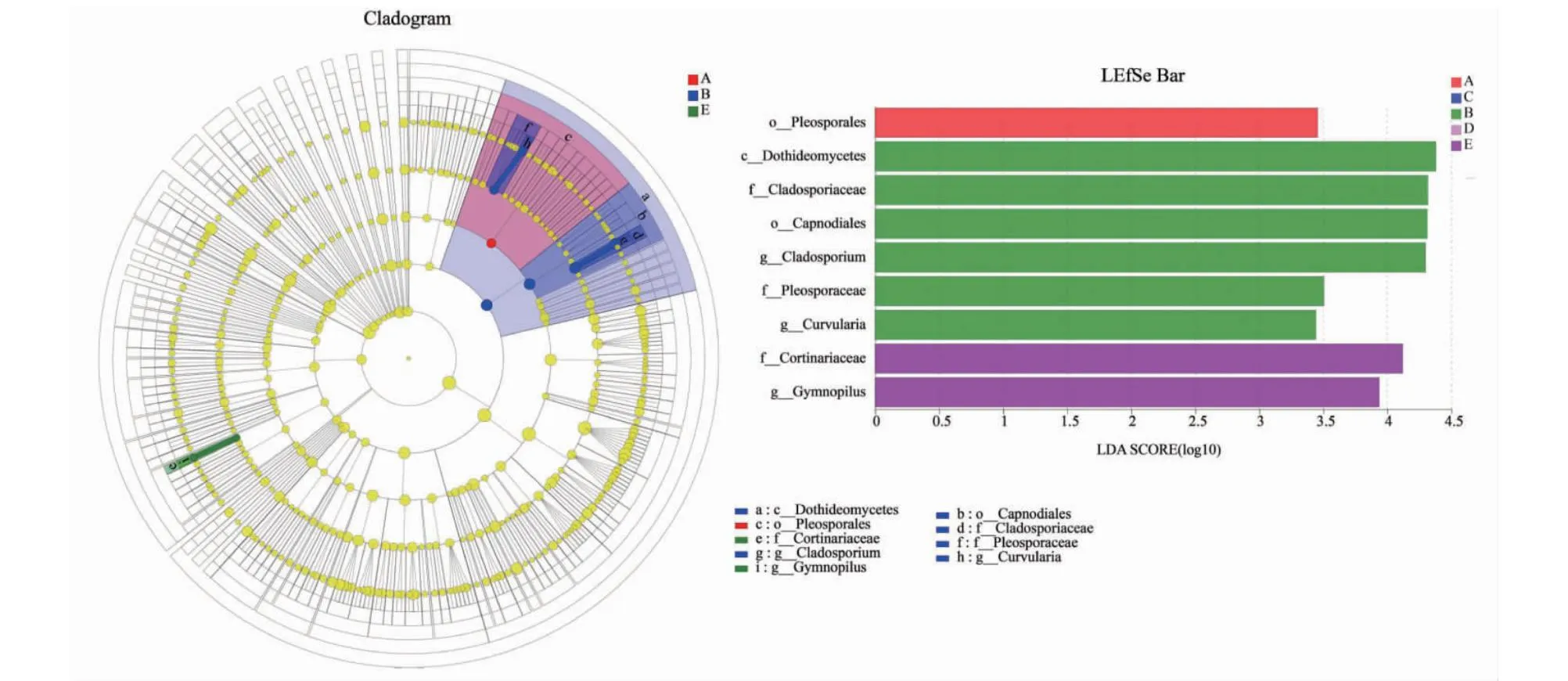
图9 番茄实生和嫁接植株根际土壤真菌的LEfSe分析(LDA阈值 ≥ 3.0)Fig.9 LEfSe analysis of rhizosphere soil fungi in tomato plants and grafted plants(LDA score ≥ 3.0)
2.12 番茄嫁接和实生植株根际土壤真菌Venn图分析
由图10可知,新砧、桃太郎和桃太郎/新砧处理植株根际土壤中分别共获得509、549、510个OTUs;其中,新砧、桃太郎和桃太郎/新砧处理植株根际土壤中特有的OTU数量分别为54、78和59个。宝砧、桃太郎和桃太郎/宝砧处理植株根际土壤中共获得491、549、495个OTUs;其中,宝砧、桃太郎和桃太郎/宝砧植株根际土壤中特有的OTU数量分别为54、75、44个。嫁接改变番茄植株根际土壤OTU数量,嫁接植株根际土壤OTU数量与实生植株相比有所减少,且不同砧木作用效果不同,这是嫁接增强植株抗性的重要原因之一。
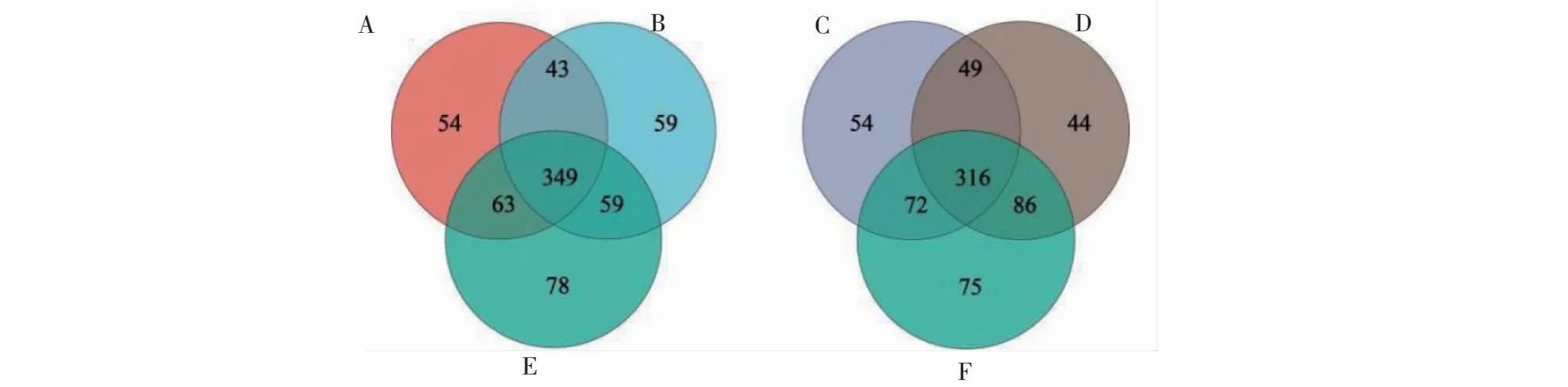
图10 番茄根际土壤真菌OTU分类水平的Venn图Fig.10 Venn diagram of OTU distribution of rhizosphere soil fungi in tomatoes
3 讨 论
植物根际土壤中拥有大量微生物,被称为植物的第二基因库[22],且每种微生物具有不同功能,进而可从多方面影响植物的生长和发育[23]。植物根际微生物群落组成受多种因素的影响[24],其中植物主要通过根系代谢活动产生分泌物影响根际微生物群落的组成,如:糖类、氨基酸、有机酸、植物激素、水解酶、酚类等物质;而植物根际微生物对于维护土壤健康与肥力具有重要意义,植物根际富集的有益微生物可产生抗菌物质或激活植物免疫系统,从而达到减少病原微生物危害的作用[25]。另一方面,根际微生物通过自身代谢作用促进土壤物质的循环,进而影响植物的生长发育[26]。
本研究中,在门分类水平上,番茄实生和嫁接植株根际共有的优势细菌门种类并未发生变化,但嫁接改变了优势细菌门种类的相对丰度占比。其中,与桃太郎和新砧处理植株相比,桃太郎/新砧处理植株根际土壤的放线菌门和厚壁菌门丰度占比增加;与桃太郎和宝砧处理植株相比,桃太郎/宝砧嫁接植株根际土壤的绿弯菌门、酸杆菌门、芽单胞菌门细丰度占比增加,这与沈仁芳和赵学强[19]的研究得出嫁接可以改变植株根际土壤中微生物的丰度占比的结果一致。