ε-聚赖氨酸对西番莲果实采后病害与抗病物质代谢的影响
2024-03-10林育钊陈蕾伊陈佳怡蒋璇靓郑金水陈洪彬
林育钊,陈蕾伊,陈佳怡,蒋璇靓,郑金水,陈洪彬,3,*
(1.泉州师范学院海洋与食品学院,福建 泉州 362000;2.泉州市农业科学研究所,福建 泉州 362212;3.近海资源生物技术福建省高校重点实验室(泉州师范学院),福建 泉州 362000)
西番莲(Passiflora caeruleaL.),亦称鸡蛋果或者百香果,在我国广西、福建、广东、台湾等地区广泛栽培[1-2]。西番莲果实含有丰富的矿物质、酚类、有机酸、维生素等营养成分[2-3],因而具有较高的食用价值。此外,香气也是影响西番莲果实品质的重要特性,丁酸乙酯、己酸乙酯、丁酸己酯、醋酸乙酯、α-松油醇、己醛、β-月桂烯等是果实中重要的香味成分,使得果实具有浓郁的香气[4-5]。然而,西番莲果实大多成熟和采收于高温高湿季节,采后极易发生果实感病、腐烂等品质败坏现象,不利于采后贮藏及品质保持[2]。病原菌的侵染是诱导西番莲果实采后感病、腐烂发生的关键因素,严重限制果实采后贮藏、远距离贮运与销售,也是西番莲果实产业健康发展需要迫切解决的关键问题[6]。目前,使用化学杀菌剂(例如丙氯灵、二氯异氰尿酸钠等)虽然能够控制西番莲果实采后病害发生,但是由此带来的药物残留、人体健康损伤及环境污染等问题突出,因而化学杀菌剂不适于西番莲果实采后病害控制[7]。因此,很有必要寻找一种安全、有效的采后处理技术,以控制西番莲果实采后病害、腐烂发生,进而提高西番莲果实采后贮藏品质。
ε-聚赖氨酸(ε-poly-L-lysine,ε-PL)是从白色链霉菌(Streptomyces albulus)NO.346中分离而得[8-9]。它是一种安全、有效的抑菌多肽[10-11],由25~35 个L-赖氨酸残基通过ε-氨基和α-羧基之间形成的酰胺键所形成[11-12]。ε-PL因其具备强抗菌能力、安全性、生物降解性等特性,可在食品中应用[11,13]。有研究报道,ε-PL可控制龙眼[9-10]、草莓[14]等果实采后病害发生,进而稳定果实品质。此外,ε-PL控制采后果实病害的发生,可能是由于ε-PL能提高果实采后抗病相关酶的活性或者提升木质素等抗病物质的含量,进而诱导果实采后抗病性[10,15-16]。
另外,笔者在前期研究中发现,可可毛色二孢(Lasiodiplodia theobromae)是致使福建黄金西番莲果实采后腐烂的主要病原菌之一,严重降低了果实采后品质。然而,目前鲜见ε-PL提高黄金西番莲果实抗病物质代谢水平而增强采后抗病性,进而抑制果实采后发生病害的研究报道。因此,本研究以福建省近年来快速发展的黄金西番莲果实为研究对象,研究ε-PL抑制L.theobromae接种黄金西番莲果实采后病害发生及其与果皮细胞膜透性、果实抗病性及抗病物质代谢的关系,旨在阐明ε-PL抑制L.theobromae侵染所致黄金西番莲果实采后病害发生的作用机制,为ε-PL维持黄金西番莲果实采后品质提供科学依据。
1 材料与方法
1.1 材料、菌株与试剂
以福建省主栽黄金西番莲品种‘福建百香果3号’(Passiflora caeruleaL.cv.Fujian Baixiangguo NO.3)果实为研究对象,果实(8 成熟)采摘于福建南安溪美镇缘味家庭农场,挑选色泽一致、大小均一、无感病、无损伤、健康的果实,将经过清水清洗、次氯酸钠浸泡10 s(表面消毒)、无菌水(sterile distilled water,SDW)清洗等步骤后的果实用于后续研究。
