低温沙藏及外源激素添加对唐古特瑞香种子萌发及生理生化的影响
2024-03-12贾袭伟久西加郭晓霞徐宠然曾翠云陈红刚
贾袭伟,久西加,郭晓霞,徐宠然,马 潇,曾翠云, ,陈红刚, ,杜 弢,∗
(1.甘肃中医药大学药学院,甘肃 兰州 730000; 2.甘肃省药品检验研究院,甘肃 兰州 730070; 3.西北中藏药协同创新中心,甘肃 兰州 730000)
唐古特瑞香DaphnetanguticaMaxim.来源于瑞香科瑞香属,又称陕甘瑞香、甘肃瑞香、甘青瑞香、冬青草等,藏药称“省相那玛”,是中药祖师麻的基原植物之一[1],主要分布于我国西北、西南等海拔2 700 ~3 100 m 的高山林下、林缘及灌木丛中,其根皮和茎皮入药,具有祛风除湿、散寒止痛、活血散瘀功效[2]。唐古特瑞香作为常绿灌木,具有较高的生态价值和观赏价值[3],但其种子存在休眠期长、发芽率低、种群开拓性差等问题,资源再生困难,且野生资源过度采挖,难以满足市场需求[4]。人工栽培可能是缓解资源短缺的主要手段,因此开展唐古特瑞香种子休眠研究具有重要的实际意义和应用价值。
种子休眠是植物维持生存或适应环境变化的重要特性,有利于种子顺利度过逆境,在环境适宜的时候生根发芽,繁衍后代[5-6]。Baskin 把种子休眠主要分为生理休眠、物理休眠、形态休眠、形态生理休眠及复合休眠5 大类型[7]。目前唐古特瑞香种子休眠类型及原因仍不明确,相关研究报道较少,赵洁等[8]考察变温沙藏和不同浓度赤霉素对唐古特瑞香种子萌发的影响,发现相比自然条件下,发芽率小幅提高,但种子基础生物学指标、休眠原因、休眠类型及沙藏过程中种子内部生理生化特性未见报道。本实验以当年采收的唐古特瑞香种子为研究对象,通过种子吸水、种皮硬度测定、沙藏、外源激素浸种、种子发芽等试验,探究唐古特瑞香种子休眠破除方法,为唐古特瑞香育苗技术的制定提供参考。
1 材料
1.1 药物 唐古特瑞香种子于2020 年购买于武威泰康制药有限公司,经甘肃中医药大学药学院杜弢教授鉴定为唐古特瑞香DaphnetanguticaMaxim.的种子。
1.2 试剂 6-苄氨基腺嘌呤(批号CB291530100)、赤霉素(批号616H022) 均购自南京吉特恩生物科技有限公司;蛋白定量测定试剂盒(批号A045-2-2)、植物可溶性糖检测试剂盒(批号A145-1-1)、植物淀粉含量测试盒(批号A148-1-1) 均购自南京建成生物工程研究所; IAA (批号QC7898)、GA (批号 QC5680)、ABA (批号 QC4585)ELISA 检测试剂盒均购自上海钦诚生物科技有限公司。
