城市森林群落林下草本植物多样性及其环境响应
——以贵阳市为例
2024-01-27蔡洪丽
蔡洪丽,欧 静
(贵州大学林学院,贵州 贵阳 550025)
林下草本植物是森林群落的重要组成部分,占森林中所有维管植物种类的80%以上[1—2],在促进土壤养分循环、能量流动,群落演替和维持森林稳定性等方面都发挥着积极作用。在森林群落中,林下草本植物能与乔木幼苗竞争资源,影响树苗的存活、发育和生长[3—4],可作为样地质量、乔木层再生模式及稳定状态的一个重要指标[5—6],其物种分布及多样性格局对森林群落整体发育至关重要。乔木层占据森林群落生物量的主要部分,导致森林生态学的基本理论几乎来自对乔木层的研究,相较于乔木层,草本层的多样性得到的关注度还远远不够[7]。近年来,城市化发展使城市森林遭受严重的人为干扰[8],因此,探究城市林下草本植物物种组成及多样性的分布格局,对进一步阐明城市森林物种多样性维持和生态环境保护具有重要意义。
物种多样性反映了植物群落的稳定性和可持续性水平,体现了植物群落结构类型、物种组成和生境条件的差异[9]。明确林下草本植物物种多样性的主要影响因素已然成为森林生态学近年来的一个热点[10—12]。多项研究表明,森林群落中的草本植物分布及多样性格局受光照、温度、土壤、地形等多种环境因素影响[13—15]。秦浩等[16]研究表明,海拔是影响林下草本植物多样性的主要因素。同时,草本植物生长少不了土壤这一必要介质,土壤作为养分供给场所,更是影响草本植物多样性的直接因素[17]。但对于林下草本植物来说,上层乔木也会影响其多样性。有研究认为林下草本植物多样性与林分空间结构密切相关,林分类型、林分密度、郁闭度、林龄、发育程度等因素能够直接或间接影响其多样性[18]。左永令等[19]的研究也说明,植被类型对草本植物物种组成与分布具有较好的解释力。
贵阳市是我国西南地区典型喀斯特城市,森林植物多样性保护、植被恢复难度极大[20]。要维持森林群落物种多样性与稳定性,尤其需要重视林下草本植物的发展。在喀斯特地区,人们比较关注草地生态系统的植物物种组成与土壤结构[21]、木本植物群落结构与多样性变化[22],以及植物多样性与地形的关系[23],但缺乏针对林下草本植物物种多样性与环境因子关系的报道。该研究基于贵阳市城市森林典型群落样地林下草本植物调查和多样性研究,分析不同群落林下草本植物组成特征及不同林分类型草本植物的多样性分布格局,探究物种多样性与环境因子的关系,期望对黔中地区城市森林群落物种多样性保护与优化提供理论依据。
1 材料与方法
1.1 研究区概况
贵阳市地理位置处于 26°11′~26°55′N、106°07′~107°17′E,位于云贵高原东部,黔中山原丘陵中部(图1)。面积约8043 km2,平均海拔1100 m。具有典型喀斯特岩溶地貌,属亚热带季风性湿润气候,年平均气温15.3 ℃,年均降雨量1129.5 mm,平均相对湿度80%左右,年平均阴天日数为235.1 d。截至2022 年,森林覆盖率55.3%,林下草本植物丰富。城市森林主要树种为马尾松Pinusmassoniana、杉木Cunninghamialanceolata、柏木Cupressus funebris、华山松Pinusarmandii、樟Camphora officinarum、朴树Celtissinensis、女贞Ligustrum lucidum、枫香Liquidambarformosana等。