L.theobromae由本课题组自主分离、鉴定及保存。另外,参考Zhang Shen等[17]的方法制备L.theobromae孢子悬浮液,将L.theobromae接种至燕麦米糠琼脂培养基,在光照下连续培养21 d,用SDW洗脱成熟孢子,制成1×105/mL的悬浮液。
ε-PL 郑州拜纳佛生物工程股份有限公司;氯化钡、苯丙氨酸、葡萄糖-6-磷酸二钠、磷酸氢二钠、邻苯二酚、磷酸二氢钠、氯化镁、抗坏血酸、亮抑酶酞、丙三醇、三氯乙酸、反式肉桂酸、三磷酸腺苷、昆布多糖、柠檬酸、辅酶A、p-香豆酸、愈创木酚、四硼酸钾、3,5-二硝基水杨酸等(化学纯)国药集团化学试剂有限公司。
1.2 仪器与设备
CP224C型分析天平 奥豪斯仪器上海有限公司;Infinite M 200 Pro型酶标仪 瑞士帝肯公司;SQ510C型高压灭菌器 重庆雅马拓科技有限公司;S230型电导率仪 梅特勒-托利多仪器(上海)有限公司;H2050R型离心机 湖南湘仪仪器有限公司;PRX-450A型智能人工气候箱 浙江宁波赛福实验仪器有限公司;HH-4型数显恒温水浴锅 国华电器有限公司。
1.3 方法
1.3.1 果实采后处理与损伤接种
在预实验中,采用0(对照)、50、100、150 mg/L和200 mg/Lε-PL浸泡西番莲果实10 min(浸泡时间参照Sun Junzheng等[10]的方法),晾干,采用无菌打孔器在果实赤道面打孔1 个(直径5 mm,深度3 mm,下同),并在孔中接种20 μLL.theobromae孢子悬浮液,晾干之后装盘(10 个/盘),再用聚乙烯薄膜袋包装,置于(28±1)℃、90%相对湿度(relative humidity,RH)的恒温箱中贮藏7 d。结果发现,100 mg/Lε-PL处理再接种L.theobromae的西番莲果实具有较低的病斑直径,果实感病症状不明显。因此,选用100 mg/Lε-PL进行后续实验。
在获得ε-PL最佳浓度的基础上,将挑出来的西番莲果实分成2 组,进行以下处理(每组均设3 个重复):1)接种L.theobromae组:SDW浸泡果实10 min后,晾干,打孔,接种20 μL浓度为1×105/mL的L.theobromae孢子悬浮液;2)ε-PL+接种L.theobromae组:ε-PL浸泡果实10 min后,晾干,打孔,接种20 μL浓度为1×105/mL的L.theobromae孢子悬浮液。
西番莲果实经上述处理之后,晾干,装盘(10 个/盘),包装聚乙烯薄膜袋,贮藏在(28±1)℃、90% RH的恒温箱中。在贮藏期,随机取样观察果实外观变化,测定果实病斑直径,并从果实发病部位外10~30 mm内取样,测定相关指标。
1.3.2 果皮细胞膜透性的测定
参考Gong Di等[18]的方法测定西番莲果实的果皮细胞膜透性,结果用%表示。
1.3.3 果实病斑直径的测定
参考Gong Di等[18]的方法使用十字交叉法测定西番莲果实的病斑直径。
1.3.4 果皮木质素质量分数的测定
参照郭欣[7]与Sun Junzheng[10]等的方法测定西番莲果皮木质素质量分数,结果以%表示。