1.3 仪器 BSA224S-CW 电子天平(北京赛多利斯科学仪器有限公司); GTOP-500 Y 智能光照恒温培养箱(浙江托普仪器有限公司); Leica-CM 1950 冷冻切片机[徕卡显微系统(上海) 贸易有限公司]; 543OR 超速冷冻离心机(常州金坛良友仪器有限公司); Vortex-Genie 2T 漩涡混合离心机(北京海天友诚科技有限公司); T6 新世纪紫外-可见分光光度计(北京普析通用仪器有限责任公司); THZ-82 气浴恒温震荡箱(常州金坛恒丰仪器制造有限公司);HH-4 数显恒温水浴锅(江苏金坛市环宇科学仪器厂)。
2 方法与结果
2.1 试验设计 除去瑞香种子残留果皮,水沉去除漂浮在水面上的瘪子,剩余种子置阴凉处晾干,室温条件下贮存备用。挑选饱满、无霉变、完整无机械损伤的种子装入尼龙网袋中,分层储藏于含水量为20%的湿沙盒内,沙盒置于4 ℃冰箱中,沙藏周期为120 d,每20 d 取样1 次,测定内含物、内源激素含量。基于前期发芽预实验的条件选择,同时将各沙藏阶段的种子分别以6-苄氨基嘌呤(6-BA,20 mg/L)、赤霉素(GA,300 mg/L)、蒸馏水 (CK) 浸种24 h后于25 ℃培养箱内进行暗培养,统计发芽率、发芽时滞天数等指标,每组实验重复3 次。
2.2 研究方法
2.2.1 唐古特瑞香种子千粒重测定 参照GB/T3543 标准对唐古特瑞香种子的千粒重进行测定[9]。
2.2.2 唐古特瑞香种子吸水率测定 随机称取风干的唐古特瑞香种子1.0 g,浸泡于蒸馏水中,分别在2、4、6、8、10、12 h 取出,用滤纸吸干种子表面水分,精密称定(0.000 1 g),重复3 次[10-11],直至种子质量变化范围在0.005 g 内时结束测定,计算种子吸水率。公式为吸水率=(Wt-W) /W×100%,式中Wt 为种子吸水后重量,W为种子干重。
2.2.3 唐古特瑞香种皮硬度测定 随机取唐古特瑞香种子3 份,每份30 粒,于谷物种皮硬度测定仪中测定,计算平均值[12]。
2.2.4 发芽指标测定 定期取不同沙藏阶段的唐古特瑞香种子各450 粒,用2%次氯酸钠溶液浸泡消毒15 min,随后蒸馏水冲洗30 min 用以发芽试验,分别以300 mg/L GA 和20 mg/L 6-BA 浸种处理24 h,以蒸馏水浸种处理24 h 为对照(CK),最后播于纸床上,每皿播种50 粒,重复3 次,在25 ℃恒温暗处培养,及时补足水分,统计发芽种子数,计算发芽率,公式为发芽率=发芽种子粒数/总种子粒数×100%。发芽时滞天数自种子播于纸床上开始计算,持续到第一粒种子发芽所需时间。
