长牡蛎壳橙性状遗传参数评估及与生长性状的关联性
2024-01-22陈雨露徐成勋刘士凯孔令锋
陈雨露, 徐成勋, 刘士凯, 孔令锋, 李 琪,2*
(1. 中国海洋大学,海水养殖教育部重点实验室,山东 青岛 266003;2. 青岛海洋科学与技术试点国家实验室,海洋渔业科学与食物产出过程功能实验室,山东 青岛 266237)
长牡蛎(Crassostrea gigas)又称太平洋牡蛎,因其繁殖力高、环境适应能力强、养殖周期短、营养丰富等特点,已成为世界上最为重要的海产经济贝类,其产量连续多年高居世界之首。作为我国商业化利用程度高的贝类养殖品种,2020年牡蛎全国海水养殖产量约为542万t[1]。目前,我国长牡蛎养殖所用苗种主要来源于人工育苗,培育生产性能优良、品质与品相兼备的新品种,对进一步提升牡蛎的产值,促进牡蛎产业的高质量发展尤为重要。
壳色作为海产经济贝类的外观性状,影响着消费者的喜好,在提升贝类附加值方面发挥着重要的作用。壳色性状的选择育种和遗传学研究一直受到育种工作者的关注[2-4],国内已相继培育出长牡蛎、栉孔扇贝(Azumapecten farreri)、菲律宾蛤仔(Ruditapes philippinarum)和文蛤(Meretrix meretrix)等[5-8]壳色新品种,推动了我国贝类养殖产业的良种化。在长期的牡蛎育种实践中,已获得了长牡蛎橙色壳色突变体品系,通过品系间杂交和群体选育开发出长牡蛎壳橙快速生长优良品系,为长牡蛎新品种培育提供了重要素材。
经济性状的遗传参数评估是水产动物遗传改良过程中的一项基础工作。其中,狭义遗传力是加性方差在表型方差中所占的比例,有助于判断亲本性状遗传给后代大小的程度,性状间的遗传相关性则是多性状选育方案的制定依据[9]。在长牡蛎的育种中,与经济价值紧密相关的生长性状、存活率、抗逆性、壳型、颜色等遗传参数评估相继报道,并以目标性状遗传力和不同性状间的关联性为依据,制定了育种计划[10-14]。同一性状的遗传参数因生长环境、品系而异,因此,在新品系的继代选育过程,对特定性状的遗传参数进行评估,探索其作为目标性状的选育潜力,是确保后续选育工作取得改良效果的重要前提。
本研究以长牡蛎壳橙快速生长品系为素材,通过巢式交配设计建立了48个全同胞混养家系,利用微卫星亲子鉴定技术获取系谱信息,采用计算机视觉系统和Lab颜色模型获取壳色数据,估算壳橙性状遗传力及其与生长性状的关联性,以期为长牡蛎壳橙快速生长品系选育提供基础资料。
1 材料与方法
1.1 实验设计
实验所用的亲本是经过连续3代选育、生长性状得到改良的橙壳色长牡蛎个体,即第1代长牡蛎壳橙快速生长品系(G1)。2020年4月从山东乳山海区养殖的第1代长牡蛎壳橙快速生长品系(G1)中选取生长性状优良个体作为亲贝,转移至室内进行人工促熟。2020年5月,长牡蛎亲本性腺发育成熟,辨别雌雄后,利用平衡巢式设计法进行人工授精,1雄配3雌,共建立48个全同胞家系、16个半同胞家系。获取所有亲本的闭壳肌,保存于70%酒精中。
1.2 孵化、苗种培育和养成
