蜾蠃亚科物种的筑巢生物学研究现状
2024-02-26贺春玲刘潇宇
贺春玲, 王 翔, 刘潇宇
(河南科技大学园艺与植物保护学院, 洛阳 471000)
蜾蠃(potter wasps)是蜾蠃亚科(Eumeninae)物种的统称,是膜翅目(Hymenoptera)胡蜂科(Vespidae)中种类最丰富的一个亚科,全世界已知3族:蜾蠃族(Eumenini)、盾蜾蠃族(Odynerini)和长腹胡蜂族(Zethini)204 属 3 800 种(Carpenter, 1982; Hermesetal., 2014; Lietal., 2019; Brozoskietal., 2023);中国已知 51 属 267 种(Tanetal., 2018)。该亚科分布广泛,筑巢策略多样,生活习性从绝大多数独栖到极少数种类的原始社会性,对研究社会性昆虫演化具有重要价值(West-Eberhard, 2005; Hermesetal., 2013; Kelstrupetal., 2023)。该亚科绝大多数物种成虫吸食花蜜,幼虫取食亲代雌蜂贮备在巢室中的鳞翅目(Lepidoptera)和鞘翅目(Coleoptera)幼虫,偶见有取食膜翅目叶蜂的幼虫,它们对于农业生态系统和自然生态系统的植物授粉和害虫生物防治具有至关重要的作用(Fateryga and Amolin, 2014; Fateryga, 2018b; Dang and Nguyen, 2019; Lietal., 2019)。
筑巢生物学是研究筑巢蜂的筑巢策略、巢结构和功能以及巢内行为规律的科学,其目的是揭示筑巢蜂筑巢的多样性和科学性,为保护和应用筑巢蜂资源奠定基础。繁殖期筑巢是蜾蠃亚科物种生活史的重要阶段,是子代繁殖成功的重要保障(李铁生, 1993)。其筑巢由雌蜂完成,筑巢繁殖过程为:寻找合适的筑巢地点,建造巢室,在巢室内产1粒卵(部分有卵端丝粘于巢室壁上),捕猎其他昆虫的幼虫经蛰刺麻醉后带回巢室内贮存(贮存数量因种类而异),制作巢室隔板封闭巢室;然后继续建新的巢室并产卵, 直至繁殖期结束(Fateryga and Podunay, 2018; Dang and Nguyen, 2019);卵孵化后,幼虫吸食巢室内麻痹的猎物,经蜕皮、化蛹,最后羽化出成虫,成虫咬破巢室隔板,从巢内飞出,开始下一代的自由生活(李铁生, 1985; Kelstrupetal., 2018; Segolietal., 2020)。按照其筑巢场所、筑巢方式和基质的不同,传统上可将蜾蠃亚科物种的巢分为挖掘型(excavators)(在筑巢基质上挖掘洞穴建巢)、租住型(renters)(利用已有基质的空腔室建巢)和建造型(builders)(以筑巢基质为支撑通过采集筑巢材料建巢)3种类型(Cowan, 1991; Hugoetal., 2020)。然而,蜾蠃亚科物种的筑巢也具有很强的可塑性(de Castro Jacquesetal., 2022),同一物种有时也可以建造不同类型的巢(Hermesetal., 2013, 2015)。目前,国内外已报道蜾蠃亚科物种的筑巢生物学的研究文献少且零星分散。本文按照其传统的筑巢分类方法,从不同类型的巢结构入手,对国内外蜾蠃亚科的筑巢生物学文献进行较为系统的整理,同时,详细综述了蜾蠃亚科不同物种筑巢的策略和巢内行为,总结分析了影响蜾蠃筑巢的因素,希望本综述能引起更多的科研工作者关注这一重要类群,从筑巢生物学角度深入揭示其物种的价值和奥秘。
1 各类型巢的巢结构和筑巢行为
1.1 挖掘型
