基于材用云南松表型性状构建核心种质策略研究
2024-03-08付朴艳曹子林王晓丽
李 华,付朴艳,闫 熙,曹子林,王晓丽*
(1 西南林业大学 林学院,昆明 650224;2 西南林业大学 生态与环境学院,昆明 650224)
核心种质(core collection)的概念于1984年由Frankel[1]首次提出,李永祥等[2]认为核心种质具有异质性、代表性、实用性、动态性和有效性等特征。核心种质是以尽可能少的种质资源数量最大程度的代表整体种质的遗传多样性,提高森林资源利用效率,促进种质资源的整合开发与利用[3]。目前,主要基于表型性状[4-8]、分子标记、或结合分子标记与表型性状[9-11]方法构建农作物、木本植物等核心种质。研究表明,表型性状反映种质资源表型遗传性状,分子标记则反映种质间遗传差异和亲缘关系[12],利用表型性状构建核心种质是较为传统的方法,得到广泛学者的认可[3-10]。有较多学者基于表型性状构建林木核心种质,如灰楸(CatalpafargesiiBur.)[13]、尾叶桉(EucalyptusurophyllaS.T.Blake)[4]、日本柳杉[Cryptomeriajaponica(L.f.) D.Don][14]、西伯利亚杏(Armeniacasibirica)[5]、核桃(JuglansregiaL.)[15]、水青树(TetracentronsinenseOliv.)[16]等。
云南松(PinusyunnanensisFranch.),又称“飞松”、“青松”、“长毛松”,为松科松属的常绿乔木,其生长迅速、耐干旱、耐瘠薄等优质特点[17-18],是西南地区荒山造林的先锋树种[19],是主要用材造林树种[17],也是治理水土流失的重要树种[20],同时具有经济效益、生态效益等重要作用[17]。云南松林是云南省的主要森林植被类型之一,主要分布在中国西南地区,如云南、贵州、四川以及西藏、广西等地[21],云南松林占云南省林分总面积的19.63%[22]。目前,云南松出现林分衰退,林分中低矮、扭曲等不良个体的比例偏高,优良基因资源少[20,23]。由于云南松种质资源群体分布广泛,这对优良材用云南松种质资源选育与保护具有一定挑战。前人基于表型性状[23]、SRAP分子标记[24]构建材用云南松种质资源库,虽然其效果较好,但构建的材用云南松核心种质样株分布较为分散,不利于种质资源更新收集。因此,研究基于地理角度构建材用云南松核心种质的可行性,对于提高材用云南松种质采集、保存及更新的便利性和有效性,具有重要的实用价值。