1.3.5 果皮抗病相关酶活力的测定
参考Sun Junzheng等[10]的方法进行测定西番莲果皮苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)活力;果皮4-香豆酰辅酶A连接酶(4-coumarate CoA ligase,4-CL)、肉桂酸-4-羟化酶(cinnamate-4-hydroxylase,C4H)、肉桂醇脱氢酶(cinnamyl alcohol dehydrogenase,CAD)活力的测定均参照郭欣[7]、Sun Junzheng[10]与Tang Jinyan[19]等的方法;果皮多酚氧化酶(polyphenol oxidase,PPO)和过氧化物酶(peroxidase,POD)活力参考郭欣等[7]的方法进行测定;果皮几丁质酶(chitinase,CHI)、β-1,3-葡聚糖酶(β-1,3-glucanase,GLU)活力参照Tang Jinyan等[19]的方法进行测定。另外,参照Bradford[20]的方法测定蛋白质含量,上述酶活力均以蛋白质含量进行计算。
1.4 数据处理
在本实验中,除测定病斑直径进行10 次重复之外,其他指标均设置3 次重复。利用Microsoft Excel 2019软件进行绘图;利用SPSS 22.0软件进行数据统计与相关性分析。
2 结果与分析
2.1 果实外观变化和病斑直径
由图1A可知,在贮藏0 d时,西番莲果实果皮呈现明亮的黄绿色,果实外形完整。然而,随着贮藏时间的延长,果实果皮色泽由黄绿色变成黄色,之后再变成褐色。对于接种L.theobromae组,从贮藏2 d开始,果实接种处快速长出菌丝,并由白色转变为黑色,果实感病加重;同时接种处伴有严重的黄褐色腐烂症状,并向周边扩散,果实腐败程度加剧。然而,与接种L.theobromae组比较,ε-PL+接种L.theobromae组的果实接种处菌丝生长较缓,果实感病症状较轻;同时接种处黄褐色腐烂症状较轻,腐烂症状向周边扩散速度较慢,果实腐败较不严重。

图1 ε-PL对接种L.theobromae西番莲果实外观(A)和病斑直径(B)的影响Fig.1 Effect of ε-PL on appearance (A) and lesion diameter (B) of L.theobromae-inoculated passion fruits
果实病斑直径是一个评价果实采后病害发生的重要指标。由图1B可知,两个处理组的西番莲果实病斑直径在贮藏0~1 d均为0,但是从贮藏1 d开始,表现出不同的变化趋势。相比于接种L.theobromae组,ε-PL+接种L.theobromae组病斑直径增大速度较为缓慢。基于显著性分析,ε-PL+接种L.theobromae组的病斑直径在贮藏4 d显著低于接种组(P<0.05),在贮藏5~7 d极显著低于接种组(P<0.01)。
因此,ε-PL能延缓接种L.theobromae西番莲果实采后病害的发生。
2.2 果皮细胞膜透性