挖掘型筑巢的蜾蠃亚科多数是在土壤中筑巢,筑巢地点包括草地、土丘、盐碱地、沙滩等(Chiappa and Rojas, 1991; Fateryga, 2018b)。雌性蜾蠃选择平坦、斜坡或有裂缝的土壤上挖掘巢穴,并在内部建造1个或多个巢室。巢分为有巢塔和无巢塔2种类型,其中有巢塔的巢是在巢入口建造烟囱状的入口管道(Fateryga, 2013a, 2020b; Faterygaetal., 2017)(图1: A, B)。在挖掘巢和巢塔制作过程中,雌蜂采集水或叶片的汁液来浸泡筑巢土壤并制作成泥浆(Faterygaetal., 2017; Fateryga and Onchurov, 2021)。巢由内而外是由1个、1列或分叉状线性巢室排列而成,最外巢室距离巢口之间的空间称为前庭(Fateryga and Ivanov, 2013; Fateryga, 2013c)。

图1 挖掘型蜾蠃亚科的巢结构
筑巢时,首先选择巢址,雌蜂以触角接触地面,仔细搜索巢址附近有无障碍物,然后低空飞行选择最终筑巢地点;确定巢址后,雌蜂寻找水源,并取水后反刍至巢址基质表面,使水与土壤混合后制作小泥球,又将小泥球排列成一个圆圈,标志着未来的巢入口;然后沿着巢口的位置向内挖掘巢道建巢室,挖掘出的黏土颗粒有堆放在巢室入口制成巢塔的,也有不筑巢塔直接丢弃的,丢弃的位置一般会距巢口有一定的距离(Fateryga, 2013a; Faterygaetal., 2017; Fateryga and Onchurov, 2021)(图1: C, D, E);当雌蜂巢室建成后会在巢室内产1粒卵,卵常直接放置在巢室底部或有一白色丝状物将卵悬挂在巢室最内端的壁上;紧接着,雌蜂外出捕捉猎物带回巢室,贮备一定数量的猎物后,用泥建造隔板封闭巢室入口,有连续建造数个新的巢室,也有建好一个巢室封闭后,另外挖掘新巢道建巢室;最后,雌蜂用泥封堵巢入口(Fateryga and Ivanov, 2013; Fateryga and Amolin, 2014; Fateryga, 2020b)。在盐碱地筑巢的蜾蠃Brachyodynerusmagnificusmagnificus从巢附近藜科(Chenopodiaceae)两种盐生植物(盐地碱蓬Suaedasalsa和短苞叉毛蓬Petrosimoniaoppositifolia)的多汁叶子中吸取的汁液混合黏土筑巢(Faterygaetal., 2017)(图1: F, G, H)。
巢塔(turret)是雌蜂将建巢过程中挖掘出的黏土作为材料,或衔泥在筑巢位置建造的烟囱状的入口管道。巢塔开口有漏斗型和弯曲型(Smith, 1978; Fateryga and Ivanov, 2013)。巢塔伸展的方向为直立或从巢穴入口向重力方向偏转有一定夹角。雌蜂将巢建造完成后绝大多数种类会拆除巢塔,并用拆除的巢塔材料封赌巢口(Fateryga, 2013a, 2018b, 2020b; Fateryga and Amolin, 2014)。在建造巢塔的过程中,如果被雨水冲刷或者人为破坏,筑巢雌蜂会修补巢塔;如果巢塔筑好后,雌蜂在建造巢室的过程中,巢塔被损坏,雌蜂不会修补巢塔(Fateryga and Onchurov, 2021)。查阅的13篇蜾蠃亚科挖掘型筑巢的文献,包括12属16种,其中无巢塔筑巢的有2种,巢建成后完全拆除巢塔的有6种,巢塔未完全拆除的有1种,不拆除巢塔的有4种,未描述巢塔是否拆除的有3种(表1)。盾蜾蠃属Odynerus是已报道4种蜾蠃均为挖掘型筑巢。我国有关挖掘型蜾蠃的筑巢生物学尚未见报道。

表1 挖掘型蜾蠃亚科的巢结构
1.2 租住型