图1 研究区位与样地分布Fig. 1 Study area and sample plot distribution
1.2 样地设置与植物调查
调研时间为2022 年5~9 月,依据城市森林分布的代表性、均匀性,选择人为干扰较少、植被生长状况较好的地段,保障能够基本涵盖各种地形条件和主要的植被类型,设置森林群落样地开展林下草本植物调查。为更加全面地掌握不同森林群落林下草本植物分布特征,选择23 种森林群落,共44个样地(图1),每个样地面积1 hm2,并将23 种群落划分成5 种林分类型,分别为针叶纯林(Ⅰ )、针叶混交林( Ⅱ)、阔叶纯林(Ⅲ )、阔叶混交林( Ⅳ)、针阔叶混交林( Ⅴ)( 表1),以探究不同林分类型林下草本植物的多样性情况[24]。参考方精云等[25]的群落清查方法,在样地内均匀布设3 条100 m 长的平行样线,在每条样线上每隔10 m 布设一个1 m × 1 m 草本植物样方,共1320 个。记录每个1 m × 1 m 样方内的草本植物的种名、株数、高度、盖度、生活力,草本植物的科、属及物种学名参考《中国植物志》。

表1 样地基本信息Table 1 Basic information of sample plots
1.3 环境因子测定
调查并记录每个样地的林分类型、乔木密度、郁闭度、凋落物厚度、海拔、坡度、坡向、坡位(表1)。将针叶纯林、针叶混交林、阔叶纯林、阔叶混交林、针阔叶混交林5 种林分类型分布赋值为1、2、3、4、5。乔木密度通过计算单位面积内乔木株数得出,郁闭度采用树冠投影法测量。凋落物厚度测量样地内每个1 m2样方的枯枝落叶层厚度,取平均值。坡向利用公式NI=COS[RADIANS(Aspect)]得到标准化坡向值,其中Aspect 为坡向,指数的范围为[-1,1],数值越大说明坡向越倾向于北。坡位划分为山脊、上坡、中坡、下坡,分别赋值为1、2、3、4。
1.4 林下草本植物重要值及物种多样性指数计算
参照原野等[26]、马克平等[27]、陈廷贵等[28]、Hurlbert[29]的物种多样性测度方法,采用重要值(IV)衡量林下草本植物在森林群落中的相对重要程度。采用Margalef 丰富度指数(R)、Shannon-Wiener 多样性指数(H)、Simpson 优势度指数(D)、Pielou 均匀度指数(J)评价林下草本植物物种多样性。计算公式如下:
重要值(IV):
式中,rA 表示相对多度,rF 表示相对频度,rC 表示相对盖度。
Margalef 指数(R):
Shannon-Wiener 指数(H):
Simpson 指数(D):
Pielou 指数(J):
式中,S表示样地内物种数,N表示样地内物种个体总数,Pi表示种i的个体数占样地内总个体数的比例,Ni表示种i的个体数。
1.5 数据处理
运用Canoco5 软件进行物种分布与环境因子的关系分析,预先的除趋势对应分析(DCA)显示最长梯度长度为6.6,所以物种分布与环境因子的关系采用典范对应分析(CCA)[30],对44 个样地、286 个物种建立物种-重要值(44×286)矩阵,并对出现频率过低的物种进行降权处理,仅分析重要值排名前50 的物种。采用单因素方差分析(One-Way ANOVA)探究不同林分类型草本植物多样性的差异性、相关性分析探究物种多样性与环境因子的关系、多元线性逐步回归分析探究影响林下草本植物多样性的主导环境因子,回归分析中引入哑变量将分类变量林分类型和坡位两个环境因子进行量化,均运用SPSS 软件操作。利用Origin Pro 2021 软件绘图。