完整的细胞膜对植物细胞正常生命活动具有重要作用[18],细胞膜的损伤将影响采后抗病性,一般用细胞膜透性表示细胞膜的损伤程度[21]。由图2可知,西番莲果实的果皮细胞膜透性于贮藏0~7 d快速升高。相比之下,ε-PL+接种L.theobromae组的果皮细胞膜透性在贮藏0~7 d低于接种L.theobromae组,并在贮藏2 d达到显著水平(P<0.05),在贮藏4~7 d达到极显著水平(P<0.01)。因此,ε-PL能延缓接种L.theobromae西番莲果实采后果皮细胞膜透性的上升。

图2 ε-PL对接种L.theobromae西番莲果实果皮细胞膜透性的影响Fig.2 Effect of ε-PL on cell membrane permeability in pericarp of L.theobromae-inoculated passion fruits
2.3 果皮木质素质量分数
木质素作为植物抗病系统中的重要组成,能抵抗病原菌入侵[7,10]。由图3可知,接种L.theobromae组的西番莲果实果皮木质素质量分数在贮藏0~5 d快速升高,随后快速下降。然而,ε-PL+接种L.theobromae组的果皮木质素质量分数在贮藏0~4 d快速升高,随后快速降低。进一步比较可知,与接种L.theobromae果实的果皮木质素质量分数对比,ε-PL+接种L.theobromae组具有较高水平,并在贮藏3、5~7 d具有显著水平(P<0.05),在贮藏4 d具有极显著水平(P<0.01)。

图3 ε-PL对接种L.theobromae西番莲果实果皮木质素质量分数的影响Fig.3 Effect of ε-PL on lignin content in pericarp of L.theobromaeinoculated passion fruits
因此,ε-PL可提升接种L.theobromae西番莲果实果皮木质素水平。
2.4 果皮PAL、C4H、4-CL、CAD和POD活性
PAL、C4H、4-CL、CAD与POD是苯丙烷代谢途径中的重要酶,参与合成木质素[7,10,22]。由图4A可知,接种L.theobromae组的西番莲果实果皮PAL活性在贮藏0~1 d较快降低,1~2 d较快升高,2~3 d较快降低,3~6 d加剧提高,之后快速减少。然而,ε-PL+接种L.theobromae组的果皮PAL活性于贮藏0~1 d快速上升,1~2 d快速减小,2~6 d急剧提高,6~7 d快速下降。与接种L.theobromae果实对比可知,ε-PL+接种L.theobromae组的果皮PAL活性在贮藏0~7 d具有较高水平,同时在贮藏3、7 d具有极显著较高水平(P<0.01),在贮藏4 d表现为显著较高水平(P<0.05)。
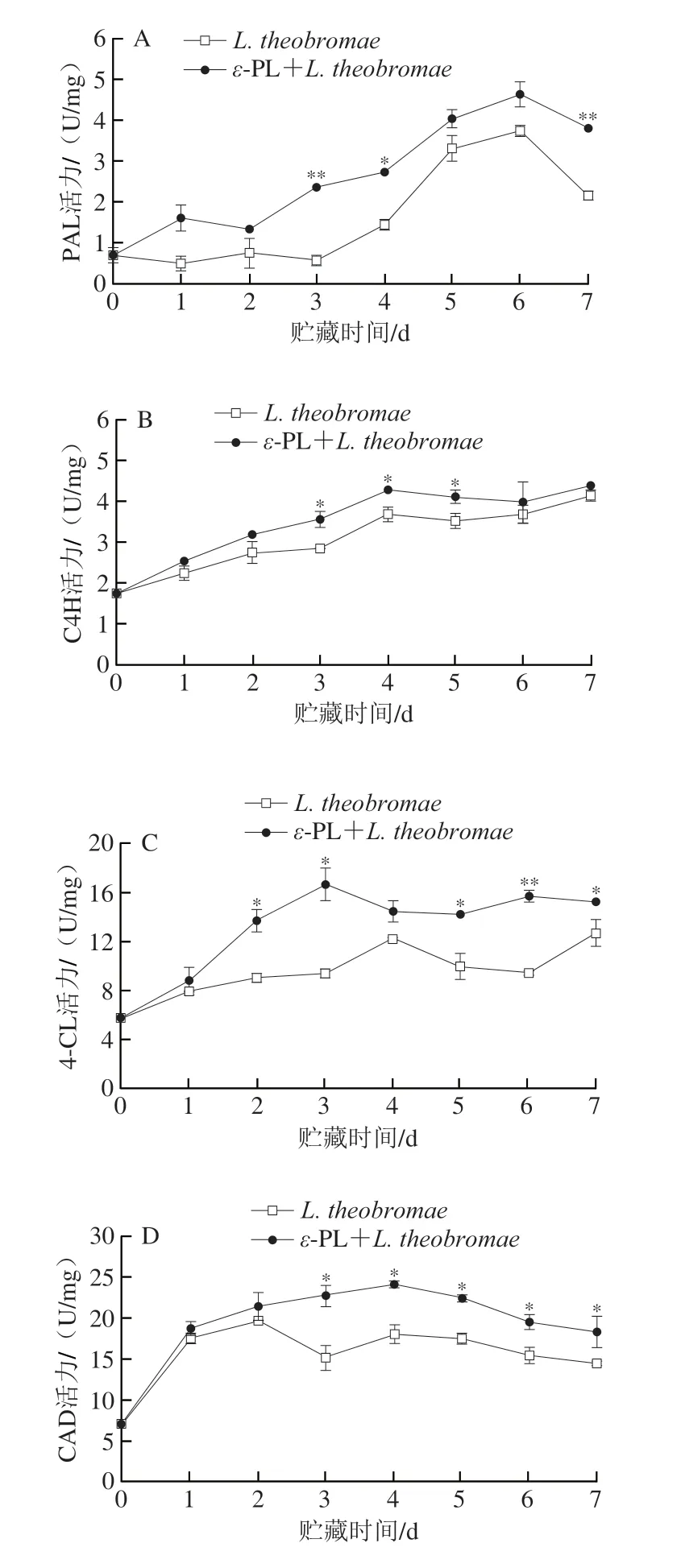

图4 ε-PL对接种L.theobromae西番莲果实果皮PAL(A)、C4H(B)、4-CL(C)、CAD(D)和POD(E)活性的影响Fig.4 Effect of ε-PL on activities of PAL (A),C4H (B),4-CL (C),CAD (D)and POD (E) in pericarp of L.theobromae-inoculated passion fruits
