中华绒螯蟹幼体的栖息生境选择与行为特征
2024-01-22伍烨菱王思凯李春波
伍烨菱, 王思凯, 赵 峰,3*, 李春波, 庄 平,3
(1. 上海海洋大学,水产科学国家级实验教学示范中心,上海 201306;2. 中国水产科学研究院东海水产研究所,农业农村部东海渔业资源开发利用重点实验室,上海 200090;3. 上海长江口渔业资源增殖与生态修复工程技术研究中心,上海 200090)
自然界中,动物具有趋利避害的本能,总是趋向于选择最有利于其生存和繁衍的环境作为栖息地[1]。栖息地,尤其是早期发育阶段的栖息地,生境条件适宜与否将直接影响幼体存活率[2]、变态发育[3-4]、生物量补充[5-6]及其资源分布[7-8]。因此,研究动物早期发育阶段的栖息地与生境需求不仅可以丰富其基础生物学资料,而且对于资源保护利用及其栖息地生态修复均具有重要意义。
虾蟹类水生动物早期发育阶段普遍存在着从营浮游生活至营底栖生活的转变过程,通常发生在后期大眼幼体阶段。此时,若栖息生境适宜,后期大眼幼体将顺利沉降,从而进一步生长发育,反之,将继续维持浮游状态寻找合适的栖息生境。研究表明,蓝蟹(Callinectes sapidus)后期大眼幼体在发现海草床生境后开始沉降定居,并快速发育至Ⅰ期仔蟹[9]。虾蟹类随着幼体发育,栖息生境的选择也会发生变化。Webley等[10]发现锯缘青蟹(Scylla serrata)大眼幼体在泥、沙、水草中均匀分布,而Ⅰ期仔蟹主要分布在水草中。蓝蟹大眼幼体和早期幼蟹主要栖息在海草床生境,而头胸甲宽20~30 mm的幼蟹则开始往沙、泥生境中迁移[9]。Richards[11]发现大部分中小型北方黄道蟹(Cancer borealis)在鹅卵石底质栖息,而大型北方黄道蟹在鹅卵石和沙子中均有分布。
中华绒螯蟹(Eriocheir sinensis)俗称河蟹,是我国重要的渔业对象和淡水养殖蟹类,具有很高的商业和生态价值[12]。长江口是长江水系中华绒螯蟹的唯一繁育场,其交配产卵、孵化和早期发育均在长江口完成。因此,长江口的栖息生境对于中华绒螯蟹的迁移分布、早期发育、资源补充等具有重要影响。目前,对于自然条件下中华绒螯蟹幼体在长江口的栖息生境选择尚无系统研究,仅在养殖上发现,种植水草有利于提高中华绒螯蟹大眼幼体的蜕壳率和存活率[13-14]。本研究以长江口微生境类型为基础,通过模拟藨草(Scirpus triqueter)、泥底、铁板沙和砾石等4种生境,研究中华绒螯蟹早期发育阶段对不同生境的选择利用及其行为特征,旨在丰富中华绒螯蟹早期生活史资料,并为其资源保护和栖息地生态修复提供基础参考。
1 材料与方法
1.1 实验材料
中华绒螯蟹大眼幼体[头胸甲宽(CW)=(1.5±0.2) mm]取自如东养殖基地,运回实验室进行养育。通过详细观察与记录幼体的蜕壳次数,结合刘启彬[13]的仔蟹生长参数,挑选出Ⅰ期仔蟹[头胸甲长(CL)=(3.1±0.3) mm]、Ⅲ期仔蟹[CL=(5.0±0.5)mm]、Ⅴ期仔蟹[CL=(7.6±0.7) mm]。养殖过程中保持自然光照,每日投喂冷冻卤虫,充氧、吸污、换水,水温维持在18.5~22.1 °C,溶解氧含量为6.8~7.2 mg /L,pH值为7.5~7.9。实验过程中操作人员严格遵守东海水产研究所动物保护和使用伦理规范,并按照东海水产研究所动物保护和使用伦理委员会制定的规章制度执行。