2 结果与分析
2.1 林下草本植物组成特征
2.1.1 物种组成特征
研究区内共调查到林下草本维管植物257 种(附录),分属67 科186 属;种子植物226 种,占总种数的87.94%;蕨类31 种,占总种数的12.06%;多年生草本177 种,占总种数的68.87%;一二年生草本80 种,占总种数的31.12%。
2.1.2 优势科、属
优势科为菊科、禾本科、百合科、唇形科4 个科(图2)。菊科含有27 属38 种,包括紫菀Aster tataricus、千里光Senecioscandens、宽叶兔儿风Ainsliaealatifolia、假福王草Paraprenanthessororia等物种。禾本科有15 属19 种,主要有淡竹叶Lophatherumgracile、芒Miscanthussinensis、求米草Oplismenusundulatifolius、显子草Phaenosperma globosa等。百合科有14 属18 种,主要是大百合Cardiocrinumgiganteum、多花黄精Polygonatum cyrtonema、山麦冬Liriopespicata、万寿竹Disporum cantoniense等物种。唇形科8 属14 种,主要物种为华鼠尾草Salviachinensis、灯笼草Clinopodium polycephalum、细风轮菜Clinopodiumgracile等。
物种数包含10 种及以上的多种科有4 个,占总科数5.97%;包含64 属,占总属数34.41%;种数为89 种,占总种数34.63%。种数含5~9 种的中等科有堇菜科、伞形科、凤尾蕨科、兰科、毛茛科等10科,占总科数14.93%;属数为47,占总属数的25.27%;种数为74,占总种数的28.79%。含2~4种的少种科有25 个,占总科数37.31%;包含47 属,占总属数的25.27%;种数为66,占总种数的25.68%。单种科28 个,占总科数41.79%;含28 属,占总属数的15.05%;含28 种,占总种数10.89%(表2)。
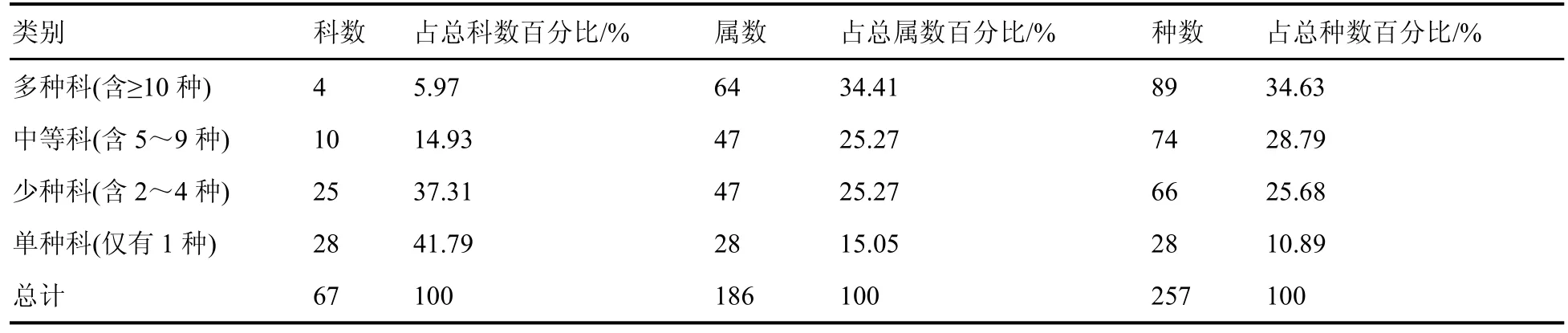
表2 林下草本植物科的类别统计Table 2 Class statistics of understory herbaceous plant families
样地内林下草本植物共包含186 属,以单种属(137 属)占主要优势,包含的种数占所有物种数的53.31%,说明属的组成具有分散性。含9 种以上的属有1 个,为堇菜属;含5~9 种的属仅1 个,为凤尾蕨属;含2~4 种的少种属有47 属,占总属数的25.27%,包含105 种,占总种数的40.86%(表3)。
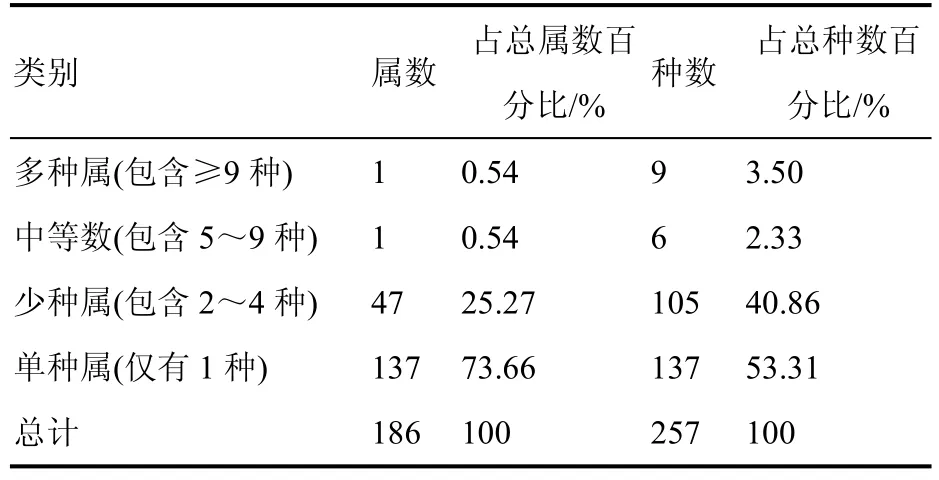
表3 林下草本植物属的类别统计Table 3 Class statistics of understory herbaceous plant genera