由图4B可知,接种L.theobromae组的西番莲果实果皮C4H活性在贮藏0~4 d加速上升,4~5 d较快降低,随后快速提高。然而,ε-PL+接种L.theobromae组的果皮C4H活性在贮藏0~5 d变化趋势与接种L.theobromae组类似,但在贮藏5~7 d却较快升高。另外,ε-PL+接种L.theobromae组的果皮C4H活性在贮藏0~7 d高于接种L.theobromae组,且于贮藏3~5 d显著高于接种L.theobromae组(P<0.05)。
由图4C可知,接种L.theobromae组的西番莲果实果皮4-CL活性于贮藏0~4 d快速提高,4~6 d快速下降,随后加速升高。但是,ε-PL+接种L.theobromae组的果皮4-CL活性在贮藏0~3 d加剧提升,3~5 d快速减小,5~6 d快速升高,6~7 d较快减少。另外,与接种L.theobromae组相比,ε-PL+接种L.theobromae组的果皮4-CL活性始终处于较高水平,且于贮藏2~3、5、7 d达到显著水平(P<0.05),在贮藏6 d具有极显著水平(P<0.01)。
由图4D可知,接种L.theobromae组的西番莲果实果皮CAD活性于贮藏0~2 d加剧上升,2~3 d加速降低,3~4 d快速提高,随后较快下降。然而,ε-PL+接种L.theobromae组的果皮CAD活性于贮藏0~4 d急剧上升,随后快速减少。此外,和接种L.theobromae果实对比,ε-PL+接种L.theobromae组在贮藏期维持较高果皮CAD活性,并在贮藏3~7 d具有显著较高水平(P<0.05)。
由图4E可得,接种L.theobromae组的西番莲果实果皮POD活性在贮藏0~1 d加速升高,1~4 d较快降低,4~5 d快速提高,5~6 d快速下降,6~7 d缓慢上升。但是,ε-PL+接种L.theobromae组的果皮POD活性于贮藏0~4 d加剧提高,4~5 d缓慢下降,之后加速降低。另外,ε-PL+接种L.theobromae组的果皮POD活性在贮藏2~7 d高于接种组,并在贮藏3 d有极显著水平(P<0.01),在贮藏4~5、7 d有显著水平(P<0.05)。
因此,ε-PL保持L.theobromae接种西番莲果实较高的果皮PAL、C4H、4-CL、CAD及POD活性。
2.5 果皮PPO活性
PPO可氧化酚类物质,从而产生对病原菌有毒的醌类物质[7,22-23]。由图5可知,接种L.theobromae组的西番莲果实果皮PPO活性在贮藏0~4 d加剧提高,4~5 d快速下降,5~6 d缓慢上升,6~7 d加速减少。然而,ε-PL+接种L.theobromae组的果皮PPO活性于贮藏0~3 d急剧上升,3~4 d快速降低,4~5 d快速提高,之后加速减少。此外,和接种组对比,ε-PL+接种L.theobromae组的果皮PPO活性均保持较高水平,同时在贮藏1 d具有显著水平(P<0.05),在贮藏3、5 d具有极显著水平(P<0.01)。

图5 ε-PL对接种L.theobromae西番莲果实果皮PPO活性的影响Fig.5 Effect of ε-PL on PPO activity in pericarp of L.theobromaeinoculated passion fruits
因此,ε-PL能维持接种L.theobromae西番莲果实较高水平的果皮PPO活性。
2.6 果皮GLU和CHI活性