该研究基于王晓丽等[23]研究结果基础上,以材用云南松分布区不同地理来源的26个天然居群为原种质,每个天然居群采集干形通直圆满30株样株表型数据,基于材用云南松18个表型性状构建其核心种质,参考Miyamoto等[14]利用地理角度的核心种质构建方法,以及改进的最小距离逐步取样构建方法,比较2种策略构建材用云南松核心种质的效果,以获得优质的材用云南松种质资源,解决材用云南松种质资源来源问题,可为造林提供优良种质。
1 材料和方法
1.1 材料来源及表型数据采集
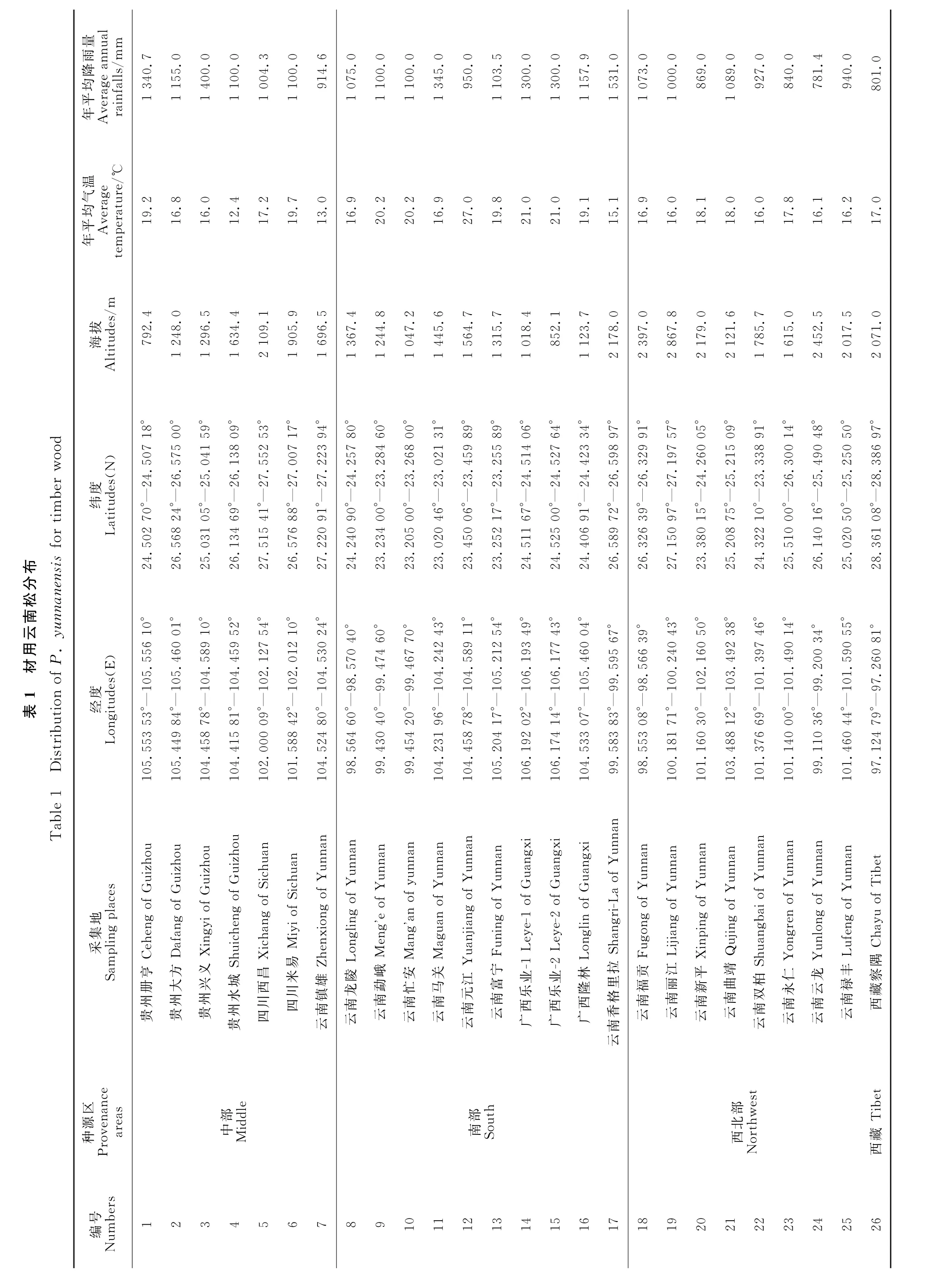
根据各地林业部门和相关研究学者提供的资料,选取26个具有材用代表性天然居群云南松种质资源的调查区域。于2016年12月至2017年2月完成26个居群780株云南松样株经纬度和海拔数据的收集(表1),每个天然居群采集30株样株,采集每株长势较为均匀的针叶束、种子、球果等数据。参考刘德浩等[3]和王晓丽等[23]关于表型数据标准化方法,对18个表型性状做10分级,1级≤μ-2δ,10级>μ+2δ,每级相差0.5δ,μ为各性状平均值,δ为各性状的标准差。
1.2 核心种质构建方法
1.2.1 地理角度构建方法
参考Miyamoto等[14]核心种质构建方法,采集云南松地理信息数据,并利用Arcgis做成空间散点图,将数据汇总于标准面积网格内,即第三方格(100 km×100 km)(图1,A)。根据第三方格实际情况变动具体距离值,合成第二方格(300 km×300 km)(图1,B)。计算每个第二方格从中心点到最近样株的距离为N(28 349.82~194 555.42 m)(图1,C),再计算所有第二方格中的N,并取它们的平均值,得到最近距离的平均值为100 144.61 m。以最近距离平均值为半径,在每个第二方格中画圆,选取所有在圆内的样株,组成种质子集,即可得到利用地理角度构建的种质子集(图1,D)。
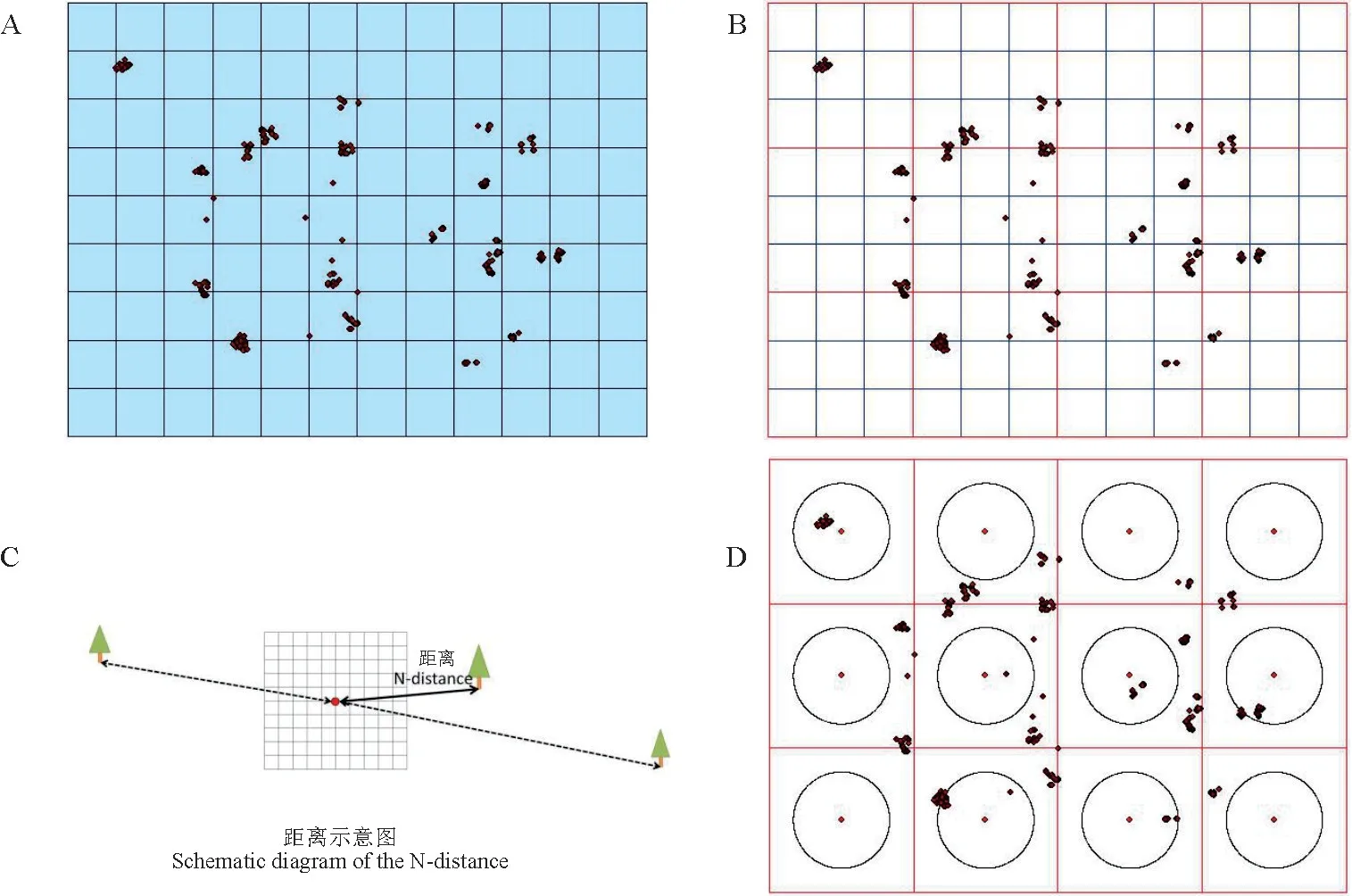