2.2 物种重要值分析
2.2.1 研究区林下草本植物重要值分析
在所有样地内,重要值最高的物种是求米草,为12.96%,其相对优势度在研究区内远高于其他草本植物,其次毛蕨Cyclosorusinterruptus、芒、蕨Pteridiumaquilinum、狗脊Woodwardiajaponica、对马耳蕨Polystichumtsussimense、芒萁Dicranopteris dichotoma、竹叶草Oplismenuscompositus、淡竹叶、地果Ficustikoua、中华薹草Carexchinensis的重要值在2%以上(表4)。这些物种在研究区的森林群落中占有相对较高的优势度,在群落物种组成和发展中起主要作用。

表4 林下草本植物优势种及其重要值Table 4 Dominant species of undergrowth herbs and their important values
2.2.2 不同群落林下草本植物重要值分析
不同群落类型林下草本植物的优势种各不相同(图3)。在23 种群落类型中,刺槐纯林、枫香纯林、华山松桦木混交林、华山松纯林、亮叶桦纯林、马尾松纯林、马尾松青岩油杉混交林、青岩油杉纯林、杉木混交林、杉木纯林、香樟女贞混交林、马尾松刺槐混交林12 种群落林下草本植物的优势种的优势度明显,优势种与次要优势种相对重要值差值≥5%,物种分布不均匀;华山松朴树混交林、马尾松杉木混交林、朴树女贞混交林、香樟石楠混交林4种群落林下草本植物优势种的优势度较为明显,优势种与次要优势种相对重要值差值小于5%大于等于2%,物种分布较不均匀;柳杉纯林、水青冈枫香构树混交林、马尾松华山松混交林、朴树女贞混交林、青冈盐麸木混交林、香樟纯林、竹林7 种群落林下草本植物优势种的优势度不明显,排名第一位和第二位的物种相对重要值差值在2%以下,物种分布均匀。可见,在不同林分类型之间,林下草本植物分布的均匀性没有明显规律。在这些群落类型中,出现频次最多的优势种是求米草,其次有芒、毛蕨、狗脊、艾Artemisiaargyi、芒萁、沿阶草Ophiopogon bodinieri等物种。通过不同群落林下草本植物的相对重要值可大致判断乔木层与草本层的相互适应关系,如山麦冬分布在枫香纯林下;冷水花Pileanotata分布在柳杉纯林下;蝴蝶花Irisjaponica分布在水青冈枫香构树混交林、香樟纯林、竹林下,且在香樟纯林中为主要优势种;中国石蒜Lycorischinensis和血水草Eomeconchionantha分布在水青冈枫香构树混交林下;单花红丝线Lyciantheslysimachioides分布在马尾松杉木混交林、马尾松刺槐混交林下;斑叶兰Goodyeraschlechtendaliana、多花黄精、斑叶堇菜分布在青岩油杉纯林下;沿阶草、万寿竹分布在玉兰纯林下。
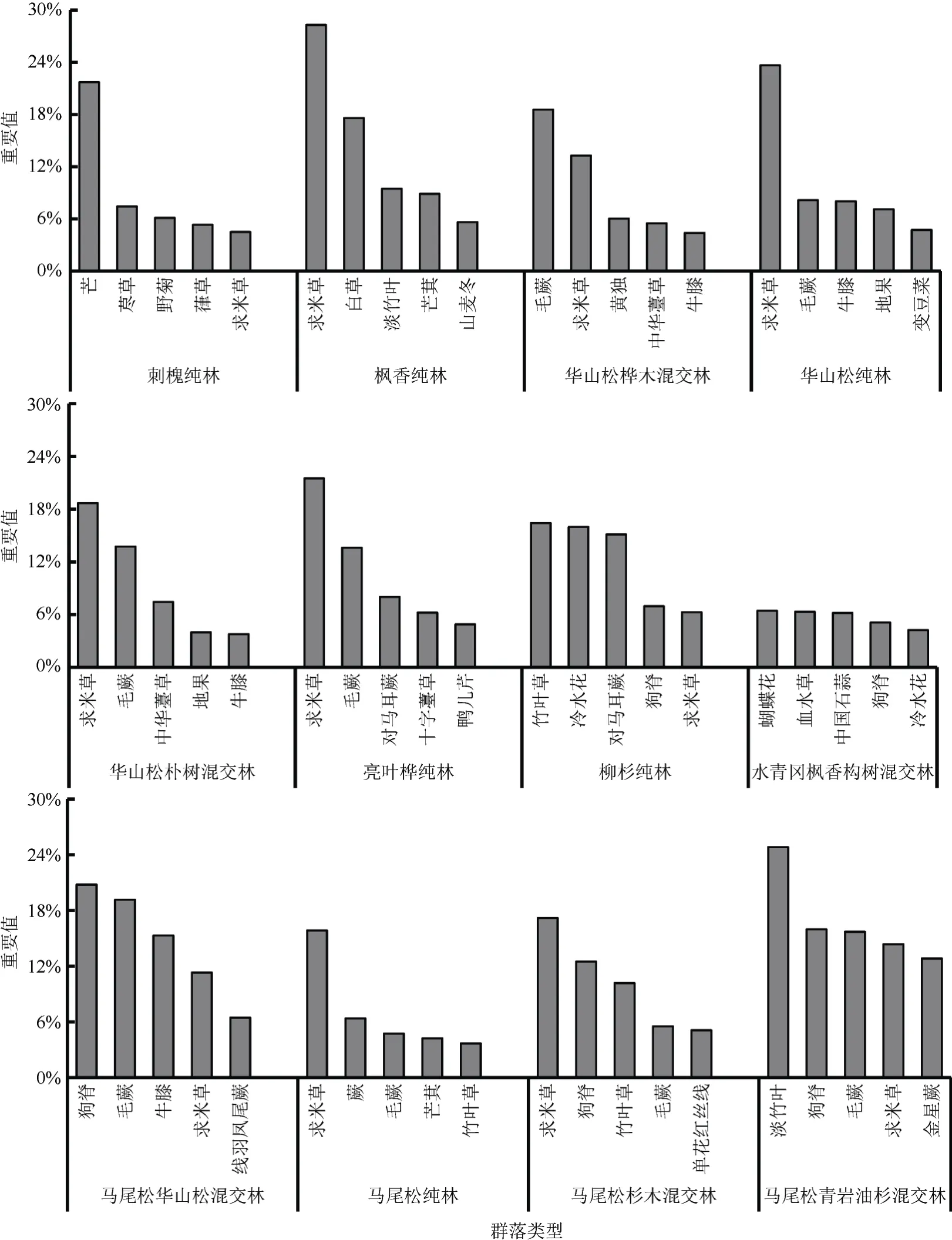
图3(A) 不同群落类型重要值排名前5 位的林下草本植物Fig. 3(A) Top 5 understory herbaceous plants in terms of importance values for different community types
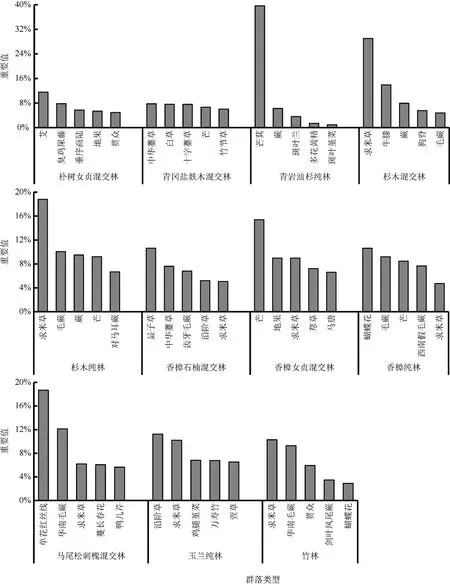
