盐胁迫下基于TMT技术的罗布麻蛋白质组学分析
2024-03-08刘旭洋张晓明
甄 熙,刘旭洋,张晓明,宛 涛*
(1 内蒙古农业大学 草原与资源环境学院,呼和浩特 010019;2 内蒙古自治区人工影响天气中心,呼和浩特 010051;3 内蒙古自治区气候中心,呼和浩特 010051)
蛋白质组学是研究整个生物体内全套的蛋白质组及其动态变化规律的科学[1-2],植物蛋白质组学主要通过对植物在特定条件下蛋白质表达的种类、水平以及修饰状态进行鉴定,解析蛋白质功能和蛋白互作的机制[3]。目前,定量蛋白质组学检测方法主要有2种:iTRAQ(isobaric tags for relative and absolute quantitation)和TMT(tandem mass tags)[4]。李俊良用iTRAQ 方法对甜菜(Betavulgaris)在盐胁迫下的蛋白质表达进行分析,发现差异蛋白主要通过参与渗透调节、光合作用和能量代谢等过程抵抗盐胁迫[5]。Chen等对耐盐和盐敏感2个玉米品种进行盐胁迫处理,通过iTRAQ 方法共鉴定出1 056种差异表达蛋白,差异蛋白主要与苯丙素生物合成、淀粉和蔗糖代谢以及丝裂原活化蛋白激酶(MAPK)信号通路相关[6]。有人用TMT 技术分析了金银花(Lonicerajaponica)的耐盐性,发现苯丙素类生物合成和单萜类生物合成途径的相关基因上调表达,差异蛋白主要参与能量代谢和碳水化合物代谢过程途径以增强耐盐性[7]。
近年来对罗布麻(ApocynumvenetumL.)的相关研究主要集中在药用成分、遗传多样性和盐胁迫耐受性等方面[8-10]。对于罗布麻在盐胁迫条件下的适应机制研究大多数都集中在生理和表型方面,也有一些研究试图从分子机制方面阐明罗布麻的耐盐机制。基于转录组水平研究发现黄酮类物质与罗布麻提高耐盐性有关,由于黄酮类化合物的积累,转基因拟南芥比野生型拟南芥表现出更高的耐盐性[10]。罗布麻DEAD-box解旋酶AvDH1基因在棉花基因组中稳定遗传,经耐盐性鉴定发现转基因棉花株系的耐性显著提高[11]。罗布麻黄酮醇合成酶基因AvFLS过表达能够提高拟南芥的黄酮类化合物含量和光合速率,降低丙二醛含量,从而显著提高转基因拟南芥的耐盐能力[12]。综上所述,国内外学者在罗布麻药用成分、遗传多样性及耐盐性等方面开展了大量研究工作,但对野生罗布麻耐盐性机理的关注较少,从而限制了对其种质资源的保护和发掘利用。
罗布麻虽属于耐盐性较强的植物,但随着生境遭到破坏及土壤盐渍化程度加重,罗布麻的野生种群数量逐渐缩减,探究罗布麻的耐盐性机制,对罗布麻野生种质资源的保护和利用具有重要意义。前期研究发现罗布麻对土壤盐渍化具有较强的适应能力,研究拟通过盐胁迫下罗布麻蛋白组学分析探究其对盐胁迫的应答机制,以期揭示罗布麻耐盐的分子机制并筛选出罗布麻响应盐胁迫的关键蛋白,拓展罗布麻盐胁迫应答相关的分子资源,并为罗布麻天然抗逆种质的发掘与利用提供试验与理论依据。
1 材料和方法
1.1 试验材料
材料为罗布麻野生植株,取样地点在内蒙古杭锦旗罗布麻样地(东经108°44′44″,北纬40°35′23″),该样地土壤含盐量较高且有较多1 年生罗布麻幼苗,便于筛选长势均匀的植株。在野外样地,选择植株高度、茎干粗细一致的罗布麻幼苗,放入Hoaglang营养液中带回实验室培养。
选取长势良好的野生罗布麻植株,设置盐胁迫处理组与对照组2个组别,每个处理3次重复。根据前期盐胁迫下生理指标测定结果,选取300 mmol/L为蛋白组试验的胁迫浓度,该胁迫浓度下罗布麻生理代谢活动最强,渗透调节物质含量及抗氧化酶活性最高,表现出较强的渗透调节及代谢活性。操作过程:将试验组罗布麻植株放于纯水中小心清洗后移入配制好的胁迫营养液(含300 mmol/L NaCl);对照组植株清洗后移入另一不含NaCl的营养液中。分别胁迫0 h、2 h、6 h、12 h、24 h和48 h,取罗布麻叶片迅速放入冻存管中进行液氮深冻,然后将样品放入-80 ℃冰箱中保存备用。
1.2 蛋白提取和定量
样品加入1%的PVPP和适量BPP溶液,使用高通量组织研磨仪振荡3 次,每次40 s;4 ℃下12 000g离心20 min,取上清液,加入等体积Tris-饱和酚,4 ℃旋涡振荡10 min;4 ℃下12 000g离心20 min,取酚相,加入等体积BPP溶液,4 ℃旋涡振荡10 min;4 ℃下12 000g离心20 min,取酚相,加入5倍体积预冷的乙酸铵甲醇溶液,-20 ℃沉淀过夜;次日4 ℃下12 000g离心20 min,弃上清液,向沉淀中加入90%预冷丙酮混匀后离心弃上清液,重复2次;沉淀用蛋白裂解液(8 mol/L 尿素+1%SDS,含蛋白酶抑制剂cocktail)溶解,冰上超声2 min;4 ℃ 12 000g离心20 min,取蛋白上清液;BCA 法测定蛋白浓度,SDS-PAGE 电泳。用Thermo Scientific Pierce BCA 试剂盒进行蛋白定量。用BCA 试剂盒配制BCA 工作液和不同质量浓度的标准蛋白质溶液,其质量浓度分别为0,0.125,0.250,0.500,0.750,1.000,1.500,2.000 mg/mL。各样品均取2 μL和18 μL水混合,加入200 μL BCA 工作液。振荡混匀,37 ℃反应30 min,在562 nm 处读取吸光度。
1.3 酶解烷基化以及标记