图1 种质子集构建方法A.第三方格;B.第二方格;C.中心点到样株距离;D.第二方格画圆。Fig.1 Construction method of core germplasmA.The third square.B.The second square.C.Distance from center point to sample plant.D.Drawing a circle for the second square.
1.2.2 改进的最小距离逐步取样构建方法(ILDSS)
先将原种质按种源分为4组,组内按照种源分为26个小组(表1),据此26个小组的780份样株设定4个抽样比例10%、20%、30%和40%,对18个表型性状标准化并用不加权类平均法(UPGMA)做聚类分析,遗传距离采用欧式距离,组内取样方法采用系统取样的多样性指数法,即计算4个组中各18个性状的遗传多样性指数,取此4组平均值作为该小组多样性指数,并计算每个小组的多样性指数占所有小组多样性指数的占比,并根据占比确定各小组抽样量。
然后计算原种质所有样株间的遗传距离,再根据遗传距离进行聚类,依据聚类结果找出遗传距离最小的一组,定向删除该组的1个样株,保留另1个样株进入下一轮聚类,如此循环直至保留样株达到固定抽样比例为止。
1.3 核心种质的评价及确认
利用4个评价指标(均值差异百分率、方差差异百分率、极差符合率和变异系数变化率)结合各性状的均值t检验、方差F检验、Shannon-Weaver遗传多样性指数、主成分分析方法综合考察构建的核心种质。Shannon-Weaver遗传多样性指数(H′)的计算参考郑福顺等[4]与陈存等[25]的方法,各表型性状频率的计算公式参考陈存等[25]的方法;按照Hu等[26]的计算方法,计算均值差异百分率(MD)、方差差异百分率(VD)、极差符合率(CR)和变异系数变化率(VR)4个指标评价核心种质的代表性,再根据Hu等[26]等提出初级核心种质的极差符合率(CR)大于80%和均值差异百分率(MD)小于20%这2个要求,当初步筛选的结果完全满足这2个指标,可认为对核心种质筛选出的样株能够较好地代表原种质[12-13,15]。
2 结果与分析
2.1 种质子集样株信息
2.1.1 地理角度抽样