图3(B) 不同群落类型重要值排名前5 位的林下草本植物Fig. 3(B) Top 5 understory herbaceous plants in terms of importance values for different community types
2.3 林下草本植物组成对环境因子的响应
8 个环境因子共解释了贵阳市林下草本植物组成变异量的20.2%,林分类型和海拔是影响林下草本植物分布的主要环境因子(表5)。CCA1 轴从左至右,表示凋落物厚度增加,坡度、坡位上升,坡向由阳坡向阴坡变化,林分类型由针阔叶混交林转变为针叶纯林。说明荩草Arthraxonhispidus、蛇床Cnidiummonnieri、皱叶狗尾草Setariaplicata、血水草Eomeconchionantha等物种对应的环境条件为下坡位、阔叶或针阔叶混交林;芒萁、粗毛淫羊藿Epimediumacuminatum、金星蕨Parathelypteris glanduligera、斑叶堇菜Violavariegata等物种在针叶林、阴坡、郁闭度较高的林下分布较多。CCA2 轴从下至上,海拔、乔木密度降低。野菊Dendranthemaindicum、蜈蚣凤尾蕨Pterisvittata、白茅Imperatacylindrica等物种对应的环境条件为低海拔、低密度林下;而贯众Cyrtomiumfortunei、紫萼Hostaventricosa、唐松草Thalictrumaquilegiifolium等物种与之相反,适宜分布在研究区较高海拔林下(图4)。
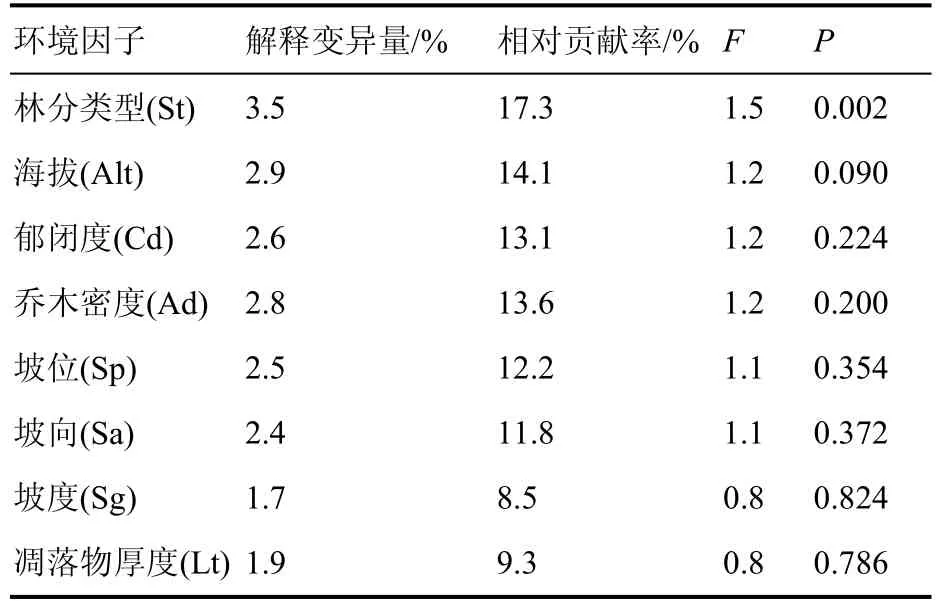
表5 环境因子对物种组成的解释率Table 5 Interpretation rate of CCA environmental factors on species composition
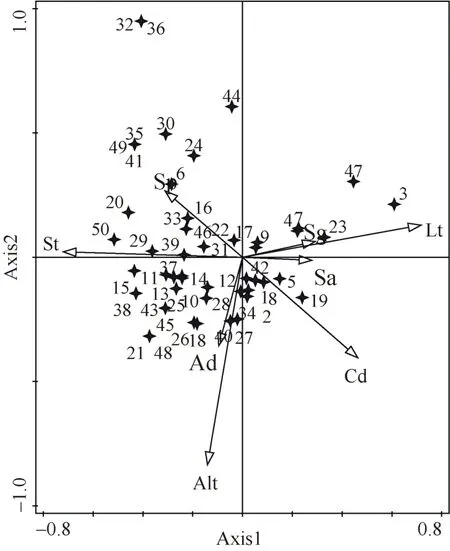
图4 物种与环境因子的典范对应分析排序图Fig. 4 Ranking chart of exemplary correspondence analysis between species and environmental factors
2.4 林下草本植物多样性分析
2.4.1 不同林分类型林下草本植物多样性
不同林分类型之间的林下草本植物多样性指数存在显著差异性(图5)。各林分类型林下草本植物的Margalef 丰富度指数在3.5716~6.3668 之间,其中针阔叶混交林的物种丰富度最高,显著高于针叶林和针叶混交林(P<0.0 5),针叶混交林最低;Shannon-Wiener 指数范围在2.1086~2.8149 之间,用来描述物种的多样性情况,林下草本植物的多样性水平在阔叶混交林、针阔叶混交林显著高于针叶纯林和针叶混交林;Simpson 指数是描述群落中物种的集中性的指标,指数范围为0.7625~0.9004,其中阔叶混交林的最高,显著高于针叶纯林;Pielou指数区间为0.6592~0.7797,说明林下草本植物在不同林分类型之间分布较不均匀,均匀度最高的林分类型是阔叶混交林,其次是阔叶纯林,均显著高于针叶纯林。整体上看,多样性指数表现出混交林高于纯林,阔叶林高于针叶林的规律性。因此,林冠结构的复杂程度会影响林下草本植物的物种多样性格局,林冠结构可能通过调控林下光环境、枯枝落叶层、土壤理化性质等的差异影响林下草本植物的生长和发育。