租住型是指蜾蠃亚科在中空植物茎秆、树干缝隙、蜗牛壳、有孔的木块等已存在空腔的场所内筑巢的物种(Fateryga and Podunay, 2018; Faterygaetal., 2020a; Maqbooletal., 2022)。雌性蜾蠃选择了空腔巢址后,首先清除空腔中的残留物,然后衔泥建造底塞,育幼巢室,在育幼巢室内产卵,捕食并麻醉猎物贮存在育幼巢室内,用泥土制作隔板封闭巢室(图2: D),每头雌蜂育幼巢室的数量依物种和巢址的环境而异,最后用泥土封堵巢口(Brozoskietal., 2023)。除用泥胶制作隔板和封堵巢口外,也有用碎石制作隔板和封堵巢口(Fateryga and Podunay, 2018)(图2: E)。蜾蠃Leptochiluslimbiferus在蜗牛Xeropictaderbentina的空壳中筑巢,1个蜗牛壳中仅有1个巢室(Faterygaetal., 2020a)(图2: B)。

图2 租住型蜾蠃亚科的巢结构
近年来,国内外学者利用蜾蠃在空腔室内筑巢的习性,通过野外安装陷阱巢监测筑巢蜾蠃亚科的种类、筑巢行为、生活史以及生境对其物种多样性的影响(黄敦元等, 2012; Staabetal., 2018; 窦飞越等, 2022; 林木青等, 2022)。在陷阱巢中筑巢的物种,巢结构包括底塞、育幼巢室、间室、巢室隔板、前庭和巢封口(Dang and Fateryga, 2021; Maqbooletal., 2022; Brozoskietal., 2023)(图2: A)。底塞是巢最内侧的结构,是雌蜂用筑巢材料涂抹接近巢管最内侧的位置,形成一个凹陷面光滑的隔板;巢最内端如果不平整,雌蜂会先建造底塞,用来限制幼虫只能在育幼室内活动,防止幼虫受伤和死亡;若巢管最内端光滑平整,雌蜂可能不会建造底塞。育幼巢室指雌蜂产卵和贮存幼虫食物的巢室;间室是指两个育幼室之间的空巢室,可能是防御寄生天敌从一个育幼室转移到邻近的育幼巢室的作用(Shimizuetal., 2012; Maqbooletal., 2022)。前庭是巢中最外侧的空室,较长的前庭能够减少子代在发育最初阶段的死亡率,同时防止其他雌性前来筑巢(Asísetal., 2007; Phametal., 2021)。底塞、间室和前庭在同一物种中并不是在每个巢内中都会出现,受巢的材料、内径、长度的影响,筑巢资源适合该物种筑巢时,巢空间利用率会更高(Maqbooletal., 2022; Tsujiietal., 2022)。
查阅的32篇蜾蠃亚科租赁型筑巢的文献,包括16属35种, 其中在空蜗牛壳中筑巢的有3种, 天牛幼虫蛀道中筑巢的有1种,蝗虫卵囊中筑巢的有1种, 野外中空茎秆和树缝中筑巢的有6种, 人工安装的陷阱巢监测到的筑巢种类有24种(表2)。佳盾蜾蠃属Euodynerus,短小蜾蠃属Leptochilus和Pachodynerus等物种多数是租住型筑巢。我国租住型筑巢蜾蠃的筑巢生物学研究文献涉及日本佳盾蜾蠃Euodynerusnipanicus(黄敦元等, 2013)和黄缘蜾蠃Anterhynchiumflavomarginatum(窦飞越等, 2022; 林木青等, 2022) 2种。
1.3 建造型
建造型是指蜾蠃亚科在建筑物、树叶、植物茎秆、岩石等基质上通过衔泥建造各种各样精美的泥陶巢的物种,最常见的是泥壶形(图3: A)(Méndez-Abarcaetal., 2012; Deshmukh, 2021)。建造型蜾蠃的每个巢的内部有1个巢室,筑巢时巢入口处存在漏斗形结构,建造完成后拆除作为封口材料(Lopes and Noll, 2019)。多数建造型的巢由几个巢室或数层巢室堆叠组成(图3: B, C),其巢室不呈线性排列(Méndez-Abarcaetal., 2012),不同属的巢大小形状差异很大,常见的泥壶形巢尺寸约为直径十几毫米的球形,有记录较大巢穴如Abispaaustraliana巢穴尺寸为15.6 cm×36.9 cm×34.5 cm(Matthews and Matthews, 2004)。黄斑蜾蠃属Katamenes是在石头的表面筑巢,巢室壁是由雌蜂采集的小碎石堆砌而成,然后用泥涂抹缝隙,巢的颈部和巢口是采集泥胶制成,当巢全部完成后,拆除漏斗形的巢口,巢口封堵后再用小碎石堆砌(Fateryga and Ivanov, 2022)。