种质子集筛选结果(图1,D),少部分样株在圆内,大部分样株散落在圆外(参考日本柳杉核心种质的确认方法),圆内为核心种质样株,经过筛选,从780株样株构成的原种质中选出219株样株组成种质子集,抽样比例为28%,种质子集样株具体来源为丽江30株,察隅30株,勐峨11株,忙安23株,马关30株,乐业-1 有30 株,乐业-2 有30 株,册亨5株,曲靖30株。
2.1.2 改进的最小距离逐步取样法
按照原种质10%、20%、30%、40%抽样比例进行核心种质筛选,样株数分别为78(每个小组有3株样株被选入种质子集)、156(每个小组有6株样株被选入种质子集)、234(每个小组有9株样株被选入种质子集)、312株(每个小组有12株样株被选入种质子集)。
2.2 种质子集代表性分析
2.2.1 不同构建方法获得的种质子集均值t 检验、方差F 检验
对原种质与种质子集各性状进行均值t检验、方差F检验,可知所构建的种质子集是否具有代表性,对不同构建方法及不同抽样比例所构建5个种质子集的18个数量性状均值t检验结果(表2)表明,2种构建方法所构建的种质子集与原种质均值间均无显著差异,从t检验结果来看,这5个种质子集均可代表原种质。
F检验结果(表2)表明:与原种质的18个性状相比,40%抽样比例构建的种质子集有11个性状与原种质具有显著性,30%抽样比例构建的种质子集有14个性状与原种质具有显著性,20%抽样比例构建的种质子集有15 个性状与原种质具有显著性,10%抽样比例构建的种质子集有12个性状与原种质具有显著性。地理角度抽样构建的种质子集F检验结果(表2)表明,有15个性状与原种质具有显著性,其种质子集性状显著性与原种质相比,呈现性状显著性数量越多,则说明其构建的种质获得了更大的变异,即构建的种质子集更具有代表性、异质性。因此,从均值t检验和方差F检验结果角度来看,从原种质种筛选出这5个种质子集均能够代表原种质。

2.2.2 原种质及不同构建策略种质子集的遗传多样性指数比较
原种质与不同构建策略构建的5个种质子集的18个性状遗传多样性指数分析结果(表3)表明,利用不同方法构建的5个种质子集中,20%抽样比例构建的种质子集多样性指数平均值最高,且总体多样性指数均优于其余4个种质子集多样性指数,远高于原种质。
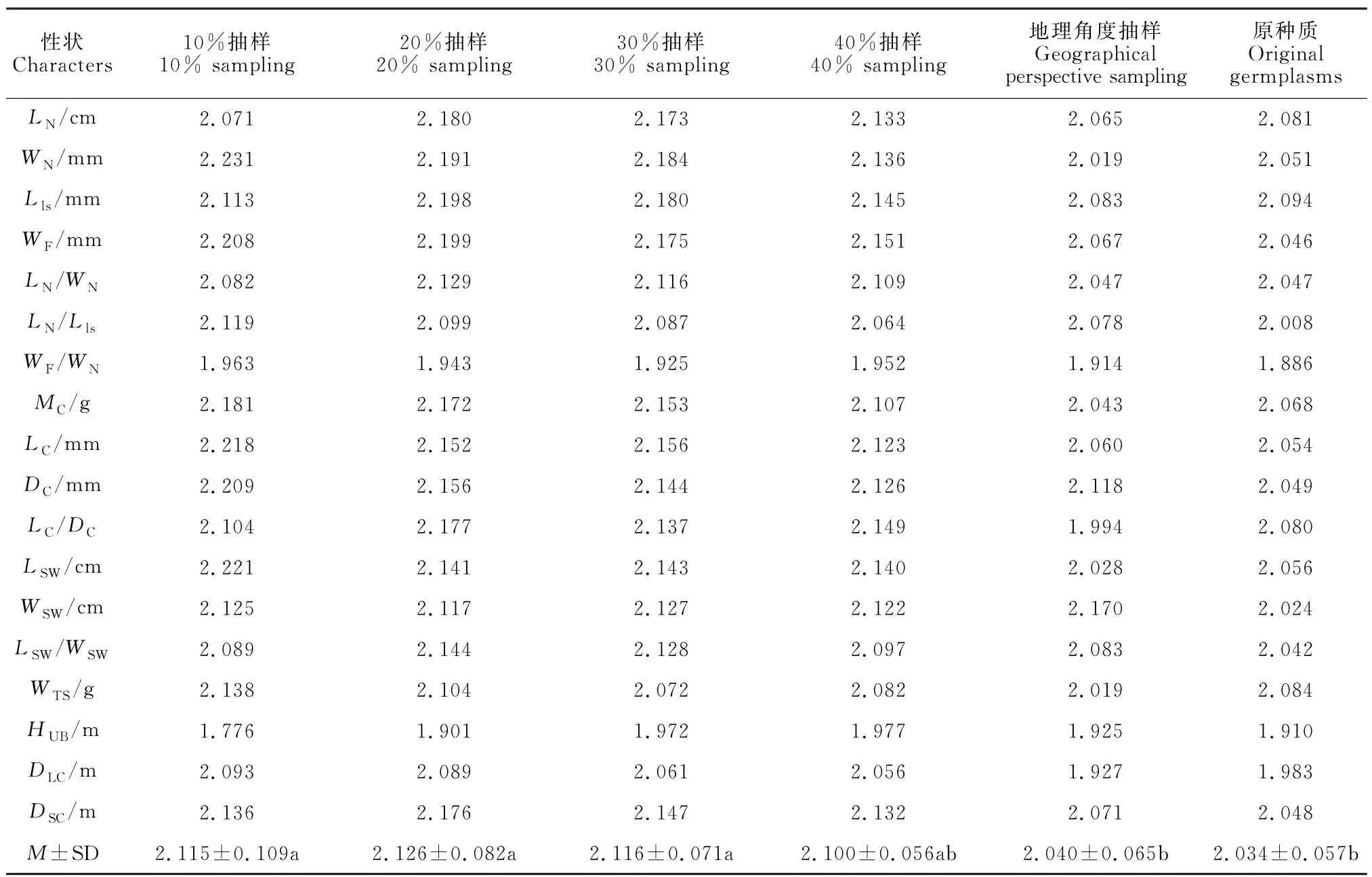