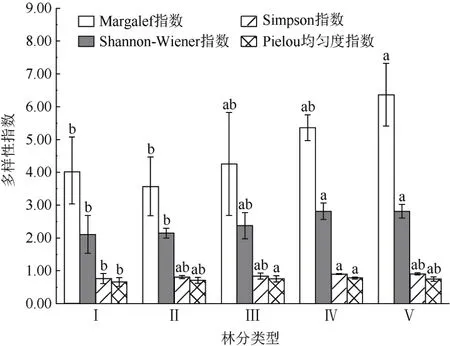
图5 不同林分类型林下草本植物的多样性Fig. 5 Diversity of understory herbaceous plants in different stand types
2.4.2 林下草本植物物种多样性与环境因子的相关性
林下草本植物与环境因子的相关性分析表明,影响林下草本植物生活力的环境因子主要是林分类型、乔木密度、郁闭度和凋落物厚度,影响其多样性的环境因子主要是林分类型、凋落物厚度、坡向和坡位(表6)。生活力与林分类型表现出极显著正相关(0.490,P<0.01),与乔木密度、郁闭度呈极显著负相关(-0.426、-0.503,P<0.01),与凋落物厚度呈显著负相关(-0.312,P<0.05),说明在森林群落中,林下草本植物更趋向于在针阔叶混交林中生长,乔木密度越大、郁闭度越高、凋落物越厚对林下草本植物的健康状况越不利。4 个多样性指数均与林分类型呈极显著正相关(P<0.01),与乔木密度、郁闭度、海拔、坡度相关性不显著。Margalef 指数与坡向表现出极显著负相关(-0.473,P<0.01);Shannon-Wiener指数与凋落物厚度、坡向显著负相关(-0.383、-0.334,P<0.05),与坡位显著正相关(0.299,P<0.05);Simpson指数与凋落物厚度极显著负相关(-0.413,P<0.01),与坡位极显著正相关(0.390,P<0.01);Pielou 指数与凋落物厚度极显著负相关(-0.404,P<0.01),与坡位显著正相关(0.380,P<0.05)。可见,林冠结构越丰富、凋落物越薄、坡向越靠近南向、坡位靠近下坡位时,林下草本植物多样性水平越高。由于森林群落中的草本植物依靠林下土壤汲取养分,而复杂的冠层结构使凋落物的组成更加丰富,且针阔混交林的枯落物相较于针叶纯林更容易被分解,因此富含养分的浅层凋落物更适宜林下草本植物生长。同时,阳坡更易于草本植物获取光照,下坡位由于地表径流冲刷使养分堆积,从而对草本植物多样性和分布格局起到至关重要的作用。

表6 林下草本植物多样性与环境因子的Pearson 相关性系数Table 6 Pearson correlation coefficients between understory herbaceous plant diversity and environmental factors
2.4.3 林下草本植物多样性与环境因子多元线性逐步回归分析
为进一步探究林下草本植物多样性的主要影响因子,对多样性指数与环境因子进行多元线性逐步回归分析,结果显示回归方程显著(表7)。林分类型和坡向能够解释Margalef 指数变异量的29.7%,是影响Margalef 指数的主导因子。影响Shannon-Wiener 指数的主导因子为林分类型,解释变异量为27%。Simpson 指数主要受林分类型、坡位影响,二者能够解释Simpson 指数变异量的28.7%。影响Pielou 指数的主导因子为凋落物厚度、坡位、海拔,三者共同解释Pielou 指数变异量的29.6%。