2.2.5 生化指标测定 在低温沙藏期间,定期取种子,经液氮冷冻后,贮存于-80 ℃冰箱中。使用酶标仪,通过考马斯亮蓝法测定可溶性蛋白; 蒽酮比色法测定可溶性糖;酸水解法和蒽酮比色法测定淀粉; 酶联免疫分析方法(ELISA) 测定样品中赤霉素(GA)、生长素(IAA) 及脱落酸(ABA) 等内源激素含量,具体方法参照各试剂盒说明书[13-14]。
2.3 数据分析 使用Microsoft Excel 2010 软件进行数据记录并整理,采用SPSS 19.0 软件对试验中各测定指标数据进行差异显著性分析,运用软件Origin 2019 软件制图,图中的结果均以(±SE) 表示。
2.4 结果分析
2.4.1 唐古特瑞香种子形态、千粒重、种皮硬度及吸水率变化 唐古特瑞香种子发芽图见图1A。唐古特瑞香果实脱去果肉后其种子外种皮呈黑褐色,外形圆润饱满,为双子叶无胚乳种子,种胚形态健全,见图1B。当年采收的唐古特瑞香种子千粒重59.40 g,种壳硬度3.66 kgf,未除去外种皮的种子22 h 可吸水饱和。

图1 唐古特瑞香种子发芽(A) 和形态特征图(B)
2.4.2 不同沙藏时间对唐古特瑞香种子发芽时滞天数的影响 随着沙藏时间延长,唐古特瑞香种子在各个沙藏阶段发芽时滞天数逐渐缩短,未沙藏种子发芽时滞天数为24 d;沙藏20 d 后,发芽时滞天数缩短为10 d; 沙藏60 d 后,各阶段种子发芽时滞天数趋于稳定,缩短为4 d,见图2。
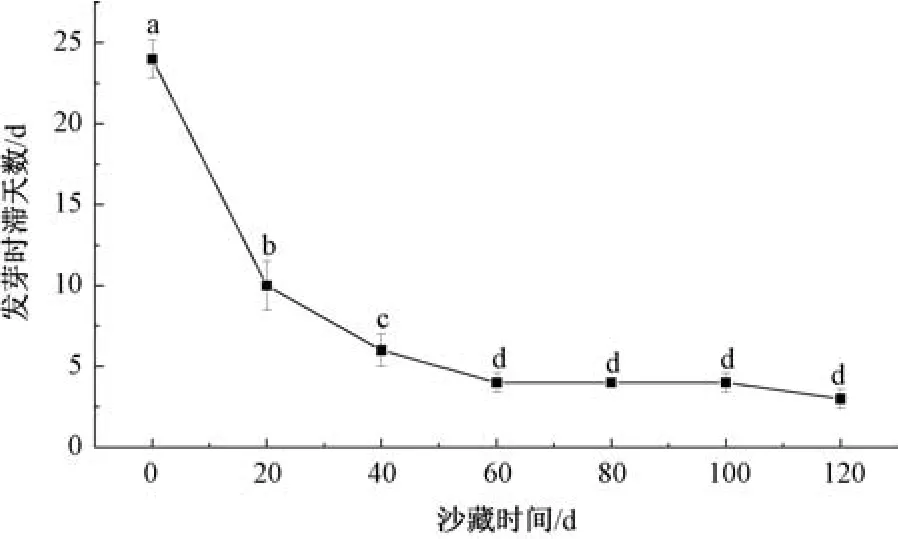
图2 沙藏时间对种子发芽时滞天数的影响
2.4.3 不同浸种处理对各阶段沙藏种子发芽率的影响 图3 表明,随着沙藏时间的延长,外源激素对种子萌发影响显著,20 mg/L 6-BA 浸种处理在沙藏60 d 时发芽率最高,为21.33%; 300 mg/L GA 浸种处理在沙藏80 d 时发芽率最高,为17.33%; 而清水浸种(CK) 在各沙藏阶段中发芽率均不足5%。综上所述,沙藏60 ~80 d 最有利于种子萌发,但必须结合外源激素浸种手段以提高发芽率,单纯延长沙藏时间或施以外源激素浸种处理并不能有效打破唐古特瑞香种子的休眠。
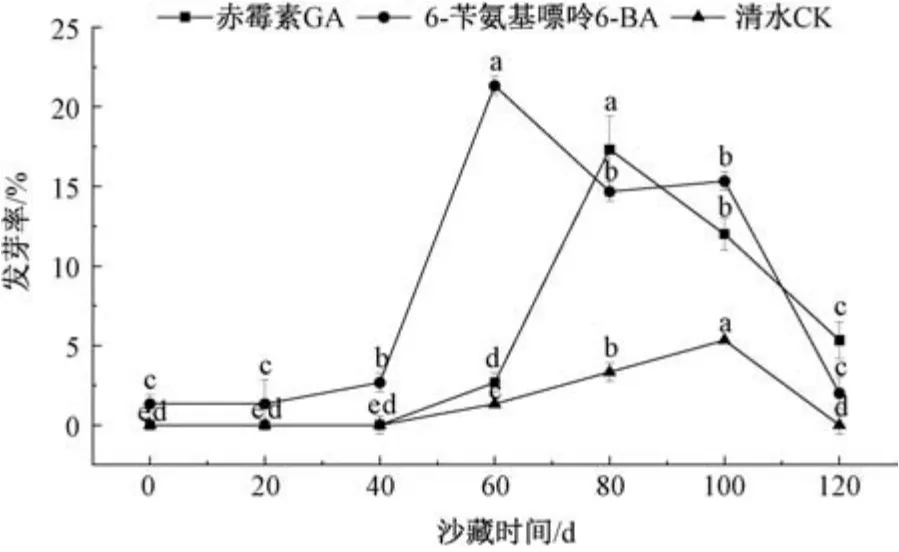
图3 不同浸种处理对沙藏种子发芽率的影响
2.4.4 不同沙藏时间对唐古特瑞香种子内含物含量的影响