取蛋白样品100 μg,用裂解液补充体积到90 μL,加入终浓度10 mmol/L TCEP还原剂,在37 ℃下反应60 min;加入终浓度40 mmol/L 碘乙酰胺,室温下避光反应40 min。每管各加入预冷的丙酮(V丙酮∶V样品=6∶1),-20 ℃沉淀4 h,10 000g离心20 min,取沉淀。用50 mmol/L TEAB充分溶解样品,按照质量比1∶50(酶∶蛋白)加入Trypsin在37 ℃酶解过夜。
TMT 标记与混样,-20 ℃取出TMT 试剂(Thermofisher 90111)恢复到室温,加入乙腈,涡旋离心,每100 μg多肽加入1管TMT 试剂(TMT10-126标记,127N 标记A2,127C 标记A3,128N 标 记B1)。室温孵育2 h;加入羟胺,室温反应15 min,将等量标记产物混合于一管中,置于真空浓缩仪浓缩处理,样品冷冻保存备用。
1.4 RPLC一维分离
用UPLC 上样缓冲液复溶多肽样品,用反相C18柱进行高pH 液相分离。A 相2%乙腈(氨水调pH 至10),B相80%乙腈(氨水调pH 至10),0~2 min,100% A;2~17 min,0~3.8% B;17~35 min,3.8%~24% B;35~38 min,24%~30% B;38~39 min,30%~43% B;39~40 min,43%~100% B;40~46 min,100%~0 B。紫外检测波长为214 nm,体积流量为200 μL/min,洗脱时间为66 min。根据峰形和时间共收取20个馏份,合并成10个馏份,真空离心浓缩。
1.5 液相串联质谱
第二维采用纳升级液相色谱串联质谱技术(Easy-nLC 1200 结合Q Exactive质谱仪)进行分析。肽段用质谱上样缓冲液溶解,上样后经C18色谱柱(75 μm×25 cm,Thermo,USA)分离120 min,体积流量为300 μL/min。EASY-nLC 液相梯度洗脱,A 相2%乙腈(加0.1%甲酸),B相80%乙腈(加0.1%甲酸),0~1 min,0~5% B;1~63 min,5%~23% B;63~88 min,23%~48% B;88~89 min,48%~100% B;89~95 min,100% B。
MS和MS/MS 采集之间自动切换,质谱分辨率分别是70 K 和35 K。MS进行全扫(350~1 300 m/z),选择母离子top20进行二级碎裂,动态排除时间18 s。
1.6 蛋白搜库和差异蛋白筛选
采用Protein Pilot Software 5.0(AB Sciex,美国)和Swiss Prot/UniProt数据库进行蛋白鉴定。对质谱下机的原始数据经Proteome DiscovererTM Software 2.2(Thermo Fisher,美国)进行处理。搜库过程中肽段鉴定的错误发现率(FDR)设置为FDR≤0.01。蛋白至少含有1个特异肽段。
项目共检测到12 457个蛋白,使用R 语言计算样本间差异显著性P值,同时计算组间差异倍数(αFC)。显著差异表达蛋白的筛选标准为:P<0.05且αFC>1.2为上调蛋白,P<0.05且αFC<0.83为下调蛋白。
1.7 生物信息学分析
选择GO(gene ontology,http://www.blast2go.com/b2ghome;http://geneontology.org/)对所有的差异蛋白进行功能聚类分析,用KEGG(kyoto encyclopedia of gene and genomes,http://www.genome.jp/kegg/pathway.html)通路数据库对差异蛋白涉及的代谢通路进行分析。用STRING 和Cytoscape软件构建蛋白质相互作用网络。
1.8 平行反应监测(PRM)分析
在TMT 数据中筛选出与响应盐胁迫相关的8个差异表达蛋白进行表达量验证。用平行反应监测(parallel reaction monitoring,PRM)方法对其表达水平进行验证分析。从样品中提取蛋白后,对质量达标的蛋白样品进行还原烷基化处理。样品经过胰蛋白酶酶解纯化后,分别用FULL MS/DD-MS2(TOP20)和FULL MS-PRM 2种方法进行质谱分析和PRM 蛋白鉴定质谱检测。最后,分析质谱采集数据,确认目标蛋白的目标肽段进行后续定量可行性分析。液相色谱、扫描范围和质谱分辨率等参数与TMT 蛋白质组测序的条件相同。
2 结果与分析
2.1 盐胁迫下罗布麻叶片蛋白提取检测结果
对罗布麻叶片盐胁迫样品进行蛋白质定量测序,共得到569 883张图谱,用转录组建立的蛋白组数据库对原始质谱数据进行搜库,鉴定到59 274个肽段,12 457 个蛋白。蛋白质提取检测,肽段长度范围在5~20个氨基酸之内(图1,A、B),表明酶解充分且后续鉴定结果可靠。氨基酸个数从1~5到41~60之间蛋白覆盖度都高于15%,高于15%的蛋白覆盖度达到91.61%(图1,C、D),说明鉴定结果可靠。

图1 蛋白的鉴定与分析A.肽段数量分布;B.肽段长度分布;C.蛋白分子量分布;D.蛋白覆盖度分布。Fig.1 Analysis and identification of proteinsA.Peptide number distribution.B.Peptide length distribution.C.Molecular weight distribution of the identified proteins.D.Distribution of protein sequence coverage.