图3 建造型蜾蠃亚科的巢结构
建造型巢和挖掘型、租住型巢在巢内的筑巢过程相似,不同的是建造型蜾蠃要用泥自己建巢室,建造的巢绝大多数都是暴露在基质的表面,因此相比于其他两种类型,建造型筑巢的物种的巢更容易被发现和观察。目前文献报道的建造型蜾蠃自然筑巢的种类较多,巢建筑和巢结构也多种多样(图3)。巢穴的主要建造材料为黏土和砾石,还有一些物种使用其他材料对已经建成的巢室进行伪装,避免遭受天敌攻击(Lopes and Noll, 2019; Fateryga and Ivanov, 2022);如:蜾蠃Pirhosigmasuperficiale和Pirhosigmalimpidum的巢室外表壁上均用一些比巢更长的植物材料进行伪装(Hermesetal., 2013);蜾蠃Minixisuffusum的雌蜂收集鸟类排泄物覆盖巢穴外表面(Aukoetal., 2015),这些伪装能阻止在筑巢期间以产卵器刺破巢室壁或从巢穴入口进入的天敌,但是无法阻止寄生在幼虫食物中的天敌。
Abispa属蜾蠃有着非常巨大的巢,有记录的最大巢穴长40 mm,开口处直径16 mm,重达0.5 kg(Matthews and Matthews, 2009),巢入口存在巨大的漏斗状结构(图3: D)。对Abispaephippium巢进行了拆除漏斗和破坏巢室壁试验,每次拆除漏斗后雌蜂都会立刻开始重建,破坏的巢室壁也会重新涂抹泥浆,但是当漏斗状结构破裂时,雌性的反应不激烈也不立即开始修复漏斗,说明漏斗状结构在筑巢时的优先级并不高,并且巢室建造完成雌蜂就会拆除漏斗作为封口材料(Matthews and Matthews, 2004, 2009)。Abispa属的巨大漏斗结构功能类似巢塔,雌蜂会守在漏斗结构中攻击靠近的天敌,而其他建造型巢穴如大华丽蜾嬴Deltapyriforme和Eumenesfraternus等物种的漏斗比较小。建造漏斗的作用是:(1)方便雌蜂把腹部伸入巢室产卵;(2)保持巢室内的环境相对稳定;(3)为了便于封口,雌蜂在封口时先衔泥把巢入口封住,再用水润湿漏斗,以拆除漏斗的材料制作封口(Slmok, 2008; Matthews and Matthews, 2009; 经福林等, 2010; Grissell, 2014)。
已查阅的建造型蜾蠃筑巢特征见表3,共记录16属27种。筑巢支撑物的选择有建筑物、树叶、植物茎秆、岩石等,其中在人工建筑物上筑巢的最多。国内仅记录3种建造型巢穴,分别是孪蜾蠃Eumenesfraternus、大华丽蜾蠃Deltapyriforme和原野华丽蜾蠃Deltacampaniformeesuriens。
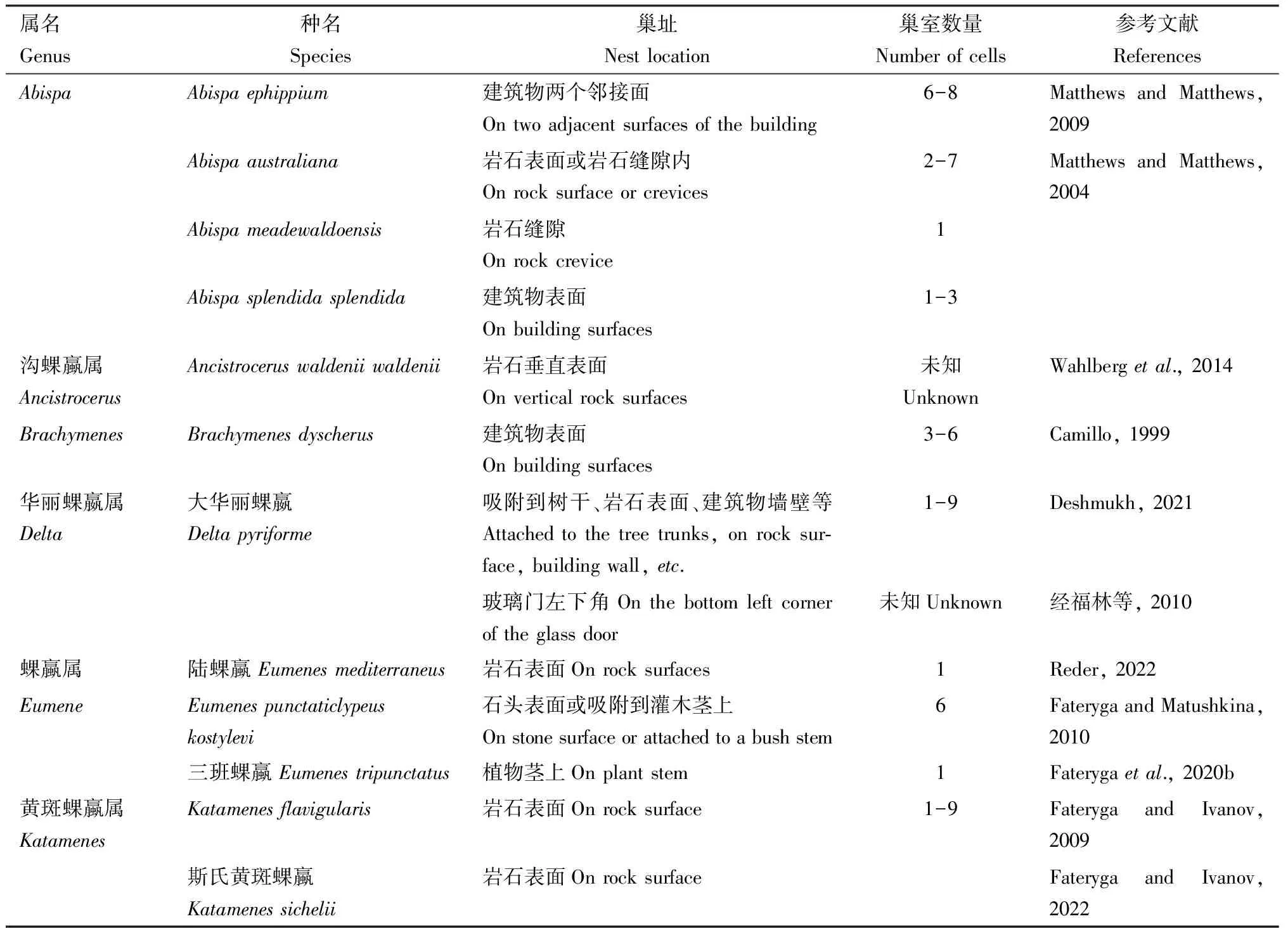
表3 建造型蜾蠃亚科的巢特征
1.4 筑巢可塑性
蜾蠃亚科的巢结构纷繁、多样,除上述3大类型外,因蜾蠃的原始社会性行为、与其他物种同巢共存、建造假巢、物种间篡夺巢室和猎物等行为,使得蜾蠃的巢结构更加多样和复杂。