表3 不同构建策略种质子集遗传多样性指数比较Table 3 Comparison of genetic diversity index of germplasm subsets with different construction strategies
地理角度构建的种质子集的多样性指数总体略高于原种质,有8个性状多样性指数低于原种质(针叶长、针叶宽、叶鞘长、球果重量、球果长/球果直径、种翅长、千粒重、长冠径),总体多样性指数均低于其余4个种质子集。
对原种质与5个种质子集多样性指数进行方差分析和多重比较(表3)表明,原种质与5个种质子集均存在不同程度的差异,其中20%抽样比例的种质子集多样性指数极显著大于原种质,地理角度构建的种质子集多样性指数与原种质无显著差异。
综上所述,从多样性指数分析结果来看,地理角度构建的种质子集多样性指数低于20%抽样比例构建的种质子集,这2种构建策略构建的种质子集均能代表原种质,但20%抽样比例构建的种质子集能更好代表原种质的遗传多样性。
2.2.3 不同构建策略种质子集的评价指标分析
原种质与不同构建策略构建的5个种质子集的评价指标分析结果(表4)表明,5个种质子集均值差异百分率均小于20%,极差符合率均大于80%,这表明5个种质子集均符合构建要求。地理角度抽样的种质子集均值差异百分率最小,但变异系数变化率和极差符合率均低于其余4个抽样的种质子集,而方差差异百分率与20%抽样的种质子集相同且高于其余3个种质子集。改进的最小距离逐步取样法的4个种质子集中,20%抽样比例的种质子集均值差异百分率较小,变异系数变化率、方差差异百分率最大,极差符合率较大,且种质子集样株较少,这利于核心资源保存,表明20%抽样比例构建的种质子集比其余3个抽样比的种质更具有效性、实用性和代表性。
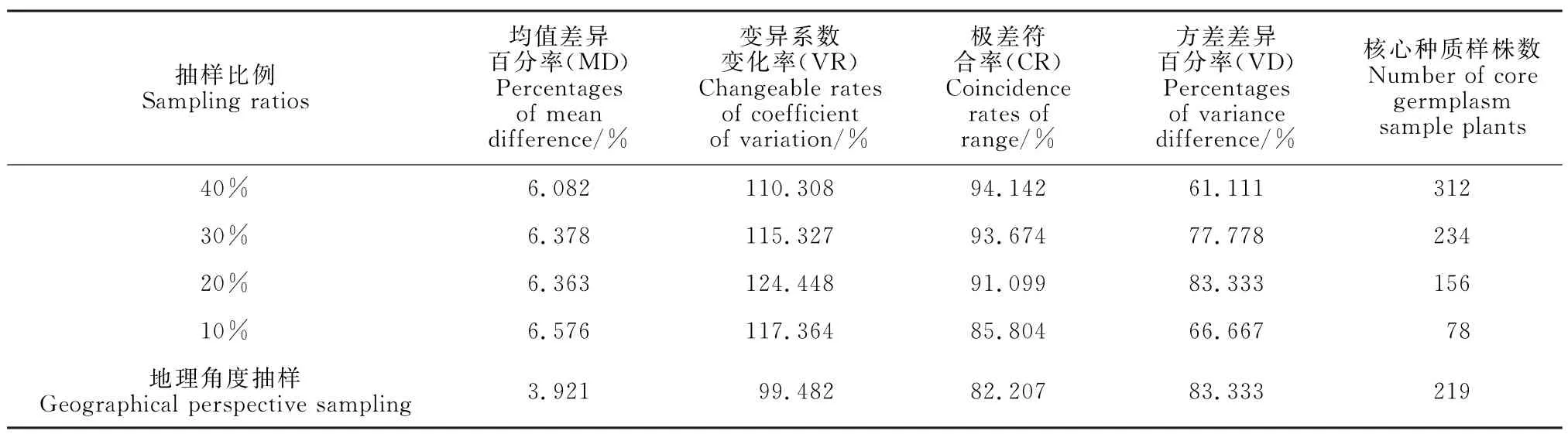
表4 不同构建策略种质子集评价指标比较Table 4 Comparison of evaluation indexes of germplasm subset with different construction strategies
综合分析表明20%抽样比例与地理角度抽样构建的核心种质存在不同程度的优势,均可代表原种质。
2.3 核心种质的确认
2.3.1 原种质与核心种质生长指标主成分分析