2.4.4.1 对可溶性蛋白质含量的影响 由图4 可知,沙藏初期可溶性蛋白含量下降平缓,其含量在60 d 时出现最小值11.237 mg/g,60 d 后又平缓上升,总体呈下降趋势。随着沙藏时间的持续延长,除60 d 时可溶性蛋白含量显著低于其他阶段外,其他沙藏阶段可溶性蛋白含量差异不显著。可溶性蛋白质主要参与各种酶蛋白的代谢或作为基础供能物质被消耗,经过一系列的生理活动最终降解为氨基酸,为种子休眠的解除和萌发提供足够的氮源,其含量变化可以间接反映种子内部生命活动代谢的强弱[15]。
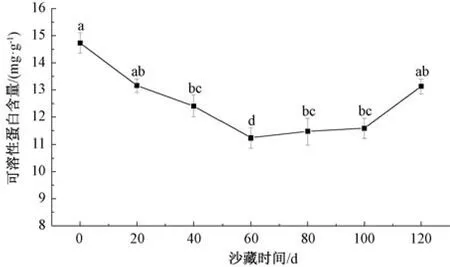
图4 沙藏时间对唐古特瑞香种子可溶性蛋白含量的影响
2.4.4.2 对淀粉含量的影响 由图5 可知,随着沙藏时间的延长,唐古特瑞香种子淀粉含量总体呈下降趋势,其中沙藏20~40 d 时,淀粉含量下降迅速,40 d 后下降较为平缓,120 d 出现最小值为1.274 mg/g。未沙藏种子可溶性淀粉含量显著高于各阶段沙藏处理,说明在沙藏过程中,呼吸作用使种子内部的储能物质发生转变,淀粉逐渐分解,转化成可溶性糖等,供应种子正常的生理代谢消耗,为种子萌发做前期准备[16]。
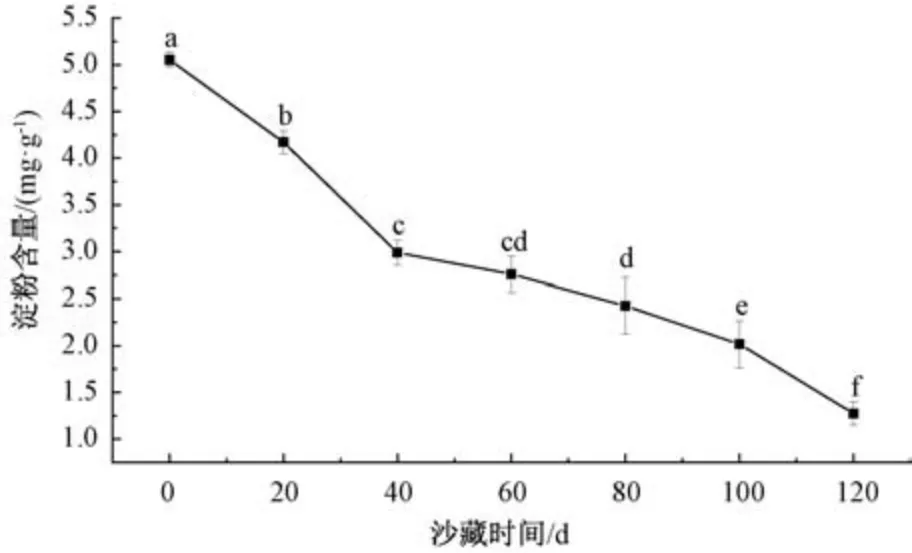
图5 沙藏时间对唐古特瑞香种子可溶性淀粉含量的影响
2.4.4.3 对可溶性糖含量的影响 由图6 可知,沙藏时间在20~60 d 期间,可溶性糖含量呈下降趋势,60 d 达到最小值9.331 mg/g,其中20~40 d 可溶性糖含量下降最为迅速,60~100 d 可溶性糖含量有所升高,在100 d 时达到峰值,120 d 时含量又有所下降,在整个沙藏过程中种子可溶性糖含量总体呈下降趋势。纵观整个沙藏过程种子中可溶性糖含量变化曲线,20 ~60 d 可能是由于此时种子内部生理代谢活动旺盛,大量消耗了子叶中贮藏的可溶性糖,在60~100 d 种子中可溶性糖含量增加,当达到100 d 时其含量显著高于60 d,从沙藏60 d 开始,唐古特瑞香种子内部可溶性糖消耗速率减缓,可能因淀粉分解使得可溶性糖含量有所升高,种子内部代谢活动发生变化。
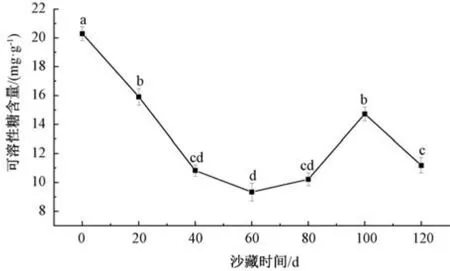
图6 沙藏时间对唐古特瑞香种子可溶性糖含量的影响
2.4.5 不同沙藏时间对唐古特瑞香种子内源激素含量的影响