1.2 实验方法
生境选择实验 根据长江口自然生境特征,分别模拟藨草生境、铁板沙生境、泥底生境和砾石生境进行选择实验。以红色塑料盆(底径635 mm,高170 mm)作为实验装置,使用亚克力板制作“四等分板”将红色塑料盆均分为4等分区块,分别作为模拟生境区域。在其中的3个区块中,分别将取自长江口东滩的铁板沙、泥和砾石[(5.0±2.5) mm]均匀铺在盆底,厚度30 mm,作为铁板沙生境、泥底生境和砾石生境;最后1个区块中,以泥底生境为基础均匀插入藨草[茎长(141.0±12.2) mm,茎直径(2.0±1.2) mm],密度约1 000 棵/m2,作为藨草生境。为消除生境位置对选择偏好的影响,分别设置4种生境放置方式(图1)。开始实验前,将实验装置中“四等分板”移走,加入16 L曝气淡水。在首日23:00时,在实验装置的中心位置放入30只体型相似且肢体完整的相同发育时期幼体,次日分昼(10:00)和夜(22:00)各记录1次幼体在不同生境中的分布数量。观察记录前,先将“四等分板”插入装置,以防止幼体在不同生境间移动,从而影响记数。夜间计数时,使用波长620~625 nm红光灯(中华绒螯蟹幼体对红光不敏感)辅助操作。实验设3个平行,共进行3 d。
图1 实验装置中模拟生境放置方式Fig. 1 Placement of simulated habitats in the experimental device
行为观察实验 以红色塑料盆作为实验装置,整个装置仅填入藨草生境。实验开始首日23:00时,在实验装置的中心位置放入30只体型相似且肢体完整的相同发育时期幼体,次日分昼(10:00)和夜(22:00)各记录1次幼体在生境中存在的所有行为。每日更换60%曝气水。实验设3个平行,共进行3 d。
死亡率和蜕壳率实验 选取肢体完整的Ⅰ~Ⅱ期仔蟹(CL=2.5~4.0 mm)作为实验对象,按照30只/5 mL冷冻卤虫比例,每日10:00进行投喂,暂养3 d。暂养期间,卤虫均有剩余。实验设立投喂组和未投喂组。两组实验各使用3个红色塑料盆作为实验装置,各组每个装置填入藨草、泥、空盆中的一种生境。每个实验装置均加入16 L曝气水,并投放Ⅰ、Ⅱ期仔蟹各15只。投喂组每日10:00投喂5 mL卤虫,未投喂组不进行投喂。实验共进行5 d。实验中不曝气,每日投喂前更换60%~70%曝气水,并尽可能清除食物残渣。实验设置3个平行。早晚各观察1次生境中散落的蟹尸体残骸、头胸甲壳以及存活幼体数量,以判断幼体蜕壳或死亡,如发现1个头胸甲壳,幼体剩余数与昨日观察数一致,判断为蜕壳;若幼体剩余数比昨日观察数减少1只,判断为死亡。
1.3 数据分析