GLU与CHI也是防御病原菌入侵果实的关键酶[24]。从图6A发现,接种L.theobromae组的西番莲果实果皮GLU活性于贮藏0~1 d快速降低,1~2 d快速上升,2~3 d快速减小,随后加速升高。但是,ε-PL+接种L.theobromae组的果皮GLU活性在贮藏0~1 d缓慢上升,1~2 d缓慢降低,2~5 d加剧上升,5~6 d快速减少,6~7 d快速提高。另外,和接种组比较,ε-PL+接种L.theobromae组于贮藏0~1、3~7 d保持较高果皮GLU活性,同时在贮藏3~7 d具有极显著水平(P<0.01)。

图6 ε-PL对接种L.theobromae西番莲果实果皮GLU(A)和CHI(B)活性的影响Fig.6 Effect of ε-PL on activities of GLU (A) and CHI (B) in pericarp of L.theobromae-inoculated passion fruits
从图6B发现,接种L.theobromae组的西番莲果实果皮CHI活性于贮藏0~1 d较快下降,1~5 d较快上升,5~6 d快速减小,6~7 d快速提高。但是,ε-PL+接种L.theobromae组的果皮CHI活性于贮藏0~1 d快速升高,1~3 d缓慢上升,3~5 d加速提升,5~6 d较快减少,随后急剧增高。另外,与接种组比较,ε-PL+接种L.theobromae组的果皮CHI活性在贮藏期维持较高水平,并在贮藏4~5 d达极显著水平(P<0.01),在贮藏6~7 d达显著水平(P<0.05)。
因此,ε-PL能保持接种L.theobromae西番莲果实较高的果皮GLU和CHI活性。
3 讨论
细胞膜损伤与病原菌入侵密切相关。较高的细胞膜透性是果实采后细胞膜结构丧失、抗病性下降、病害发生的促进因素[21,25-26]。在本研究中,接种L.theobromae组的西番莲果实果皮细胞膜透性(图2)和果实病斑直径(图1B)在贮藏期均呈现上升趋势。基于相关性分析可知,接种果实的病斑直径(图1B)与果皮细胞膜透性(图2)具有极显著的正相关关系(P<0.01),相关系数为0.917。由此可推断,L.theobromae侵染加速西番莲果实病害发生与细胞膜损伤有关。然而,与接种L.theobromae组相比,ε-PL+接种L.theobromae组在贮藏期具有较低果实病斑直径(图1B)、果皮细胞膜透性(图2)。因此,ε-PL能减轻接种L.theobromae果实果皮细胞膜损伤程度,稳定果实抗病性,从而减缓果实发生病害。这与Chen Yihui等[21]的研究结果一致,即水杨酸减缓龙眼果实发生病害,是由于水杨酸维持较好的果皮细胞膜完整性。
另外,木质素是果实抗病防御系统中的重要成分,当果实受到病原菌侵染时,迅速积累木质素,形成物理屏障,抑制病原菌进一步入侵[7,10,23]。本研究发现,接种L.theobromae组的西番莲果实果皮木质素质量分数在贮藏0~5 d不断累积上升,之后却呈现下降趋势(图3),而病斑直径在贮藏1~7 d不断升高(图1B)。这可能是因为在贮藏前期,L.theobromae入侵西番莲果实后,西番莲果实的抗病防御机制被激活,将不断累积木质素等抗病物质,进而提高果实抗病水平而应对病原菌早期入侵;但在L.theobromae侵染后期,由于果实的抗病物质(木质素等)水平下降,诱导果实抗病性丧失,进而引发果实发生病害。