蜾蠃亚科物种绝大多数是独栖性,少数表现为原始社会性行为,这些原始社会性蜾蠃的筑巢过程和独栖性蜾蠃的筑巢过程有所不同。如蜾蠃Montezumiacortesioides的巢由数个巢室堆叠而成,蜾蠃M.brethesi的巢结构为多只雌蜂在同一区域建造数十个泥球型的巢室,2种蜾蠃雌蜂均采用渐进式为子代供应猎物,有巢室守卫行为,同时也发现猎物盗窃、巢室竞争和共用巢室等现象(West-Eberhard, 2005; Lopes and Noll, 2019)。蜾蠃Xenorhynchiumnitidulum的1个巢中存在不止1头雌性,没有分工,同种其他雌性也在巢中重复使用巢穴(Srinivasan and Girish Kumar, 2009)。蜾蠃Abispaephippium雌蜂通过渐进式供应、清除碎片、修复巢穴损伤和攻击天敌等行为积极守卫巢穴,在子代幼虫化蛹前拆除巢口漏斗状结构封堵巢口,接着,再在巢室附近建新的巢室(Matthews and Matthews, 2009)。
蜾蠃Antepiponabiguttata利用泥蜂Sceliphronmadraspatanum的旧巢或占用新巢筑巢,并在巢的外面建巢塔,巢塔的端部用泥封堵(Phametal., 2021)。蜾蠃Montezumiatermitophila在白蚁Constrictotermescyphergaster巢穴中筑巢,二者在巢中共存,无利无害(Hugoetal., 2020)。蜾蠃Pseudabispaparagioides雌蜂攻击杀死蜾蠃Abispaephippium雌蜂并抢占后者的巢,但是前者不会破坏已经筑好的巢室,后者的卵能够正常发育并羽化(Matthews and Matthews, 2010)。更有意思的是一些种类会用植物材料甚至鸟类来对巢室进行伪装,如:同属的Pirhosigmasuperficiale和P.limpidum2种蜾蠃会在建造的巢室表面进一步用植物材料装饰巢壁(Hermesetal., 2013);蜾蠃Minixisuffusum会采集鸟粪对巢壁进行伪装(Aukoetal., 2015);蜾蠃Tricarinodynerusguerinii在空腔中使用泥浆筑巢,并建造了一个没有任何空隙的巢塔,在密封巢穴时不会拆除(Gess and Gess, 1991)。蜾蠃Minixibrasilianum,Eumenespapillarius和Eumenessp.在芦苇管中建造泥壶型的巢室,其巢既是建造型也是租住型(Hermesetal., 2015; Ivanovetal., 2019)(图4)。

图4 蜾蠃亚科特殊的巢结构
2 影响蜾蠃亚科筑巢的因素
2.1 筑巢的材料资源
蜾蠃亚科筑巢的主要材料是土壤和水源,它们不论采用何种方式筑巢,均为1室1卵,在巢室之间建造隔板来分割巢室,通常使用混合(或不混合)植物组织的土壤胶泥来建造巢室隔板并修饰它们的巢(Boesietal., 2005; Pannureetal., 2016)。然而,可用筑巢材料受筑巢地点周边的环境影响,也是长期对分布环境的适应性演化。在非洲西南部沿海沙丘缺乏土壤的地区,在蜗牛壳中筑巢的蜾蠃Alastorricae,巢室隔板是沙丘地区常见的沙砾(Gess and Gess, 2008);在干旱缺水的地区,雌蜂可以选择血淋巴、肉质叶片汁液或花蜜等替代水(Cowan, 1991; Faterygaetal., 2017);雌蜂也会用砾石代替黏土,以花蜜混合土壤制作少量黏土来建造隔板和封口(Hermesetal., 2013; Aukoetal., 2015; Fateryga and Podunay, 2018)。因此,从大尺度生境来看,筑巢的材料资源限制了蜾蠃巢穴的分布和数量,但可能不影响蜾蠃的筑巢行为。