2.2 盐胁迫下罗布麻叶片差异表达蛋白分析
对罗布麻在盐胁迫的不同时间鉴定到不同数量和不同种类的差异表达蛋白(differentially expressed proteins,DEPs),T2 vs T0 组差异蛋白424,其中上调蛋白281,下调蛋白143;T6 vs T0差异蛋白645,上调485,下调160;T12 vs T0共检测到791个差异蛋白,上调蛋白409,下调蛋白382;T24 vs T0检测到1 001 个差异蛋白,上调546,下调455;T48 vs T0差异蛋白1 227,上调576,下调651。以上结果(图2)表明,在罗布麻响应盐胁迫的过程中,随时间延长有更多差异表达蛋白参与胁迫过程。

图2 差异表达蛋白(DEPs)的火山图红色代表上调蛋白,蓝色代表下调蛋白,灰色代表非显著差异的蛋白。Fig.2 Volcano plot of DEPsRed points () represent up-regulated proteins,blue ones () represent down-regulated proteins,and unchanged proteins are gray points ().
2.3 盐胁迫下罗布麻叶片差异蛋白的GO 分析
为深入了解差异蛋白的功能,对罗布麻盐胁迫2 h、6 h、12 h、24 h、48 h与对照0 h对比获得的差异蛋白进行GO 分析(表1)。差异蛋白GO 功能注释主要包括生物学过程(biological process,GOBP)、细胞成分(cellular component,GO-CC)、分子功能(molecular function,GO-MF)。在T2 vs T0比较组(图3,A)中共鉴定到424个差异表达蛋白,有83,13,4,3个差异表达蛋白分别富集在氧化还原酶活性、钙离子结合、肉桂醇脱氢酶活性和茉莉酸代谢过程,显著富集的主要有叶绿素代谢过程、细胞壁修饰、酸性磷酸酶活性、天冬氨酸酯酶活性、氧化还原酶活性等。T6 vs T0组鉴定到的差异蛋白中,生物学过程(GO-BP)主要富集在:核糖核蛋白复合物生物发生、细胞成分生物发生、肽生物合成过程、细胞质翻译等;细胞成分(GO-CC)主要富集在胞质大核糖体亚基、胞质小核糖体亚基、类核等;分子功能(GO-MF)主要富集在rRNA 结合、核糖体的结构成分、结构分子活性(图3,B)。T12 vs T0组筛选出的差异蛋白中分别有128,5,3,3个差异蛋白富集在氧化还原酶活性、乙酰辅酶A 生物合成过程、维生素E生物合成过程和异柠檬酸脱氢酶活性,富集显著性最高的主要为细胞酰胺代谢过程、胞质大核糖体、rRNA 结合等(图3,C)。T24 vs T0组差异蛋白富集最多的为氧化还原酶活性通路共有163个差异表达蛋白,占这一过程中差异蛋白总数的16.3%,其余富集显著性较高的有脂肪酸代谢过程、碳水化合物代谢过程、多糖代谢过程、细胞壁、水解酶活性等(图3,D)。T48 vs T0组有195个差异蛋白富集在氧化还原酶活性通路,其余富集显著性最高的有内肽酶调节活性、细胞壁、碳水化合物代谢过程等(图3,E)。

图3 盐胁迫下罗布麻差异表达蛋白的GO 富集分析P<0.001标记为***,P<0.01标记为**,P<0.05标记为*。下同。Fig.3 GO of DEPs in A.venetum under salt stressP<0.001 marks as ***,P<0.01 marks as **,P<0.05 marks as *.The same as below.