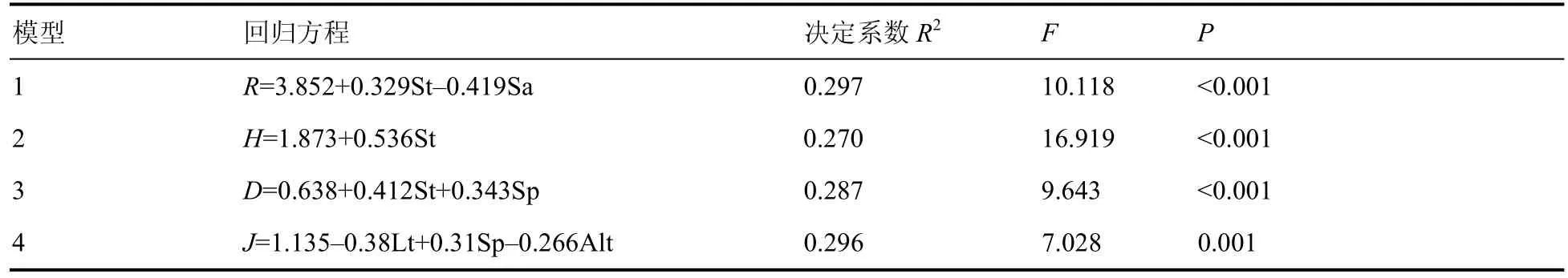
表7 林下草本植物物种多样性与环境因子的多元回归方程Table 7 Multivariate regression equation between species diversity of undergrowth herbs and environmental factors
3 讨论
3.1 林下草本植物分布与环境因子的关系
近年来,关于草本植物多样性的研究多集中在草地生态系统、人工林等生境的调查,对城市森林生态系统中的林下草本植物研究还相对较少。贵阳市作为首个“国家森林城市”,主要依靠城市森林维持物种多样性和群落稳定性。本研究中,林下草本植物优势科为菊科、禾本科、百合科、唇形科,科组成与贵州喀斯特山地草地植物特征一致[31],具有明显的热带区系性质。菊科包含的物种数最多,但禾本科植物的优势度最高;由于菊科植物适宜生长在林缘,物种丰富度虽然高,但在密林中生长状况不佳,所以相对重要值在群落中所占的比例较低;而禾本科的草本植物如求米草、淡竹叶等喜温暖阴湿,因此在森林群落中生长状况良好。
物种分布与环境因子的关系具有重要应用价值,目前被普遍认同的观点是,在全球或区域等大尺度上,地带性气候条件主导植被分布[32];在景观及群落等小尺度上,林分类型、冠层结构、生境、地形、光照、土壤养分等非地带性环境因子是决定植被分布格局的主导因素[33]。由于不同林分的物种组成不尽相同,其冠层结构和土壤理化性质存在差异,通过调控水分和温度影响着林下植物的分布[34—36]。地形在一定程度上能间接反映土壤性质的空间差异[37],尤其在山区,坡向、坡位等小地形因子控制着地表光、热、水等生态环境条件,从而主导了植被物种组成和分布格局的变异情况[38]。地形因子对物种组成的影响有不同的研究结果,在中国东北部分地区,海拔是区域尺度内影响植物分布格局的主要地形因子[39];也有研究认为,在河北地区,坡向相较于海拔对植物分布格局影响更大[40];还有研究表明在广西地区,植被差异是海拔和坡向共同作用的结果[41]。本研究中林分类型和海拔是影响林下草本植物组成和分布的主导环境因子,这与华南地区相关研究结论一致[42],说明物种组成特征是林分因子和地形因子共同作用的结果,同时表明海拔这一地形因子在解释物种组成变异中的重要地位。可见在不同区域,植物分布存在差异,因此要探究影响林下草本植物分布的主要环境因子需要结合研究区的实际情况。
3.2 林下草本植物物种多样性与环境因子的关系
林下草本植物物种多样性的维持机制始终是生态学研究的核心和热点之一,明确其主要影响因素对森林生态系统可持续发展具有重要意义[43]。相较于前人的研究,本研究综合考虑了林分因子、土壤因子、地形因子对林下草本植物多样性的影响,结果表明影响林下草本植物物种多样性的环境因子为林分类型、凋落物厚度、坡向、坡位;多元回归方程表明林分类型、凋落物厚度、坡向、坡位、海拔是影响多样性指数变化的主要环境因子,验证了在群落的尺度上非地带性因子对物种多样性的影响[44—45]。