筑巢蜂能够在相对较远的距离觅食(Kleinetal., 2004),这可能使它们能够有效地寻找其他筑巢地点。然而,由于人为活动对自然森林环境的影响和破碎化,筑巢资源变得更加稀缺 (Ferreiraetal., 2015; Toledo-Hernándezetal., 2022),可能导致一些个体在不太有利的条件下筑巢。蜾蠃通常会在环境中利用大量的筑巢基质,包括人造基质,这可能显示出高度的可塑性和自然适应性(Méndez-Abarcaetal., 2012)。在不同地区,蜾蠃Pachodynerusnasidens选择不同的基质筑巢,有人工安装的巢管(Houseetal., 2020)、废弃的建造型蜾蠃Zetaargillaceum的巢(Matthews and González, 2004)以及社会性胡蜂科马蜂亚科(Polistinae)物种的巢内(de Castro Jacquesetal., 2022)等。已有报道不同的巢管直径会产生不同的性别比例,在较小直径的巢中产卵的雄性相对较多(Polidorietal., 2011);巢管的内径和长度显著地影响育幼室数量,而育幼室和非育幼室数量显著地负影响被寄生率(林木青等, 2022)。因此,清楚蜾蠃亚科不同物种的筑巢生物学特点对利用和保护其资源具有非常重要的意义。
2.2 筑巢的食物资源
食物供应是亲代照顾的最基本形式之一。在后代依赖供应的动物中,所提供食物的数量和质量可能对它们的发育、生存、体型和繁殖成功产生深远的影响。食物数量和质量的重要性在大量供应的昆虫(主要是蜜蜂和胡蜂)中可能更加至关重要,在这些昆虫中,每个后代在密封的室中获得固定数量的食物(Fieldetal., 2020),在这种情况下,营养不足不能通过后代在发育后期获得更多的食物来弥补。在捕食性胡蜂中,猎物的数量、大小、种类和营养价值都被证明对后代的发育有重要影响(Field, 1992; Strohm, 2000; Judd and Fasnacht, 2017)。当猎物充足时,雌性蜾蠃在巢穴中建造更多的育幼室并提供较多的猎物,后代性别一般按照雌性在内、雄性在外的方式排列,雌蜂会在雌性育幼室中放置比雄性更多的猎物(Cowan, 1983; 郭鹏飞等, 2018; 林木青等, 2022)。蜾蠃通常为雌性后代提供比雄性后代更多的食物,提供的猎物数量主要是由胡蜂对产卵性别的记忆决定的(Cowan, 1983, 1991; Tsujiietal., 2022)。
捕获猎物为子代贮备食物是雌蜂筑巢花费精力最大的投资(李铁生, 1985; Kelstrupetal., 2018; Segolietal., 2020)。雌性蜾蠃将猎物蛰刺后带回巢室内作为幼虫的食物,存在2种猎物放置方式:一种是渐进式供应(progressive provisioning),雌蜂持续捕捉猎物直到幼虫停止进食(West-Eberhard, 2005; Kelstrupetal., 2018);另一种是截断式供应(truncated progressive provisioning),雌性快速捕捉放置一定量的猎物,然后停止供应并封闭巢室(Matthews and Matthews, 2009; Fieldetal., 2020)。大多数蜾蠃都是截断式供应,少数原始社会性蜾蠃是渐进式供应,渐进式供应被视为从独栖性向社会性演化的重要特征。截断式供应的蜾蠃雌蜂在育幼室放置足够的食物后,便开始封口建造新的巢穴或育幼室,其后代数量较多。渐进式供应的蜾蠃Montezumiacortesioides在幼虫期持续为子代提供食物,其子代数量较少(Field, 2005; West-Eberhard, 2005)。相比于截断式供应,渐进式供应的雌蜂会守在巢穴附近,降低了被天敌攻击风险,但是当雌蜂捕猎过程中死亡,这只雌蜂喂食的后代可能因没有足够的食物而导致死亡。