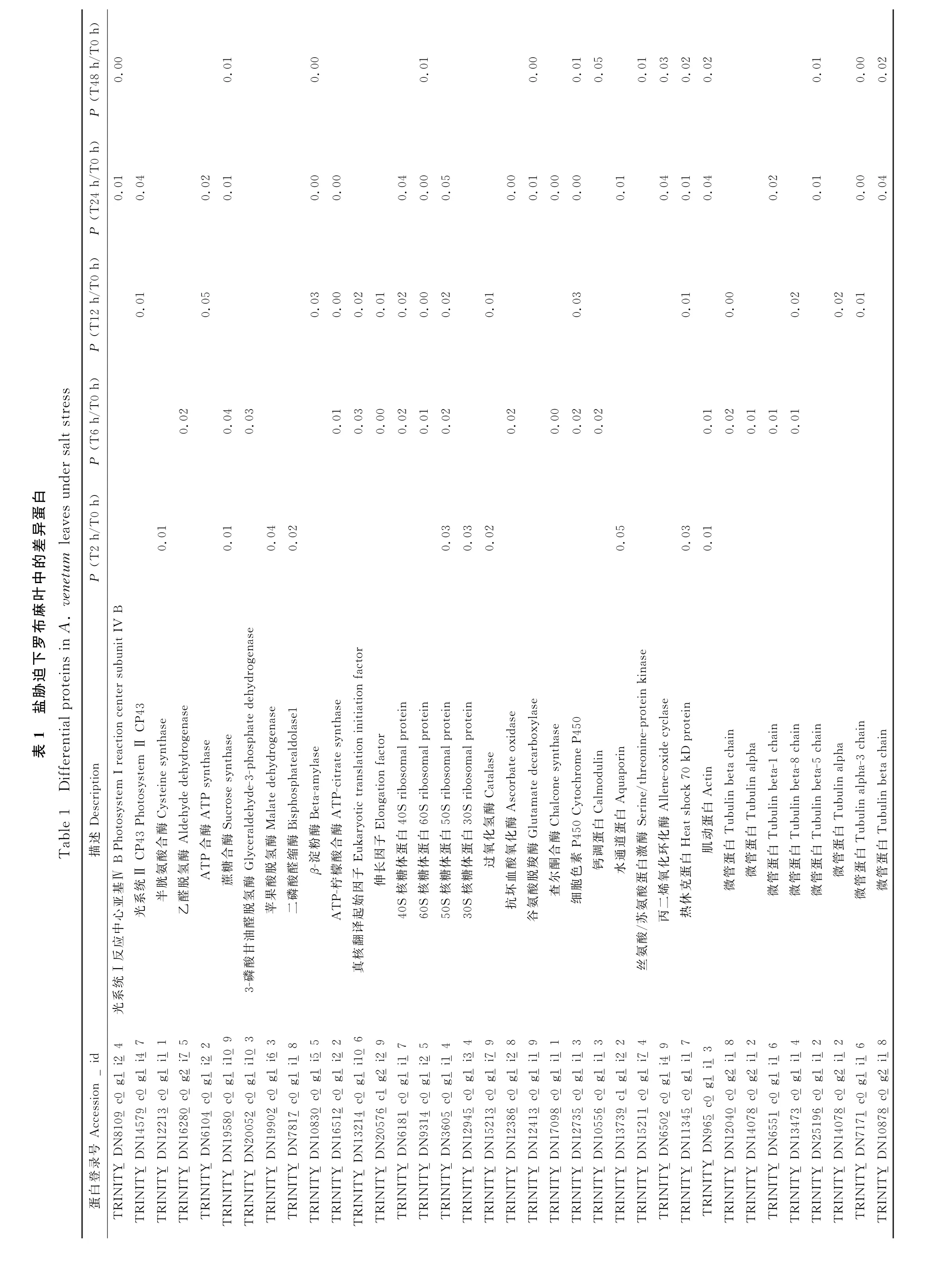
2.4 罗布麻叶应答盐胁迫差异蛋白的KEGG 分析
KEGG 分析表明,在罗布麻盐胁迫2 h、6 h、12 h、24 h和48 h与对照0 h相比,分别有82,100,99,112,111个差异表达蛋白参与了代谢途径,参与最多的为碳水化合物代谢、氨基酸代谢和翻译通路(图4)。可见罗布麻盐胁迫过程中,随着胁迫时间的延长,差异表达蛋白参与了更多代谢通路。

图4 盐胁迫下罗布麻差异表达蛋白质的KEGG 代谢通路分析Fig.4 KEGG pathway of DEPs in A.venetum under salt stress
T2 vs T0组差异蛋白主要富集在:卟啉和叶绿素代谢、鞘脂代谢、亚油酸代谢。T6 vs T0组差异蛋白主要富集在:核糖体和黄酮类生物合成、丙酮酸代谢、柠檬酸循环(TCA循环)。T12 vs T0组差异蛋白主要富集在:核糖体和黄酮类生物合成、苯丙烷生物合成、淀粉和蔗糖代谢。T24 vs T0组差异蛋白主要富集在:黄酮类生物合成,淀粉和蔗糖代谢,脂肪酸生物合成,甘氨酸、丝氨酸和苏氨酸代谢。T48 vs T0组差异蛋白主要富集在:苯丙烷生物合成、淀粉和蔗糖代谢、MAPK信号通路、戊糖和葡萄糖醛酸的相互转化。见图5。
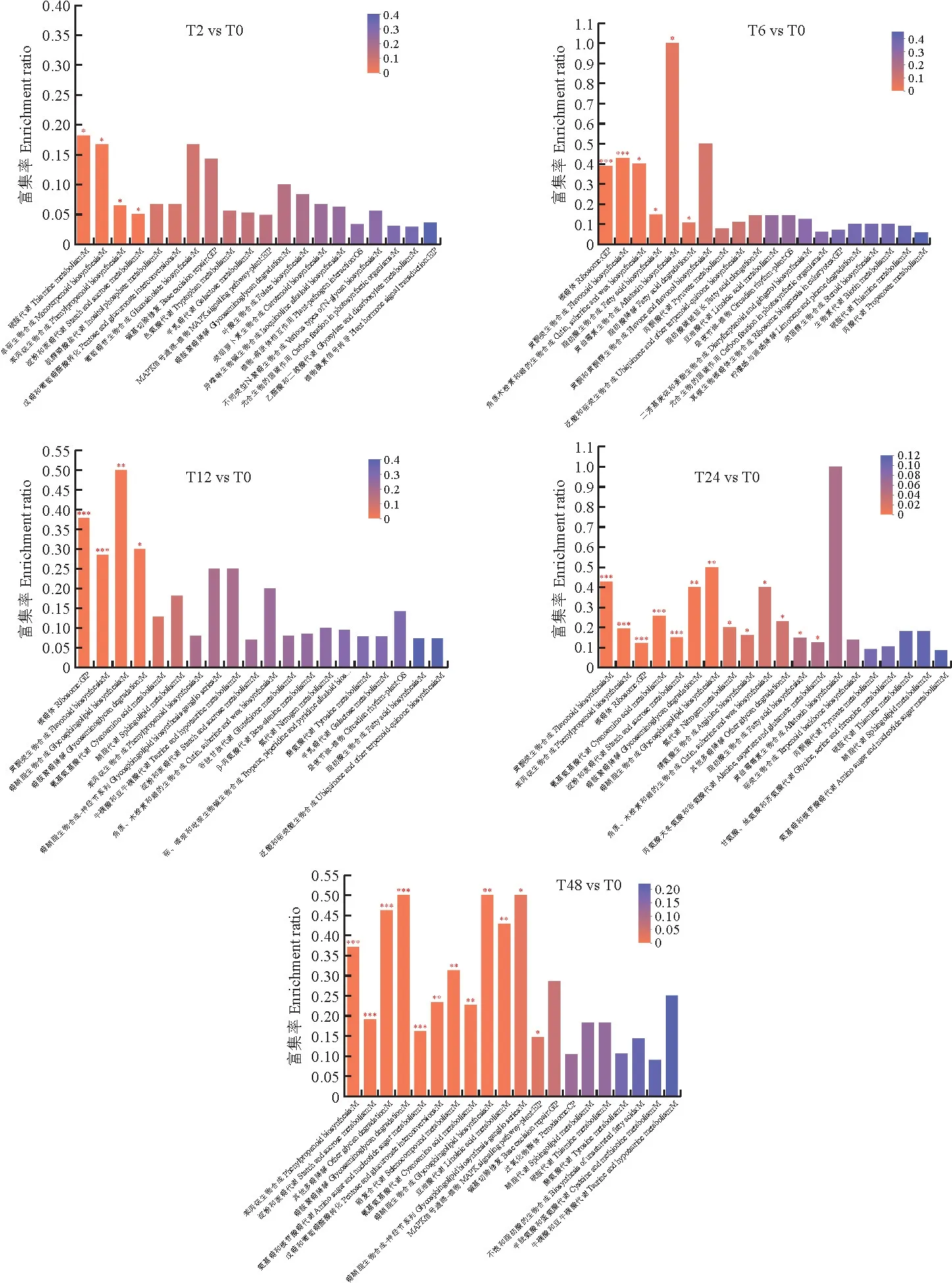
图5 盐胁迫下罗布麻差异蛋白的KEGG 富集Fig.5 KEGG enrichment of DEPs in A.venetum under salt stress
2.5 盐胁迫下罗布麻叶片共有的差异蛋白分析
共有差异蛋白的韦恩图分析表明,有11个蛋白在2 h、6 h、12 h、24 h、48 h罗布麻响应盐胁迫过程中差异表达(图6)。其中,有5个蛋白下调表达,5个蛋白上调表达,另外有1个蛋白在2 h时表达量增加,之后表达量降低(图7)。进一步进行GO 富集分析发现,这些共有差异蛋白主要富集在:蛋白质-FAD 连接、转运蛋白活性的负调控、离子跨膜转运蛋白活性的负调节、阴离子跨膜转运的负调控、阴离子通道活性的负调节、噻唑代谢过程等通路。

图6 共有差异蛋白的韦恩图分析Fig.6 Venn diagram analysis of common DEPs in A.venetum

图7 共同差异表达蛋白表达模式分析Fig.7 Expression patterns of the common DEPs
2.6 盐胁迫下罗布麻叶片差异蛋白网络互作分析
为了研究罗布麻在盐胁迫下如何通过蛋白互相作用进行盐胁迫信号的传递,笔者通过String分析,采用网络建模的方法,构建出蛋白质相互作用网络,从而挖掘出蛋白间相互作用中的关键蛋白(表2)。
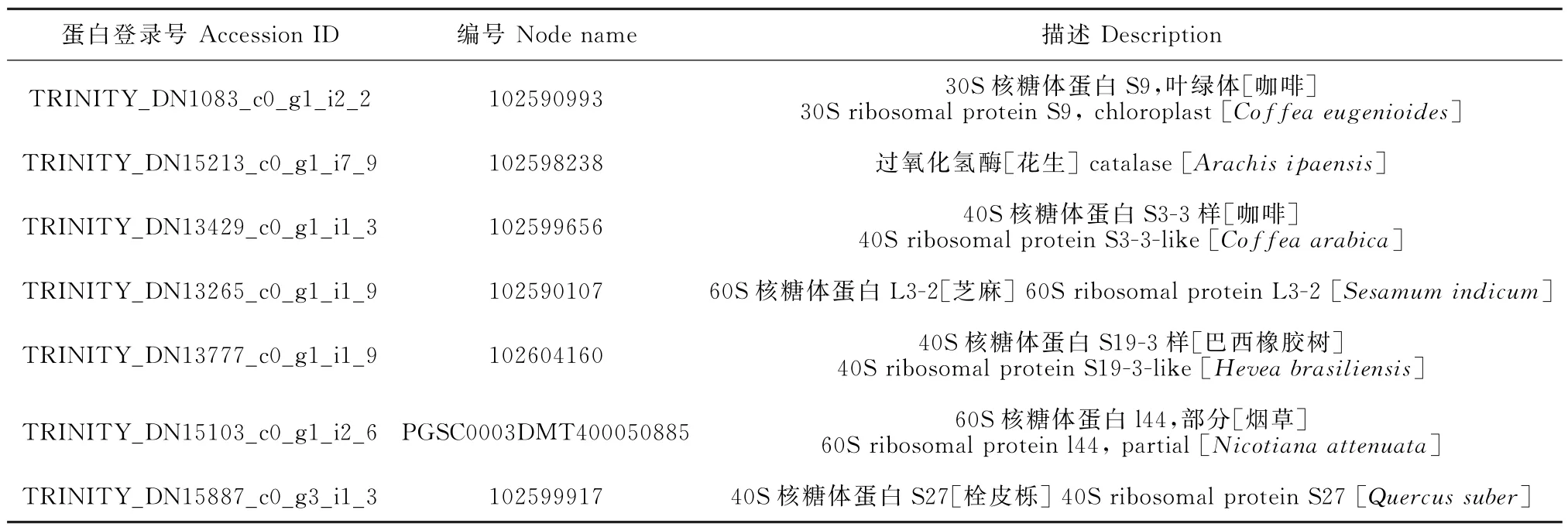
表2 蛋白网络分析中的名称描述Table 2 Description of protein names in the network analysis