使用IBM SPSS Statistics 26软件进行数据分析。利用Kruskal-Wallis检验及多重比较(经Bonferroni校正)分析幼体在生境中分布数量的差异;皮尔森卡方检验(Pearson's chi-squared test)分析生境布置方式对幼体分布的影响;Wilcoxon符号秩检验分析幼体在昼夜的分布和行为差异;单因素方差分析(One-Way ANOVA)及多重比较(S-N-K)分析幼体在藨草生境中的行为变化。双因素方差分析(Two-Way ANOVA)及多重比较(S-N-K)分析幼体在不同生境和投喂条件的死亡率、蜕壳率差异。单因素方差分析、双因素方差分析,均通过莱文(Levene)方差齐性检验。显著性水平均设为 0.05。
2 结果
2.1 中华绒螯蟹幼体的生境选择
四种生境布置方式对于中华绒螯蟹大眼幼体、Ⅰ、Ⅲ、Ⅴ期仔蟹(以下简称“四个发育时期”)的分布比例均无显著影响(皮尔森卡方检验,大眼幼体组χ2=1.808、Ⅰ期仔蟹组χ2=2.135、Ⅲ期仔蟹组χ2=15.010、Ⅴ期仔蟹组χ2=2.353,P>0.05)。昼夜差异对四个发育时期仔蟹的分布比例无显著影响(Wilcoxon符号秩检验,大眼幼体组Z> −1.155、Ⅰ期仔蟹组Z> −0.307、Ⅲ期仔蟹组Z> −1.873、Ⅴ期仔蟹组Z> −1.222,P>0.05)。
四个发育时期幼体对生境的选择结果如图2所示,四个发育时期在四个生境中分布量均有显著差异(Kruskal-Wallis 检验,大眼幼体组H=59.382、Ⅰ期仔蟹组H=55.551、Ⅲ期仔蟹组H=75.136、Ⅴ期仔蟹组H=79.110,P<0.05。“H”值为Kruskal-Wallis 检验的检验统计量)。经两两比较,四个发育时期在藨草生境的分布比例(大眼幼体73.7%±6.8%、Ⅰ期仔蟹 78.2%±8.8%、Ⅲ期仔蟹53.6%±8.0%、Ⅴ期仔蟹 54.8%±6.8%)均显著高于其他三种生境(P<0.05)。大眼幼体和Ⅰ期仔蟹在泥、铁板沙、砾石生境中的分布比例无显著差异(P>0.05)。Ⅲ、Ⅴ期仔蟹在泥生境中的分布比例分别为23.7%±6.6%和23.6%±5.3%,均显著高于砾石和铁板沙生境(P<0.05)。
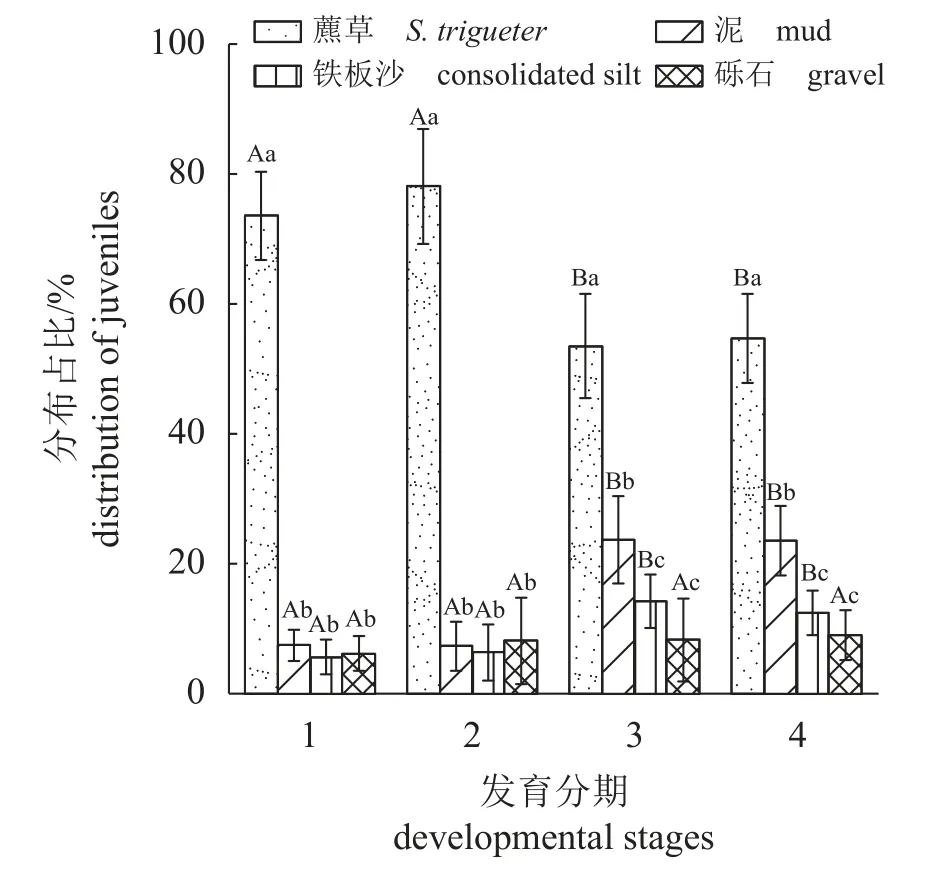
图2 中华绒螯蟹幼体在不同生境的分布比例1.大眼幼体,2. Ⅰ期仔蟹,3. Ⅲ期仔蟹,4. Ⅴ期仔蟹。不同大写字母表示组间差异显著 (P<0.05),不同小写字母表示组内差异显著(P<0.05),下同。Fig. 2 Distribution ratio of larva of E. sinensis in different habitats1. megalopa, 2. juvenile (stage Ⅰ), 3. juvenile (stage Ⅲ), 4. juvenile(stage Ⅴ). Different capital letters indicate significant difference between groups (P<0.05), and different lowercase letters indicate significant difference within groups (P<0.05), the same below.