蜾蠃在捕猎食物的过程中,如果捕猎已被天敌寄生的猎物将会对其子代产生影响。通常与被寄生猎物幼虫一起发育的蜾蠃幼虫发育成功率较低,蜾蠃幼虫对巢室内已被寄生的幼虫取食效率低于未寄生幼虫。然而,被寄生的幼虫通常更重,对捕食者的攻击反应更弱,这可能使它们更有吸引力,也更容易被捕食。然而,捕猎被寄生的幼虫食物会影响其子代的发育成功,表现为子代存活率和蛹质量与巢室中被寄生干尸幼虫的存在和数量呈负相关(Segolietal., 2020)。蜾蠃捕猎已寄生的猎物不仅对子代会有影响,同时对猎物的寄生天敌种群也会产生影响,这是非常有趣的科学问题,也有待更多的科学工作者对其进行深入研究。
2.3 天敌对蜾蠃筑巢的影响
蜾蠃亚科物种丰富,寄生蜾蠃的天敌种类和寄生方式也复杂多样。寄生后导致蜾蠃寄主死亡的天敌种类主要有双翅目(Diptera)的蜂虻科(Bombyliidae)、蚤蝇科(Phoridae)、麻蝇科(Sarcophagidae)和寄蝇科(Tachinidae),膜翅目青蜂科(Chrysididae)、钩腹姬蜂科(Trigonalidae)、蚁科(Formicidae)、姬小蜂科(Eulophidae) 和姬蜂科(Ichneumonidae),捻翅目(Strepsiptera)的胡蜂科(Xenidae)等(Fateryga and Amolin, 2014; Martynova and Fateryga, 2015; Dang and Nguyen, 2019; Dang and Fateryga, 2021)。已报道黄唇喙蜾蠃Rhynchiumbrunneumbrunneum的717个育幼巢室,死亡率占46.7%,其中被6种天敌寄生死亡占死亡总数量的51.3%(Dang and Nguyen, 2019);四秀蜾蠃Pareumenesquadrispinosus的793个子代巢室内,发育阶段死亡率占54%,其中被9种天敌寄生死亡占死亡总数量的48%(Dang and Fateryga, 2021)。
不同的天敌对寄主的寄生行为以及对寄主的影响不同。如:青蜂科和蜂虻科属于盗寄生类群,它们将卵产在蜾蠃育幼巢室,其幼虫会杀死蜾蠃的幼虫,然后以育幼巢室内的猎物为食,从而影响蜾蠃子代的正常发育(Matthews and Matthews, 2009, 2010; Shuttleworth, 2019)。胡蜾蠃属的Jucancistroceruscaspicus被青蜂科Chrysisrutilans寄生的死亡率占子代死亡率的11.5% (Fateryga and Amolin, 2014)。姬小蜂科Eulophidae sp.外寄生于四秀蜾蠃的蛹上,孵化后在蜾蠃的体表吸食体液,最终导致寄主死亡(Dang and Fateryga, 2021)。钩腹蜂科(Trigonalyidae)物种以植食性的鳞翅幼虫或叶蜂幼虫作为第1寄主并在其体内孵化,以蜾蠃等捕食性物种的幼虫作为第2寄主,当蜾蠃幼虫取食鳞翅目幼虫时进入第2寄主体内,直到寄主进入预蛹期时,钩腹蜂幼虫才会从寄主的体节间钻出,在数天内吸食掉寄主的体液,然后化蛹,最终羽化成成虫(Murphyetal., 2009; Väänänenetal., 2018)。捻翅目幼虫内寄生于寄主体内,不会造成寄主死亡,但会对寄主的生理和行为造成影响(Strakaetal., 2011; Beanietal., 2021),有关捻翅目寄生对蜾蠃物种的种群的影响还有待进一步研究。
此外,螨类共生物种会依附于独栖蜂的体表,甚至部分独栖蜂在虫体的不同部位进化出专门容纳螨的区域(Pereiraetal., 2018)。这些螨类在独栖蜂巢中以真菌和微生物为食。共生螨Ensliniellaparasitica与蜾蠃Allodynerusdelphinalis之间存在一种互惠关系,蜾蠃为螨类提供营养条件,螨类保护蜾蠃幼虫免受寄生蜂的侵害(Okabe and Makino, 2008)。蚂蚁与蜾蠃更像一种资源竞争的关系,并非致死性天敌,但是蚂蚁会搬走蜾蠃雌蜂捕捉到的猎物和已产下的卵,直接影响蜾蠃筑巢过程;如:Cataglyphisaenescens工蚁会搬走雌蜂倾倒至地面的水(Fateryga, 2013c),蚂蚁Pheidolesp. 会搬走麻痹的鳞翅目幼虫(Matthews and Matthews, 2009)。