其中酸杆菌门、放线菌门和厚壁菌门是对植物有益的菌群,酸杆菌门细菌具有分泌纤维素和木质素降解酶,加速土壤中木质素和纤维素降解,提升土壤碳源循环的功能[27];而放线菌门细菌在促进植物养分利用、植株激素调节和维持土壤健康方面具有重要作用[28];厚壁菌门细菌在固氮、降解土壤中难溶化合物、抑制病原微生物生长等方面发挥重要作用[29],表明番茄嫁接处理后通过根际土壤微生物的活动可以创造更有利的环境条件。在各处理番茄植株根际土壤的优势真菌门类中,与实生植株相比,嫁接植株根际土壤中壶菌门丰度占比增加,而子囊菌门丰度占比减少。壶菌门真菌是一类可以游动的真菌类型,在自然界分布广泛,其相对丰度占比与土壤有机碳含量相关[30],同时还可以影响植物次生代谢物质合成[31];而子囊菌门真菌则是可以引起植物病害发生[32],不利于植物的生长发育,这表明嫁接不仅可以使有益微生物数量增加,还减少了有害微生物的数量,这是番茄嫁接处理可以提高植株抗病性的原因之一。
在属分类水平上,与桃太郎和新砧实生植株相比,桃太郎/新砧处理植株根际土壤中鞘氨醇单胞菌属、芽孢杆菌属、链霉菌属、unclassified_f__Micrococcaceae、norank_f__norank_o__Gaiellales细菌属丰度占比提升;与桃太郎处理植株相比,桃太郎/新砧处理植株根际土壤增加特有优势栖大理石菌属(Marmoricola)。与桃太郎和宝砧处理植株相比,桃太郎/宝砧处理植株根际土壤中norank_f__norank_o__norank_c__Subgroup_6、类诺卡氏菌、norank_f__norank_o__norank_c__Gitt-GS-136、norank_f__norank_o__norank_c__KD4-96、norank_f__JG30-KF-CM45属细菌丰度占比增加,相反鞘氨醇单胞菌属、芽孢杆菌属、德沃斯氏菌属、norank_f__norank_o__Gaiellales、unclassified_f__Micrococcaceae细菌属丰度占比下降,同时嫁接植株根际土壤拥有特有优势norank_f__norank_o__norank_c__JG30-KF-CM66菌属。鞘氨醇单胞菌属细菌具有促进土壤营养物质循环和降解复杂芳香族化合物的功能[33],说明嫁接处理可以有效改善番茄根际土壤中的养分循环以及难分解物质的转化;芽孢杆菌属和链霉菌属细菌通过代谢作用可以产生多种化合物,从而影响其他有害微生物生长[34-35],表明番茄嫁接处理可以改变根际土壤微生物的群落组成,塑造更加有益的菌群。