因此,L.theobromae侵染加速西番莲果实采后抗病性丧失、病害发生与其果实果皮木质素质量分数下降有关。进一步对比发现,在贮藏期,ε-PL+接种L.theobromae组具有较高的果皮木质素质量分数(图3),而具有较小的果实病斑直径(图1B)。因此,ε-PL抑制L.theobromae侵染所致西番莲果实采后病害发生,是由于ε-PL促进果实果皮累积木质素,进而增强果实采后抗病性。这与黄晓杰等[27]的研究结果一致,即茉莉酸甲酯延缓蓝莓果实发生病害与果实保持较高木质素水平有关。类似地,Ren Yanfang等[24]研究报道,硝普酸钠能提高芒果果实木质素含量,进而延缓果实采后炭疽病的发生。
此外,PAL、C4H、4-CL、CAD、POD、PPO、GLU与CHI等抗病酶在果实应对病原菌入侵反应中起着重要作用[7,10,22,24]。其中,PAL、C4H、4-CL、CAD、POD等抗病酶可参与木质素的合成及累积,在抵御病原菌侵染中具有关键作用[7,10,22-23]。PPO可催化酚类发生氧化而形成醌,进而抑制病原菌的生长[7,10]。GLU与CHI分别能降解病原菌细胞壁的β-1,3-葡聚糖与几丁质,从而抑制病原菌的生长[7,28-29]。本研究发现,相比于接种L.theobromae的西番莲果实,ε-PL维持接种L.theobromae西番莲果实在贮藏0~7 d具有较高的果皮PAL、C4H、4-CL、CAD活性,在贮藏2~7 d具有较高的果皮POD活性(图4),这促进了果实累积木质素;ε-PL也保持接种果实在贮藏0~7 d有较高的果皮PPO活性(图5),这有利于PPO催化酚类而生成醌,进而抑制L.theobromae对果实的侵染;ε-PL还维持接种果实在贮藏1、3~7 d有较高的果皮GLU活性,在贮藏0~7 d有较高的果皮CHI活性(图6),这有助于破坏L.theobromae细胞壁结构而抑制其生长。因此,ε-PL抑制L.theobromae侵染所致的西番莲果实采后病害发生,是由于ε-PL提高接种L.theobromae西番莲果实的果皮PAL、C4H、4-CL、CAD、POD、PPO、GLU及CHI等抗病相关酶的活性,累积果皮木质素等抗病物质,进而提高果实采后抗病性。类似地,Sun Junzheng等[10]研究表明,ε-PL能提升龙眼果实的PAL、CAD、C4H、4-CL与POD等抗病酶的活性,促进木质素的累积,进而增强果实采后抗病能力,延缓龙眼果实采后病害的发生。Liu Yongxiang等[28]研究发现,L-赖氨酸可以提高梨果实的GLU与CHI活性,进而提高果实抗病性而控制采后病害发生。另外,刘晓佳等[30]研究报道,苯并噻二唑可提升柑橘果实的PAL、POD、PPO、CHI和GLU等抗病酶活性,提升果实对病原菌入侵的抗性水平。
4 结论
综上,100 mg/Lε-PL可有效维持接种L.theobromae西番莲果实的果皮细胞膜完整性,还能提升接种果实与果皮木质素合成有关的相关酶(PAL、C4H、4-CL、CAD、POD等)活性而促进累积木质素,提高接种果实果皮的PPO、GLU及CHI等酶活性,从而增强果实采后抗病性,抑制L.theobromae侵染所致的果实采后病害发生。因此,ε-PL可作为一种安全的采后处理技术以增强西番莲果实的抗病能力,进而抑制果实采后发生病害、稳定其采后品质。