分别对原种质及不同方法构建的2个核心种质的18个表型性状进行主成分分析(表5),原种质、20%抽样与地理角度抽样均提取7个特征值大于1的主成分,原种质、20%抽样比例与地理角度抽样7个主成分累计贡献率分别为79.376%、83.539%、82.163%。20%抽样比例与地理角度抽样的核心种质主成分累计贡献率大于原种质,且接近85%,表明20%抽样比例与地理角度抽样的核心种质能够解释的表型遗传信息大于原种质,去除了原种质中大量重复性的种质资源,减少了遗传冗余,因此可以认为20%抽样比例与地理角度抽样的核心种质具有较强的代表性和实用性。

表5 原种质与核心种质主成分分析Table 5 Principal component analysis of original germplasm and core germplasm
2.3.2 原种质与核心种质样株分布主成分分析
将780株原种质样株与核心种质样株的经纬度利用工具转化为度的数字格式后分别做散点图进行分析,横纵坐标分别为PC1和PC2(图2)。20%抽样比例筛选的156株的核心种质散点图显示,云南松核心种质的分布包含原种质的整个分布范围及外围具有特异性征的样株;同时,具有较高遗传相似性的样株得以有效保留,确保核心种质的代表性、实用性,从而更有效地保留核心种质与原种质的表型遗传和变异。地理角度抽样筛选的219株核心种质分布比较集中,基本包含了不同气候、海拔条件下云南松种质资源,同时方便收集核心种质资源。
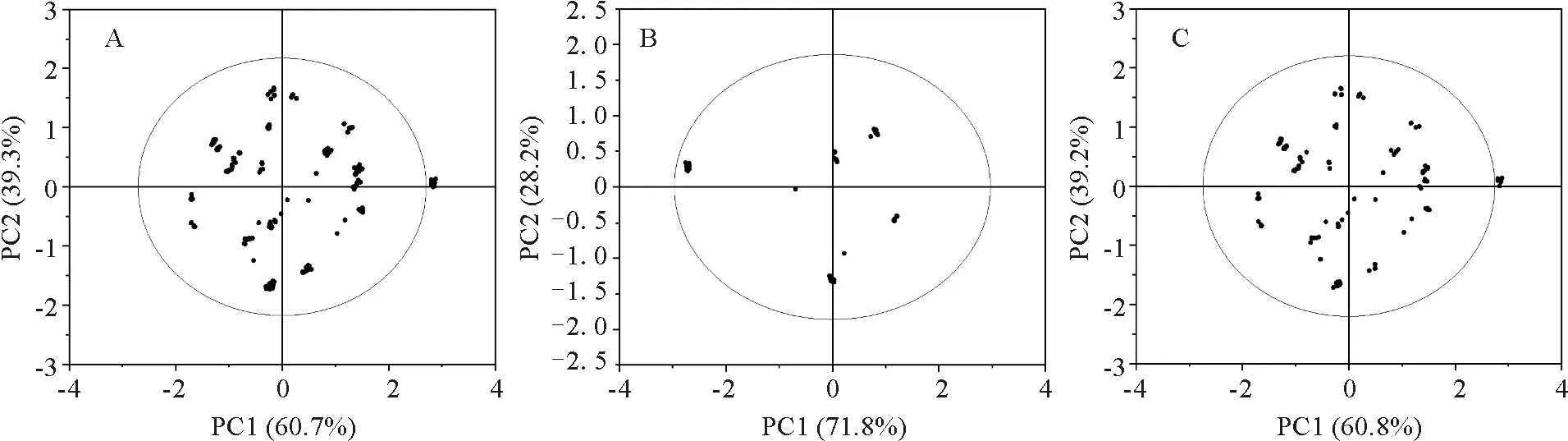
图2 原种质与核心种质样株分布主成分分析A.原种质样株分布散点图;B.地理角度样株分布散点图;C.20%抽样比例样株分布散点图。Fig.2 Principal component analysis of the distribution of the original and core germplasm samplesA.Scatter plot of original germplasm.B.Scatter plot of sample plant distribution from geographical perspective.C.Scatter plot of 20% sampling proportion.