林冠结构是森林中各层次植物组成的复杂三维空间结构,一定程度上被看作是林地生境异质性的潜在来源[46]。相关研究发现,针阔混交林相较于单树种纯林结构更有利于提高土壤养分含量、改善林内微环境,其林冠结构更复杂,林地更新潜力更高,能充分利用空间资源,通常具有最高的物种多样性、更强的抗逆性和更好的生态系统稳定性[47—48]。本研究结果与上述结论一致,不同林分类型之间草本植物多样性存在显著差异,针阔叶混交林、阔叶混交林草本植物的多样性指数高于针叶林和针叶混交林,阔叶林处于中等水平,林分类型与多样性指数的相关性系数最高,被纳入回归方程的次数也最多。但乔木密度和郁闭度与林下草本植物多样性相关性不显著,乔木密度和郁闭度是群落演替阶段、林龄、植物特性、人为干扰等因素互相作用的结果,直接关系到群落光环境特征[49]。由此可见,贵阳市城市森林林下光环境比较稳定,林下草本植物的多样性水平主要由林分类型决定。
土壤是植物生长必备因素之一[50]。土壤因子对物种多样性影响的研究大多关注土壤理化性质[51]。而本研究以凋落物厚度作为土壤因子,强调了凋落物影响林下草本植物多样性过程中的物理作用。凋落物厚度增加对Shannon-Wiener 指数、Simpson 指数、Pielou 指数存在显著负向作用,凋落物越厚,养分含量越多,却不利于林下植被的生长发育。因为凋落物可通过物理阻断和化感作用影响幼苗出土和早期生长,且短期内凋落物的覆盖主要是物理作用[52]。适当清理林下凋落物,短期内可提高表层土壤养分供应能力,促进群落天然更新[53—54]。不同的是,陈玉婷等[40]针对栎皮林群落的研究表明枯落物厚度对物种丰富度存在显著正向影响。由于不同群落凋落物厚度存在差异,在凋落物较薄的群落,适宜凋落物厚度对物种多样性有一定促进作用,而在凋落物较厚的群落,物种多样性则受到一定限制。说明,凋落物厚度增加对林下草本植物多样性的影响不是单纯的促进或抑制作用,而是适宜的浅层凋落物最有利于林下草本植物生长发育。
地形差异形成的生境特征是物种多样性维持的重要基础。本研究中,Margalef 指数和Shannon-Wiener 指数与坡向显著负相关,多元线性回归方程表明阳坡、下坡位的物种多样性较高,反之则较低。相关研究的解释为:海拔、坡度、坡向、坡位等地形因子间接影响山地水、热、土壤、光照条件,被认为是山地植物的生长、分布及多样性格局的显著影响因子[55—56]。坡向影响地表接受的太阳辐射量[57],坡位则通过对太阳辐射的调节和降雨的分配[58],从而影响土壤温度、含水量、风化作用,土壤水肥条件,最终影响植物多样性的差异。贵阳市森林群落海拔高差较小,林下草本植物多样性随地形因子变化而改变说明了在海拔梯度较小的山地城市,坡向、坡位是影响林下草本植物多样性分布格局的主要地形因子。
4 结论
贵阳市城市森林林下草本植物分布与多样性格局是林分因子、土壤因子、地形因子共同作用的结果。贵阳市城市森林群落物种多样性提升可以针对林分类型、土壤凋落物厚度、坡向、坡位的差异制定方案。例如乡土树种混交种植以增加林分稳定性,保留适宜厚度的凋落物促进草本植物生长,在不同坡向适当引入喜荫、耐荫草本植物以提升物种多样性。若要更全面地解释生态系统物种组成与多样性分布格局及其影响因素,还应综合考虑人为干扰、土壤因子、生境异质性以及种子传播等各种因素。本研究能够为物种多样性与环境因子关系的深入研究提供理论基础,为贵州地区城市森林植物多样性保护与提升提供相应的应用指导。
附录:贵阳城市森林群落林下草本维管植物名录
(见网站:http://www.yrdzwkx.com/CN/10.3969/j.issn.1009-7791.2023.05.009)