2.4.5.1 对IAA 含量的影响 如图7 所示,随着沙藏时间的延长,吲哚乙酸(IAA) 含量呈现“N” 形曲线变化,在0~40 d 和80~120 d 时出现2 次增长; 其最高峰出现在沙藏40 d,IAA 含量为4.184 ng/g,除与沙藏120 d 时比较差异不显著外,显著高于其他各沙藏阶段; 80 d 时IAA 含量最低为2.187 ng/g,显著低于其他沙藏阶段; 在40~80 d 期间IAA 含量平缓下降。有研究表明,IAA 含量在适宜的浓度范围内对种子解除休眠有一定的促进作用,间接影响种胚的发育和其他激素水平,具有释放种子休眠、促进种子萌发的作用[17],但在本实验中不同沙藏阶段IAA 含量并未表现出规律性的变化。
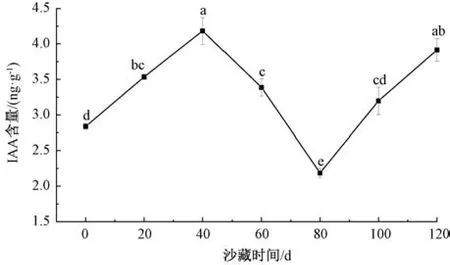
图7 沙藏时间对唐古特瑞香种子IAA 含量的影响
2.4.5.2 对GA 含量的影响 由图8 可知,唐古特瑞香种子在整个沙藏过程中赤霉素(GA) 含量呈上升趋势,各沙藏阶段的种子GA 含量均显著高于未沙藏种子,在沙藏初期(20、40 d) 含量增加平缓且无显著性差异,60 d 时含量显著升高,此后稳定,一直持续到100 d; 100 ~120 d 时其含量又急剧增加,120 d 达到最大值116.656 ng/g。赤霉素(GA) 在种子处理中应用广泛,对多种植物种子萌发具有促进作用[17],本实验中特别是沙藏后期,GA 含量逐步升高,同时发芽率亦表现出升高的趋势,说明GA 含量增加对唐古特瑞香种子休眠的解除具有积极作用,能够提高发芽率,很有可能是唐古特瑞香种子萌发的主要促进物质。

图8 沙藏时间对唐古特瑞香种子GA 含量的影响
2.4.5.3 对GA/ABA 比值的影响 种子内源激素的消长平衡调控着种子的休眠水平,大量研究表明种子内源激素的绝对含量只是调控机制中的一个方面,然而激素间的相对比值关系直接影响内源激素最终的调控效果。近年来,赤霉素与脱落酸(GA/ABA) 的比值受到更多科研工作者的关注[15],ABA 被认为是最常见的种子萌发抑制剂,与GA存在拮抗作用,正向调节种子休眠时间,抑制其萌发[18]。图9 显示,GA/ABA 值在整个沙藏阶段呈现双峰增长曲线,先缓增再急减,又急增后降低,GA/ABA 值在40 d 时急剧上升,60 d 和80 d 两个阶段均保持在较高水平,分别为12.48、10.71,与此同时60 d 和80 d 两个阶段唐古特瑞香种子发芽率显著提高,分别为21.33%和17.33%,80 d 后GA/ABA 值急剧下降并低于沙藏前期,种子发芽率随之降低,最低为2%。结果显示,GA/ABA 比值与唐古特瑞香种子发芽率有着密切联系。
3 讨论