3 讨论
3.1 基于表型性状构建林木核心种质的抽样策略
取样策略是利用地理角度构建核心种质的重要步骤。因此,选择合适的核心种质构建策略尤为重要。部分学者提出利用GIS系统构建核心种质,如Jarvis等[27]研究表明GIS 系统分析的信息有助于高效地保护和利用遗传多样性;随后Gepts等[28]提出利用GIS系统与植物的遗传多样性促进种质资源的保护和利用;Miyamoto等[14]利用基于地理、遗传和环境空间3个角度构建柳杉核心种质,该研究表明以平均值为半径的圆中构建核心种质资源的方法是可取的。因此,本研究参考Miyamoto等[14]构建方法,探究基于表型性状利用地理角度抽样构建材用云南松核心种质的合理性,利用遗传多样性指数、评价指标及主成分分析等常规核心种质评价方法。结果表明,该核心种质与Miyamoto等[14]研究结果相似,样株分布较为集中,能够代表原种质的表型遗传多样性,其各项分析均符合核心种质构建要求。
郑福顺等[4]和于晓池等[13]表明核心种质构建方法的重点是对种质合理分组及取样策略,不同遗传距离也会影响聚类结果。因此,利用改进的最小距离逐步取样法中首要问题是种质资源的分组及抽样策略。前人研究表明,地理分组是常见的分组方法,如孙永强等[5]和陈存等[25]表明基于种源地理信息结合种质表型遗传特征对原始种质进行分组构建的核心种质更具代表性。通常采用不同聚类方法结合不同取样方法[3-6,23],前人研究[3,23,29]表明,改进的最小距离逐步取样是聚类结束后,删除该组内遗传距离最小的样株,故改进的最小距离逐步取样法比最小距离逐步取样、随机取样的方法效果更佳。彭枫等[8]和于晓池等[13]研究表明遗传距离是度量样本间遗传相似性的综合数量指标,常用遗传距离为欧式距离和马氏距离[5],而欧式距离是基于表型数量性状,不受量纲影响,故多项研究表明欧式距离优于马氏距离[5,8,13]。本研究在汪磊等[7]和王晓丽等[23]研究结果上,依据材用云南松地理种群特征进行种质分组,组内取样量采用多样性指数法,遗传距离采用欧式距离,采用不加权类平均法进行聚类分析,利用改进的最小距离逐步取样法筛选种质子集样株,结果显示,该方法最佳抽样为20%的种质子集,该种质子集优于其余4个种质子集,更符合核心种质构建的要求,还提高了原种质的变异系数,且种质子集样株较少,这与刘德浩等[3]、汪磊等[7]、王晓丽等[23]和章秋平等[29]的研究结果一致。
3.2 基于表型性状构建林木核心种质的取样规模
大多学者认为抽样规模是核心种质构建重要的一部分,如果取样规模较少,这会导致构建的种质集缺乏异质性、代表性;如果取样规模过多,又不能较好地去除遗传冗余[15]。陈存等[25]认为木本植物的取样比例则在10%~45%之间,而大部分学者认为10%~30%取样比例效果比较好[30-32]。彭枫等[8]结果表明以菠菜原种质15%取样比例最佳,侯志强等[6]采用20%取样比例菊芋构建核心种质最优,张欢等[16]确定45%取样比例适宜水青树核心构建,曾钦朦等[15]以50%抽样比例构建贵州核桃核心种质资源。本研究利用地理角度抽样与改进的最小距离逐步取样法构建不同抽样比例种质子集,其构建的种质子集抽样占比分别为28%和20%,本研究构建的材用云南松核心种质取样比例均符合大多数学者的取样比例。
3.3 基于表型性状构建林木核心种质的代表性评价
对原种质与构建的种质子集进行均值t检验、方差F检验能初步了解构建的种质子集是否具有异质性、代表性。t检验结果表明,2种构建策略所构建的种质子集与原种质均值间均无显著差异,说明两者在表型性状上频率相近,故从t检验结果角度看,这5个种质子集均可代表原种质,这与孙永强等[5]、曾钦朦等[15]和赵立民等[31]构建核心种质结果相似。方差F检验结果表明,改进的最小距离逐步取样法选取10%、20%、30%、40%抽样,分别有12,15,14,11个性状与原种质18个性状有显著差异;地理角度抽样有15个性状与原种质18个性状有显著差异。因此,这5个抽样比例均能够更好地代表原种质的表型遗传多样性,也具有较强异质性,不同构建策略的F检验结果与孙永强等[5]和章秋平等[29]的研究结果相似,而与赵立民等[31]研究结果表明,对13个性状进行F检验仅3个性状呈差异显著,这说明本研究构建的种质子集更具有异质性。