对不同胁迫时间点的差异蛋白以及所有差异蛋白分别进行String分析,得到处于网络互作核心位置的蛋白(图8)。T2 vs T0组得到的差异表达蛋白中处于网络核心位置的高表达蛋白为30S核糖体蛋白S9(TRINITY_DN1083_c0_g1_i2_2)(图8,A);T6 vs T0组处于核心的高表达蛋白为40S核糖体蛋白S3-3样(TRINITY_DN13429_c0_g1_i1_3)(图8,B);T12 vs T0组的核心蛋白为未知蛋白(TRINITY_DN17640_c1_g1_i1_3)和60S核糖体蛋白L3-2(TRINITY_DN13265_c0_g1_i1_9)(图8,C);T24 vs T0 组为60S 核糖体蛋白L3-2(TRINITY_DN13265_c0_g1_i1_9)和40S核糖体蛋白S19-3样(TRINITY_DN23552_c0_g1_i1_3) (图8,D);T48 vs T0组为细胞分裂周期5样蛋白亚型X2(TRINITY_DN17302_c0_g2_i5_7)和60s核糖体蛋白l44(TRINITY_DN15103_c0_g1_i2_6)(图8,E)。对所有差异表达蛋白进行蛋白互作网络分析之后得到的核心蛋白为40S 核糖体蛋白S27(TRINITY_DN15887_c0_g3_i1_3)(图8,F)。

图8 差异蛋白的蛋白互作网络分析A.T2 vs T0;B.T6 vs T0;C.T12 vs T0;D.T24 vs T0;E.T48 vs T0;F.所有差异蛋白。Fig.8 Protein-protein interaction analysis of the DEPsA.T2 vs T0.B.T6 vs T0.C.T12 vs T0.D.T24 vs T0.E.T48 vs T0.F.All differential proteins.
2.7 差异蛋白表达水平PRM 验证
根据TMT 实验结果,随机选取8个与糖代谢、氧化还原、光合作用和渗透调节等过程相关的差异表达蛋白进行PRM(parallel reaction monitoring)验证分析。在T2 vs T0、T6 vs T0、T12 vs T0、T24 vs T0和T48 vs T0比较组中,8个蛋白的TMT 定量结果和PRM 定量结果中表达趋势一致。其中,TRINITY_ DN12352_c0_g1_i1_6、TRINITY_DN12386_c0_g1_i2_8和TRINITY_DN16647_c0_g1_i9_2,这3个蛋白在盐胁迫0 h、2 h、6 h、12 h、24 h和48 h均表现为上调表达且与TMT 结果趋势(表3)一致。PRM 结果表明蛋白组数据是可信的,能为进一步研究提供依据。

表3 差异表达蛋白的PRM 验证结果Table 3 PRM validation of the differentially expressed proteins
3 讨论
3.1 参与光合与能量代谢相关的蛋白质及作用
盐胁迫通过破坏生理过程,导致不充分的光合作用,使植物生长发育受到抑制。光合作用是一种光驱动的氧化还原过程,涉及将光能转化为化学能。在高等植物中,类囊体膜位于叶绿体中,嵌入类囊体膜的4种蛋白复合物参与电子传递过程包括:光系统Ⅱ(PSⅡ)、细胞色素(Cyt) b6/f、光系统 Ⅰ(PS Ⅰ)和ATP合酶复合物。2个光系统(PSⅠ和PSⅡ)都是色素-蛋白质复合物。Neelam 等研究表明PSⅡ在盐胁迫条件下会受到损伤,且与PSⅠ相比,PSⅡ的损伤率更高[13]。本研究中光系统Ⅰ相关蛋白在盐胁迫12 h、24 h和48 h上调,光系统Ⅱ相关蛋白在24 h下调(表1),表明光系统Ⅱ可能对盐胁迫更为敏感,利用光能驱动水的光解维持光合效率受到抑制。而光系统Ⅰ自12~48 h均保持稳定的表达丰度,表明48 h盐胁迫处理光系统Ⅰ未受到损伤。