3.1 唐古特瑞香种子休眠解除与营养物质转化的关系 本实验通过对唐古特瑞香种子在4 ℃恒温条件下进行为期120 d 的低温沙藏处理,沙藏过程中淀粉、可溶性糖含量总体呈降低趋势,作为种子呼吸作用的基础物质被消耗利用,与许美玲[19]研究结果一致。沙藏前期变化幅度较大,后期变化趋于平缓,这可能是因为种子沙藏前后环境温度不同,种子内部代谢增强,淀粉分解为可溶性糖的速率提高,以增强种子抗寒机能,便于种子保持正常的生理活性,与沙藏后种子内部新陈代谢活动改变有关[20-22]。可溶性蛋白质在沙藏前期可能作为基础供能物质有所消耗; 60 d 后含量逐渐增加,可能由于种子内部代谢规律发生变化。
3.2 唐古特瑞香种子休眠解除与内源激素的关系 在内源激素含量中,IAA 含量在整个沙藏过程中变化呈“N” 形曲线,40~80 d 时呈下降趋势,并在80 d 达到最低点。有研究表明IAA 含量在适宜浓度范围与其他内源激素相互作用可间接影响种胚发育[23]; 也有研究表明IAA 低浓度可促进种子萌发,高浓度对种子萌发具有抑制作用[24]。IAA 含量在2.187~3.386 ng/g (对应时期为沙藏60 ~80 d) 范围内有利于唐古特瑞香种子萌发,但整个沙藏过程中未表现出一致的变化规律。GA 作为一种信号物质,其促进种子发芽的作用已被广泛认可,本实验中GA 含量在沙藏过程中总体呈上升趋势,各沙藏阶段GA 含量均显著高于未沙藏种子,这与张成才等[25]关于百蕊草种子休眠研究结果一致。相对单一激素含量来讲,不同激素含量间的平衡消长变化对于种子休眠与发芽的调控作用更为重要,尤其是生长促进类激素与生长抑制类激素含量间的平衡[26]。有研究指出,ABA 是种子发芽抑制类物质,而GA 与ABA 的比值是决定种子休眠是否解除的关键因素[27],从GA/ABA 值来看,在整个沙藏阶段呈现“双峰” 增长曲线,在60 d 时出现了短暂的峰值后转为平缓下降趋势,60 d 时以20 mg/L的6-BA 浸种后种子发芽率明显提高,这与前人关于GA/ABA 值是调控种子继续休眠或萌发的重要因素的研究结论相似[28]。但沙藏后未用外源激素浸种处理的种子发芽率仍不足5%,推测在唐古特瑞香种子整个沙藏过程中,其内源激素代谢并不能达到理想的平衡状态促使种子萌发,需用适量的外源激素帮助其打破休眠状态,这与赵洁等[8]研究结论相似,尽管模拟其生境温度时设置了变温沙藏试验,但与本实验4 ℃恒温沙藏的种子发芽率接近。
4 结论
唐古特瑞香种子子叶发达,种胚形态健全,种子吸水率良好,故不存在形态休眠和吸水障碍; 在内源激素方面,GA 与ABA 含量的比值可能是解除唐古特瑞香种子休眠的关键; 4 ℃沙藏60 d 时以20 mg/L 6-BA 浸种24 h,或沙藏80 d 时以300 mg/L GA 浸种24 h 可有效破除休眠现象,提高种子发芽率。