从不同发育分期在生境的分布情况来看,Ⅲ、Ⅴ期仔蟹在藨草生境的分布比例显著低于大眼幼体和Ⅰ期仔蟹(P<0.05),在泥、铁板沙生境的占比显著高于大眼幼体和Ⅰ期仔蟹(P<0.05)(图2)。
2.2 藨草生境下中华绒螯蟹幼体的行为特征
结果显示,幼体在藨草生境中主要有游泳(在水中游动)、附着(附着在藨草茎上)、表栖(趴在泥上,头胸甲全部暴露在水中)、埋栖(头胸甲一半以上埋在底质中)行为。昼夜差异对幼体行为无显著影响(Wilcoxon符号秩检验,大眼幼体组Z> −1.653、Ⅰ期仔蟹组Z> −1.951、Ⅲ期仔蟹组Z> −1.310,P>0.05)。大眼幼体、Ⅰ期仔蟹主要表现出附着行为,分别占82.1%±5.1%、83.3%±8.7%,仅少量大眼幼体表现出游泳行为(9.0%±3.7%)。至Ⅲ期仔蟹,附着率(19.3%±6.4%)显著减少,表栖率(44.2%±17.9%)、埋栖率(36.5%±16.7%)显著增加(P<0.05) (图3)。此外,由于Ⅴ期仔蟹活动能力较强,使得盆中水过于混浊,难以进行观察,因此Ⅴ期仔蟹行为实验无结果。
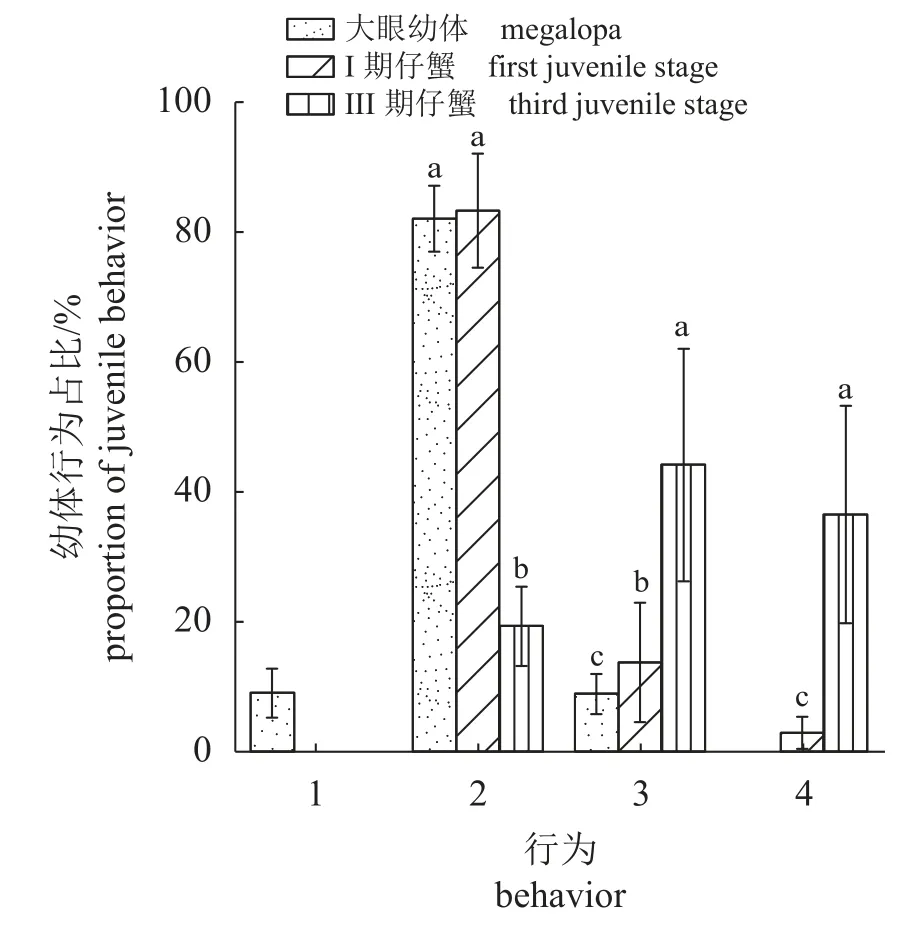
图3 中华绒螯蟹幼体在藨草生境中的行为变化1. 游泳,2. 附着,3. 表栖,4. 埋栖。Fig. 3 Behavior changes of larva of E. sinensis in S. mariqueter habitat1. swimming, 2. clinging, 3. epibiosis, 4. burying.