由于盐胁迫作用导致植物能量代谢发生显著改变,本研究中蔗糖合酶(sucrose synthase,SuSy)在盐胁迫2 h、6 h、24 h、48 h时表现为上调表达,β-淀粉酶(beta-amylase 9,BMY)在12 h、24 h、48 h时表现为上调,ATP-柠檬酸合酶上调,乙醛脱氢酶(aldehyde dehydrogenase)6 h时下调表达,但在48 h时表现为上调。其余差异蛋白表现为下调,包括半胱氨酸合酶(cysteine synthase,OASA)在2 h时下调,ATP合酶12 h、24 h、48 h时下调,参与糖酵解的3-磷酸甘油醛脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GADP)、二磷酸醛缩酶(fructose-bisphosphate aldolase)以及参与柠檬酸循环的苹果酸脱氢酶(malate dehydrogenase,MAD)在2 h、6 h、12 h、24 h和48 h一直表现为下调表达(表1)。在NaCl胁迫下,植物降低能量代谢率以节约能量并限制ROS 进一步产生[14]。此前有报道称,在NaCl处理的拟南芥根中,糖酵解、柠檬酸循环、线粒体呼吸和磷酸戊糖途径组分的转录丰度普遍降低[15]。因此,在该研究中观察到参与糖酵解、柠檬酸循环的8种蛋白质的表达量在NaCl处理后的1个或多个时间点降低并不奇怪。
植物几丁质酶不仅作用于植物防御相关过程,而且在非生物胁迫耐受中也起重要作用。在拟南芥中,编码Ⅱ类几丁质酶的AtPR3基因被高盐诱导[16]。几丁质酶活性在短期胁迫时变化不明显,随着胁迫时间的增加而上升,本研究中有4个几丁质酶均在48 h上调表达(表1),这与盐胁迫下紫花苜蓿[17]的研究结果一致。
3.2 参与蛋白质合成和氨基酸代谢有关的蛋白质及作用
转录相关蛋白和转录因子在耐盐性中起关键作用,在盐胁迫条件下上调伸长因子有助于维持正常的蛋白质合成以提高耐盐性[18]。鉴定到的伸长因子(elongation factor)主要集中在6 h和12 h下调表达,在24 h 上调表达。真核翻译起始因子(eukaryotic translation initiation factor,eTIF)全部上调表达(表1)。前人研究发现真核翻译起始因子5A(eukaryotic translation initiation factor 5A,eIF5A)不仅能够直接调控蛋白质的合成,还能通过促进某些蛋白质的高效表达,间接调控细胞自噬、核质运输、mRNA 的降解及抵抗环境胁迫等[19]。这些伸长因子和真核翻译起始因子上调表达使罗布麻能够维持较高的耐盐性。
研究发现盐胁迫能够诱导许多与蛋白质合成相关的蛋白表达量发生变化[20]。本研究也鉴定到与蛋白质合成相关的差异表达蛋白,主要包括40S核糖体蛋白、60S 型核糖体蛋白、30S 核糖体蛋白和50S核糖体蛋白(表1)。在这项研究中蛋白质合成相关蛋白在盐胁迫下保持较高的数量和活性,与水稻在逆境下核糖体蛋白发生时空调控[21]相一致,表明核糖体蛋白基因家族是调控植物抗逆性的宝贵资源,在植物耐盐性方面发挥着重要作用。
3.3 参与抗氧化和次生代谢相关的蛋白质及作用