遗传多样性指数反映种质表型多样性特征,表现原种质与核心种质各表型性状频率分布特点[12]。多样性指数越小,反映构建的种质子集中有个别性状占比较大,即核心种质表型性状间等级分布不均匀;反之,多样性指数越大,则表明构建的种质子集的表型性状等级分布均匀。因此,地理角度构建的种质子集中性状等级主要分布于平均值附近,其遗传多样性指数总体略高于原种质,保留原种质基本表型性状变异,与原种质无明显差异;而基于改进的最小距离逐步取样法构建的4个种质子集中性状等级分布均匀,其中20%抽样比例的多样性指数高于其余3个种质,且高于地理角度的多样性指数,显著大于原种质,获得了表型性状更均匀的种质子集。
大多数学者对核心种质评价常采用Hu 等[26]提出的4个评价指标,即均值差异百分率(MD)、方差差异百分率(VD)、极差符合率(CR)和变异系数变化率(VR);核心种质满足CR 大于80%和MD小于20%这2 个要求时,当MD 越小,VD、CR 和VR 越大,表明其代表性和多样性越好[12,25,32]。本研究利用改进的最小距离逐步取样法选取的4个抽样中,20%抽样比例MD较小,为6.363%,VR最大,为124.448%,VD最大,为83.333%,CR为91.099%,种质子集样株相比其余3个种质而言,能够更好地代表原种质的表型遗传多样性。而地理角度抽样MD 最小,为3.921%,VR 最小,为99.482%,CR 最小,为82.207%,VD 最大,为83.333%,符合构建要求,但该种质子集在构建效果上劣于20%抽样比例构建的种质子集。综合分析不同构建策略构建的种质子集,20%抽样较地理角度抽样获得更大的变异,均符合核心种质构建的要求,这与刘德浩等[3]、彭枫等[8]、陈存等[25]的研究结果相似,且符合Hu等[26]提出的评价指标要求。
主成分分析用于比较原种质于核心种质的表型性状的分布特征[12]。对原种质、20%抽样比例和地理角度抽样种质子集的表型性状均提取了特征值大于1的7个主成分,其累计贡献率分别为79.376%、83.539%、82.163%,这表明不同策略构建的2个核心种质均能够解释表型遗传信息大于原种质。20%抽样比例的156株材用云南松核心种质样株分布散点图显示,该核心种质包含了原种质的整个分布范围,降低了原种质重叠部分,从而去除了遗传冗余,这与孙永强等[5]、曾钦朦等[15]和陈存等[25]的研究结果一致;而地理角度构建219株材用云南松核心种质样株较为集中的分布在不同经度、纬度的气候性地带,基本包含了不同气候、海拔条件下云南松种质资源。
4 结论
研究基于780份材用云南松18个表型性状,利用地理角度以抽样比例为28%筛选219株材用云南松核心种质;改进的最小距离逐步取样法筛选出最佳抽样为20%的156株样株。t检验表明,这2种构建策略筛选的核心种质与原种质无显著差异;F检验结果表明,2个核心种质均有15个性状呈显著差异,具有较好异质性;20%抽样比例的遗传多样性指数高于地理角度的多样性指数,显著大于原种质,获得了表型性状更均匀的种质子集,而地理角度多样性指数与原种质无显著差异,这2个核心种质解释的表型遗传信息大于原种质,20%抽样比例的核心种质样株包含原种质的整个分布范围,而地理角度抽样核心种质分布比较集中。因此,对这2种构建效果综合评价与验证表明,2种策略构建结果均可代表材用云南松原种质的表型遗传多样性,但地理角度构建的种质子集更便于收集种质资源,在种质资源保存和更新方面更具有优势,减少了成本,可为材用云南松种质资源保存和优良种质选育提供科学方法,同时也为其他种质资源构建提供一种思路。