2.3 不同生境中幼体死亡率和蜕壳率
幼体在5 d内的蜕壳率、死亡率百分比累计显示,投喂、生境对幼体的蜕壳率影响显著(投喂:F=166.481,P<0.05,生境:F=198.092,P<0.05),二者的交互作用显著(F=21.843,P<0.05)。简单效应分析显示,投喂组藨草中幼体蜕壳率(86.7%±6.9%)显著高于泥和空盆(P<0.05),空盆中幼体蜕壳(64.4%±5.1%)率显著高于泥(46.6%±8.8%) (P<0.05)。未投喂组藨草中幼体蜕壳率(81.1%±5.1%)显著高于泥、空盆(P<0.05),泥(23.3%±10.1%)与空盆(24.4%±3.8%)的幼体蜕壳率无显著差异(P>0.05)。投喂与否对藨草中幼体蜕壳率无显著影响(P>0.05),对泥、空盆的幼体蜕壳率影响显著(P<0.05) (图4)。

图4 中华绒螯蟹幼体在不同生境中蜕壳率、死亡率百分比累计Fig. 4 Accumulation of molting rate and mortality rate of juvenile of E. sinensis in different habitats
生境对幼体的死亡率影响显著(F=27.161,P<0.05),投喂对幼体的死亡率无显著影响(F=0.221,P>0.05),二者的交互作用不显著(F=1.721,P>0.05)。经两两比较,投喂组泥生境中幼体死亡率(24.4%±1.9%)显著高于藨草和空盆,藨草(13.3%±3.3%)和空盆(14.4%±1.9%)生境幼体死亡率无显著差异。在未投喂组泥生境中幼体死亡率(26.6%±5.7%)显著高于藨草和空盆,空盆中幼体死亡率(17.7%±1.9%)显著高于藨草(10.0%±3.3%)。
3 讨论
虾蟹类早期发育阶段在沉降时期识别并选择合适生境,有利于提高生存机会、保证生物量补充成功[15]。本研究中,中华绒螯蟹大眼幼体在各生境分布的比例不均匀,表明大眼幼体有识别实验中四种生境的能力。大眼幼体在藨草生境中的分布比例显著高于其他生境,可能是藨草生境有着更复杂的三维构造,可作为庇护场所,有利于其发育至底栖阶段。Stevens 等[16]认为选择复杂构造栖息地是幼体躲避捕食者的适应性行为,目前已有较多研究发现蟹类大眼幼体偏好沉降定居在具有复杂构造的栖息地。如Moksnes[7]研究发现,岸蟹(Carcinus maenas)大眼幼体明显偏好贝类、藻类和大叶藻(Zostera marina)斑块等复杂构造的栖息生境。Van Montfrans等[6]和Moksnes[17]等研究发现,栖息在大叶藻中的蓝蟹大眼幼体数量是裸露砂质环境的2倍。
随着幼体的成长,特定栖息地对幼体生长、生存的益处也会发生变化[17-18],可能导致幼体迁出原栖息地。本研究中,Ⅰ期仔蟹对比大眼幼体的分布情况无显著差异,均有80%以上的幼体分布在藨草中。而Ⅲ、Ⅴ期仔蟹在藨草生境的数量显著降低,仅剩50%左右,在泥、铁板沙生境中占比增大,栖息比例显著高于大眼幼体和Ⅰ期仔蟹。诸多研究表明,这种现象,主要由两种机制驱动。第一,随着幼体体型增大,原生境中适合幼体体型的藏匿空间减少[19-20],幼体开始重新寻找适合自身体型的藏匿空间。例如在Tapella等[21]的研究中,短足拟石蟹(Paralithodes platypus)大眼幼体在鹅卵石、破碎的贝壳和砾石中均匀分布,而Ⅰ期仔蟹体形更大,主要分布在间隙更大的贝壳和鹅卵石中。第二,成长与生存之间的权衡[22]。