非生物胁迫导致植物体内活性氧(reactive oxygen species,ROS)的过度产生,ROS 一方面会对细胞成分造成损害,另一方面可以作为应激反应的信号分子[17]。植物可以通过复杂的机制调节ROS水平,例如用抗坏血酸过氧化物酶(APX)、谷胱甘肽过氧化物酶(GPX)、谷胱甘肽S-转移酶(GST)和超氧化物歧化酶(SOD)清除ROS。该研究中过氧化物酶(peroxidase,POD)和APX,这2种蛋白均表现出同样的趋势,即随着胁迫时间延长蛋白数量和表达量增加,该结果与前期生理指标测定结果中POD活性的变化趋势一致。罗布麻在盐胁迫下POD和APX 上调表达,与高盐胁迫下红树林(Kandelia candel)中的POD、SOD、APX 和CAT 蛋白质丰度显著增加[22]相一致。因此,这些参与氧化还原稳态的蛋白质呈现出上调表达意味着罗布麻启动了ROS清除机制,以减轻盐诱导的氧化胁迫,从而增强耐盐能力。
谷氨酸脱羧酶(glutamate decarboxylase,GAD)在拟南芥响应NaCl胁迫过程中,不同的GAD 蛋白表现出不同的表达特性[23]。本研究中仅鉴定到1个GAD,响应盐胁迫过程中在12 h、24 h、48 h 时下调,该结果与紫花苜蓿叶片响应盐胁迫的结果[20]一致。原因可能为GAD 不仅对NaCl有反应,而且还参与其他代谢过程,因为GAD4也被证明在拟南芥中被诱导以响应缺氧[24]、冷处理[25]和干旱胁迫[26]。本研究还鉴定到1 个查尔酮合酶(chalcone synthase,CHS)在6 h、12 h、24 h表达上调,与甘草在盐胁迫条件下高表达结果[27]一致。查尔酮是黄酮类形成的中间体,在植物对盐胁迫的反应中起重要作用[28]。
细胞色素P450(pytochrome P450,CYP)是植物中最大的蛋白质家族[29],广泛参与次生代谢产物的生物合成,并对植物响应非生物胁迫具有重要作用。研究表明小麦盐胁迫下TaCYP81D5 上调表达,加速消除ROS 增强小麦苗期和生殖期的耐盐性[30]。随着胁迫时间的延长,本研究中细胞色素P450蛋白的数量逐渐增加并且蛋白表达量也随之发生变化(表1),表明这些蛋白质在减轻植物损害中起重要作用,但随着胁迫加重其表达水平受到显著抑制。
3.4 信号转导和逆境防御相关蛋白质及作用
SOS信号途径与植物耐盐性密切相关,其实质就是调节细胞内Na+的动态平衡[31]。植物在NaCl胁迫下能够在几秒钟内提升Ca2+浓度,升高的Ca2+水平可激活处于SOS信号途径上游的Ca2+结合蛋白SOS3(salt overly sensitive 3),SOS3感知到Ca2+信号后与下游丝氨酸/苏氨酸蛋白激酶SOS2互作形成SOS3-SOS2 激酶复合体,在质膜H+-ATPase产生的质子梯度驱动下,将Na+从胞质移到表皮的质外体中[32]。植物细胞在响应盐胁迫维持渗透平衡的过程中,主要是通过调控根系导水率,水通道蛋白(AQP)能够调节植物根茎叶的渗透压,进而调控导水率动态变化[33]。对紫花苜蓿水通道蛋白基因MsPIP2;2的功能研究表明,在盐胁迫条件下,该基因能够通过促进转基因植物中Na+的外排和K+的保留来缓解盐毒害,高浓度(≥6 mmol/L)Ca2+可以增强MsPIP2;2转基因拟南芥的耐盐性[34]。本研究鉴定到的蛋白中参与SOS信号途径的蛋白包括:钙调蛋白(calmodulin,CAM)、钙依赖性蛋白激酶(calcium-dependent protein kinase,CDPK)、水通道蛋白、丝氨酸/苏氨酸蛋白激酶,其中CaM前期下调表达,到48 h时上调表达,丝氨酸/苏氨酸蛋白激酶在48 h下调表达。与离子稳态相关的水通道蛋白在上调表达,表明随着盐胁迫时间的延长,需要维持叶片内水分吸收和运动的需求增加。在这些蛋白的协同作用下完成罗布麻响应盐胁迫维持渗透平衡。