番茄实生与嫁接植株根际土壤中有5种共有优势真菌属。其中,与桃太郎和新砧处理植株相比,桃太郎/新砧处理植株根际土壤中根囊壶菌属、unclassified_k__Fungi、分枝孢子霉属真菌丰度占比增加;与桃太郎和宝砧处理植株相比,桃太郎/宝砧处理植株根际土壤中青霉属、根囊壶菌属真菌丰度占比增加。青霉属和根囊壶菌属真菌可以产生肽类、生物碱和萜类等多种次级代谢产物,同时还可产生抗生素和抑制有害病原微生物的作用[36-37];此外,与实生植株根际土壤相比,嫁接植株根际土壤中减少或缺失了具有致病作用的曲霉菌属、镰刀菌属和蛛网霉属等有害真菌属,表明番茄嫁接亦可以调控根际土壤真菌群落组成,从而增强番茄植株的抗病性。
综上所述,与对应番茄实生植株相比,嫁接不仅改变了番茄植株根际土壤细菌群落组成,同时亦改变了真菌群落组成。其中,放线菌门、厚壁菌门、鞘氨醇单胞菌属、芽孢杆菌属、链霉菌属、unclassified_f__Micrococcaceae、norank_f__norank_o__Gaiellales细菌是桃太郎/新砧处理植株根际土壤中优势细菌门和属;绿弯菌门、酸杆菌门、芽单胞菌门、类诺卡氏菌属、norank_f__norank_o__norank_c__Subgroup_6、norank_f__norank_o__norank_c__Gitt-GS-136、norank_f__norank_o__norank_c__KD4-96、norank_f__JG30-KF-CM45细菌是桃太郎/宝砧处理植株根际土壤中优势细菌门和属。另外,栖大理石菌属与norank_f__norank_o__norank_c__JG30-KF-CM66属细菌是番茄嫁接植株根际土壤中的特有优势细菌属。在真菌分类水平上,壶菌门是桃太郎/新砧和桃太郎/宝砧处理植株根际土壤中占比增加的优势真菌门;根囊壶菌属、unclassified_k__Fungi、分枝孢子霉属是桃太郎/新砧嫁接植株的根际土壤中优势细菌属,青霉属、根囊壶菌属是桃太郎/宝砧嫁接植株的根际土壤中优势细菌属。此外,番茄嫁接植株根际土壤中还拥有比实生植株更丰富的微生物OTU数量。
4 结 论
与实生植株相比,嫁接不仅改变了番茄植株根际土壤微生物群落丰度占比,而且改变了根际土壤微生物的群落组成。番茄嫁接植株根际土壤微生物群落组成的改良是嫁接植株抗性增强的重要原因之一。