随着幼体体型增大,幼体能更好地躲避或攻击捕食者,为获取更高能量食物维持成长和活动,幼体将主动开发其他的无庇护所栖息地。例如美洲螯龙虾[23](Homarus americanus)幼体会根据自身个体大小,以及育幼场生境的食物供给情况,决定是否前往无庇护所的生境觅食。本研究中Ⅲ、Ⅴ期仔蟹栖息比例变化的原因,可能与其食物需求或躲避攻击的能力增强有关。一方面,崔莹等[24]通过对野外采集的幼体进行食性分析,发现大眼幼体和早期仔蟹主要以浮游动物为食,而Ⅲ~Ⅴ期仔蟹则开始摄食底栖微藻、碎屑和底栖动物。徐德崑等[25]发现,Ⅱ期仔蟹的食量显著增加,虽喜食植物性饵料,但更喜食肉食性饲料。因此可以推测,Ⅲ期仔蟹摄食需求发生了变化,导致部分幼体开始探寻其他生境。另一方面,本研究中Ⅲ期仔蟹无论在单一藨草生境,还是在选择实验中,其埋栖率显著高于Ⅰ期仔蟹。而埋栖行为作为幼体躲避捕食者的重要策略[26],表明Ⅲ期仔蟹展现出了更强的躲避捕食者能力。但是,藨草生境仍是Ⅲ、Ⅴ期仔蟹分布最多的生境,表明此阶段藨草生境仍是其最重要的栖息场所。
通过对比不同生境中幼体的死亡率和蜕壳率,发现藨草生境对幼体的生存和生长是有益的。无论投喂与否,藨草生境中幼体死亡率显著低于除投喂组空盆的其他生境。这可能是因为藨草生境为幼蟹提供了隐蔽物,减少了幼蟹的同类残食现象。例如在刘启彬[13]的研究中,中华绒螯蟹Ⅰ~Ⅲ期仔蟹在无遮蔽环境中的存活率显著低于有空心莲子草(Alternanthera philoxeroides)的环境,作者认为空心莲子草有效减少了同类残食现象,与本研究结果类似。泥生境中幼体的累计死亡率较高,一方面可能是由于缺少庇护场所,幼体间易发生打斗,蜕壳时易受到同类攻击;另一方面,由于底泥是从野外环境中直接取回,其中可能存在少量底栖动植物碎屑,这些物质释放的信号可能会增加幼蟹的觅食活动,使幼体间为争夺有限资源而发生更多打斗[27-28],导致高死亡率。实验过程中,藨草生境中水质浊度较低,泥生境水质有一定浊度,这可能是泥生境中幼体间频繁发生打斗的结果。此外,无论投喂与否,藨草生境的幼体蜕壳率均显著高于其他生境。一方面,甲壳类的周期性生理蜕壳生长需要一定的能量积累与储备[29],当饵料不足,营养积累不够充分,蜕壳间期时间会延长,蜕壳增长率下降;另一方面,虾蟹类幼体在蜕壳时行动能力弱,最容易遭受攻击,良好的庇护空间有助于避开攻击,提升蜕壳成功率[30];此外,泥生境中更多的争斗行为可能会消耗更多能量,甚至导致断肢,而延长蜕壳周期[31]。因此,较多研究认为,有植被覆盖的栖息地为幼体提供了庇护所和充足的食物,有助于虾蟹类幼体的生长。而在无植被生境(即泥或空盆)中,存在缺少适宜的蜕壳环境、频繁的打斗等问题,可能会延长仔蟹的蜕壳周期。例如,Perkins-Visser等[4]、Ryer等[32]发现,在野外植被覆盖的栖息地(海草床)中的蓝蟹幼蟹,相比无植被覆盖的泥滩有更高的生长率。徐德觉等[25]发现,中华绒螯蟹幼蟹蜕皮所需时间与放养密度、饵料和附着物等有关,若水草作为附着物,幼蟹蜕皮增长率更高。刘庆华[14]的研究表明,在池塘网笼中,底泥环境的中华绒螯蟹扣蟹蜕壳周期相比含有伊乐藻(Elodea canadensis)的环境显著延后。
综上所述,中华绒螯蟹幼体的生境选择行为,是与自身生存和发育需求相适应的。幼体为躲避攻击,栖息于更佳的生存环境中,大部分选择结构复杂的藨草生境。虽然部分Ⅲ、Ⅴ期仔蟹开始往泥、铁板沙中迁移,但藨草生境仍是Ⅲ、Ⅴ期仔蟹分布比例最高的栖息生境。
(作者声明本文无实际或潜在的利益冲突)