在非生物胁迫下,植物激素茉莉酸已被证明可有效提高植物的胁迫耐受性[35],丙二烯氧化环化酶(allene-oxide cyclase,AOC)是茉莉酸合成过程的关键酶[36],对番茄外源应用AOC 能够提高其耐盐能力。有研究发现AOC 在拟南芥响应盐胁迫过程中表达量下降[18],本研究在差异蛋白中检测到1个AOC在12 h、24 h、48 h时表达量下降,此结果与紫花苜蓿响应盐胁迫该蛋白表达量下降[37]一致。
在逆境条件下,热休克蛋白(heat shock 70 kD protein,HSP)可以通过重新折叠蛋白质来重建正常的蛋白质构象并维持细胞稳态,从而保护植物免受胁迫[38]。本研究中鉴定到的热休克蛋白在不同胁迫时间呈上调或下调表达,但蛋白数量随盐胁迫时间延长增加。金银花盐胁迫下3个热休克蛋白上调表达,以增强金银花对盐胁迫产生防御反应[7],但在烟叶中这种蛋白显著下调[39],这意味着这些蛋白质在不同时间、不同剂量胁迫条件下表现出不同的反应。
3.5 细胞分裂和细胞结构相关蛋白质及作用
本研究鉴定得到与细胞分裂、细胞结构维持相关的蛋白有2种,分别为肌动蛋白(actin)和微管蛋白(tubulin alpha)。肌动蛋白在真核生物的生命活动中占据重要地位,不仅是构建细胞骨架所必需的物质,而且在细胞运动、物质运输、能量和信息传递、细胞分裂等过程中起着重要作用[18]。罗布麻中的肌动蛋白在盐胁迫下表达量显著增加,但随着胁迫时间延长表达量降低,可能细胞结构受到一定程度的破坏。微管蛋白在植物细胞的分裂和生长中起着关键作用。NaCl处理暗发大豆结果表明,微管蛋白α链(I1K2I1)和微管蛋白β链(I1K7J4,I1LDR2)的丰度均因盐处理而降低,而微管蛋白特异性伴侣蛋白A(C6T5I8)的丰度增加[39]。本研究共发现8个微管蛋白,不同时间蛋白种类和表达量不同,在6 h时发现4个微管蛋白表达量全部增加,12 h时有1个微管蛋白降低,24 h时有3个蛋白表达量下降,到48 h时微管蛋白表达量全部降低,随着胁迫时间延长下调表达的微管蛋白数量逐渐增加,这些变化表明它们可能通过复杂的调节机制参与应对罗布麻的盐胁迫过程。
4 结论
用TMT 技术对盐胁迫下罗布麻叶片进行定量蛋白质组学研究,分析不同时间盐胁迫蛋白质组表达水平的差异,并对差异表达蛋白进行GO 和KEGG 分析以及String网络互作分析,根据盐胁迫下罗布麻蛋白质组表达水平的差异,筛选出罗布麻应答盐胁迫的关键蛋白。
(1)共鉴定得到12 457个蛋白,胁迫时间2 h、6 h、12 h、24 h 和48 h,得到差异蛋白分别为424,645,791,1 001,1 227个。对差异蛋白进行GO 富集分析表明,差异蛋白主要参与氧化还原酶活性途径、内肽酶调节活性、细胞壁、碳水化合物代谢过程等。KEGG 富集分析表明,差异蛋白参与最多的为核糖体、黄酮类生物合成、柠檬酸循环(TCA 循环)、黄酮类生物合成、淀粉和蔗糖代谢以及丙酮酸代谢等过程。
(2)对不同时间点差异蛋白进行韦恩分析,得到11个差异蛋白为共有蛋白,GO 富集分析发现,这些共有差异蛋白主要富集在蛋白质-FAD 连接、转运蛋白活性的负调控、离子跨膜转运蛋白活性的负调节、阴离子跨膜转运的负调控、阴离子通道活性的负调节和噻唑代谢过程等通路。
(3)差异蛋白网络互作分析发现,不同胁迫时间的差异蛋白网络互作所得到的蛋白主要以核糖体蛋白为主,其中60S 核糖体蛋白L3-2(TRINITY_DN13265_c0_g1_i1_9)在胁迫12 h和24 h的差异蛋白中均为核心蛋白,表明核糖体蛋白在罗布麻响应盐胁迫过程中发挥着重要作用。