3 小结与展望
蜾蠃亚科以多种多样的筑巢策略适应其生存的环境,传统上将蜾蠃亚科物种的巢分为挖掘型、租住型和建造型3种类型。虽然这样的分类方法并非按照蜾蠃亚科的物种分类系统进行分类,但我们在文献检索的44属77种蜾蠃的筑巢生物学文献中发现,同属的物种很少发现有不同类型的筑巢策略,仅发现在华丽蜾蠃属Delta的2种蜾蠃存在2类筑巢类型,其中Deltadimidiatepeenne是租赁型,而大华丽蜾嬴Deltapyriforme是建造型筑巢。已有报道黄斑蜾蠃属Katamenes物种均在裸露的在石头表面采用建造型筑巢(Fateryga and Ivanov, 2019, 2022);Pirhosigma的蜾蠃均会用植物材料伪装巢室(Hermesetal., 2013);佳盾蜾蠃属、喙蜾蠃属和Pachodynerus已报道的14种蜾蠃均是租赁型筑巢策略。相比于蜾蠃亚科3 800种的物种数量,目前的研究还不能很好地将蜾蠃亚科的筑巢生物学特征和分类学地位结合起来,但这些文献初步说明蜾蠃亚科物种筑巢可能在属级水平上具有一定的稳定性,随着持续不断地对更多的蜾蠃筑巢生物学的研究和发现,未来会有更多的证据理解蜾蠃亚科的筑巣策略和系统演化规律。
蜾蠃在筑巢过程中,需要搬运的主要材料是泥球和猎物。观察蜾蠃衔泥和捕猎行为发现雌蜂能够根据自己的体型制造合适大小的泥球,而猎物的大小会随季节变化而不同,当鳞翅目幼虫大小超过雌性蜾蠃的负载或雌性蜾蠃只能捕捉到较大的鳞翅目幼虫时,雌性蜾蠃把猎物带回巢穴的飞行过程中可能会摔倒然后丢弃猎物(Edgar and Coelho, 2000)。体型大小作为种群的特征,对种群动态变化和生态群落内的竞争关系有很强的影响(Britoetal., 2020)。在同一生境,不同物种对猎物也有一定的偏好,同时,猎物种类和筑巢地点的偏好会因当地情况而有所不同(Wahlbergetal., 2014),有关不同物种对猎物的选择与物种个体特征之间的关系有待进一步研究。另外,环境条件对蜾蠃亚科的筑巢生物学也会产生影响,陷阱巢的筑巢率与特定地区相应季节开始时的降雨量之间有很强的相关性(Ivanovetal., 2019)。在性别比例上表现出相当大的季节性变化,夏季繁殖代的雌性多于雄性,越冬代的雄性多于雌性;越冬代对雄性的相对投资增加,可能是为了补偿冬季雄性死亡率增加,也可能是越冬代个体后代遗传变异增加的一种手段(Longair, 1981; Boesietal., 2005)。因此,蜾蠃筑巢中的食物供应与环境条件之间的关系以及对子代的影响还需要进行深入研究。
蜾蠃亚科物种是重要的捕食性天敌昆虫。蜾蠃亚科物种多样性高、营养级复杂、捕猎种类多、捕食量大,在生态系统中发挥着不可替代的作用,但目前对蜾蠃亚科筑巢生物学的系统研究还非常有限,尤其是蜾蠃捕猎的食物种类和寄生蜾蠃的天敌种类都很少识别到种的水平(Vargasetal., 2014),这对于我们认识、保护和应用物种的功能是不利的。我国蜾蠃亚科物种丰富,分布范围广,但目前有关对蜾蠃亚科的筑巢生物学知之甚少。关于未来的研究建议如下:(1)研究蜾蠃亚科物种在不同生境中的筑巢生物学特点;(2)研究植物-植食性害虫-蜾蠃亚科物种-天敌昆虫之间的食物网关系,进而揭示不同蜾蠃亚科物种在农业和自然生态系统的功能;(3)优势蜾蠃亚科物种在害虫生态防控中的应用研究。
致谢感谢重庆师范大学李廷景教授为本文蜾蠃亚科部分物种提供中文学名。