人工授精完成后,所有家系幼虫在100 L聚乙烯桶内进行孵化,孵化过程中幼虫密度需保持在20~40个/mL,水温控制在24oC左右。孵化过程持续22 h,受精卵发育至D形幼虫期时进行选优,从48个家系中分别取幼虫密度为30~40个/mL的2 L养殖水体并转移至2个500 L塑料桶中混合培育。苗种培育与养成参照王庆志等[15]的方法,幼虫密度控制在8~10个/mL,至投放附着基时调整至2~3个/mL。每日换水1次,每次1/3换水量;每日投喂4次,根据不同生长阶段调整等鞭金藻(Isochrysis galbana)和扁藻(Platymonassp.)的投喂比例及投喂量。两个聚乙烯桶的管理操作基本保持一致。当眼点幼虫比例达30%左右时,投放扇贝壳附着基采苗,幼虫完成附着和变态后,转移至室外沉淀池暂养2周。待自然海区野生牡蛎减少时,将稚贝置于扇贝笼中转移至山东荣成桑沟湾海区进行养成。养成期间各笼内的放养密度和所处水层均保持一致。2021年4月从长牡蛎家系混养群体中共收获863个个体。本研究按照中国海洋大学《实验动物使用指南》的标准操作程序进行。所有动物护理和使用程序均经中国海洋大学动物机构护理和使用委员会批准。
1.3 指标测定
生长性状测定 对所有个体的壳高(SH)、壳长(SL)、壳宽(SW)、总重量(TW)等生长性状指标进行测定。用电子游标卡尺测量壳高、壳长和壳宽(精确度0.01 mm),总重量用电子天平测量(精确度为0.01 g)。
壳色性状测量 采用计算机视觉系统(CVS, computer vision system)和均匀颜色空间L*a*b*采集、量化壳色数据。CVS系统包括标准光源、数码相机和图像处理软件。壳色图像采集过程、相机参数设置以及颜色参数L、a、b值的读取参照Wan等[16]的方法。L*、a*、b*和ΔE通过以下公式计算[17-18]:
式中,L*表示明度,a*为红绿轴色品指数,b*为黄蓝轴色品指数,L、a、b分别为所测牡蛎壳的颜色参数,L*1、a*1、和b*1是子代中某一标准壳橙个体的颜色指标,ΔE为色差值。
1.4 家系鉴定
所有雌雄亲本以及子代的闭壳肌使用苯酚-氯仿法[19]提取DNA。利用4组多重PCR共12个微卫星位点对所有样品的DNA进行扩增,多重PCR扩增的组合、引物序列和PCR扩增条件见表1。所得的扩增产物用ABI3730xl (Applied Biosystems)测序仪进行基因分型。经GeneMapper v4.0软件处理得到相应位点的片段大小,使用CERVUS 3.0软件[20],基于最大似然法,结合巢式设计的原则和12个微卫星位点提供的信息,鉴定亲缘关系。

表1 长牡蛎微卫星多重PCR引物组合及特征Tab. 1 Multiplexes of microsatellite markers for C. gigas
1.5 数据分析
使用SPSS 18.0和Excel软件对长牡蛎子代生长性状和壳色性状的表型参数进行初步处理,检测正态性和方差齐性。采用ASREML 3.0程序包中约束极大似然法(REML)进行遗传参数评估,建立个体动物模型:
式中,i、j、k依次为父本编号、母本编号、家系中第k个体的编号,Yijk为个体性状观测值,μ为总体平均值,αijk为加性遗传效应,Iij为家系遗传效应,eijk为随机残差。
壳色指标和生长性状的遗传相关和表型相关的计算,使用ASREML 3.0软件以及双变量模型。
2 结果
2.1 系谱鉴定及描述性统计
12个微卫星位点的平均等位基因数为10.6,平均观测杂合度为0.62,平均期望杂合度为0.67,多态性信息含量(PIC)的平均值为0.62,表现出高多态性。进行10 000次亲本已知情况下的模拟测试,鉴定准确率为99.98%。在95%的置信度标准下,863个个体中有851个个体鉴定到其父母本,实际亲子鉴定成功率为98.61%。家系重建结果显示,48个家系之间的子代个体数存在较大差异(表2),介于6~48。亲本贡献率最高的是2号父本和4号母本,共繁育了48个子代。
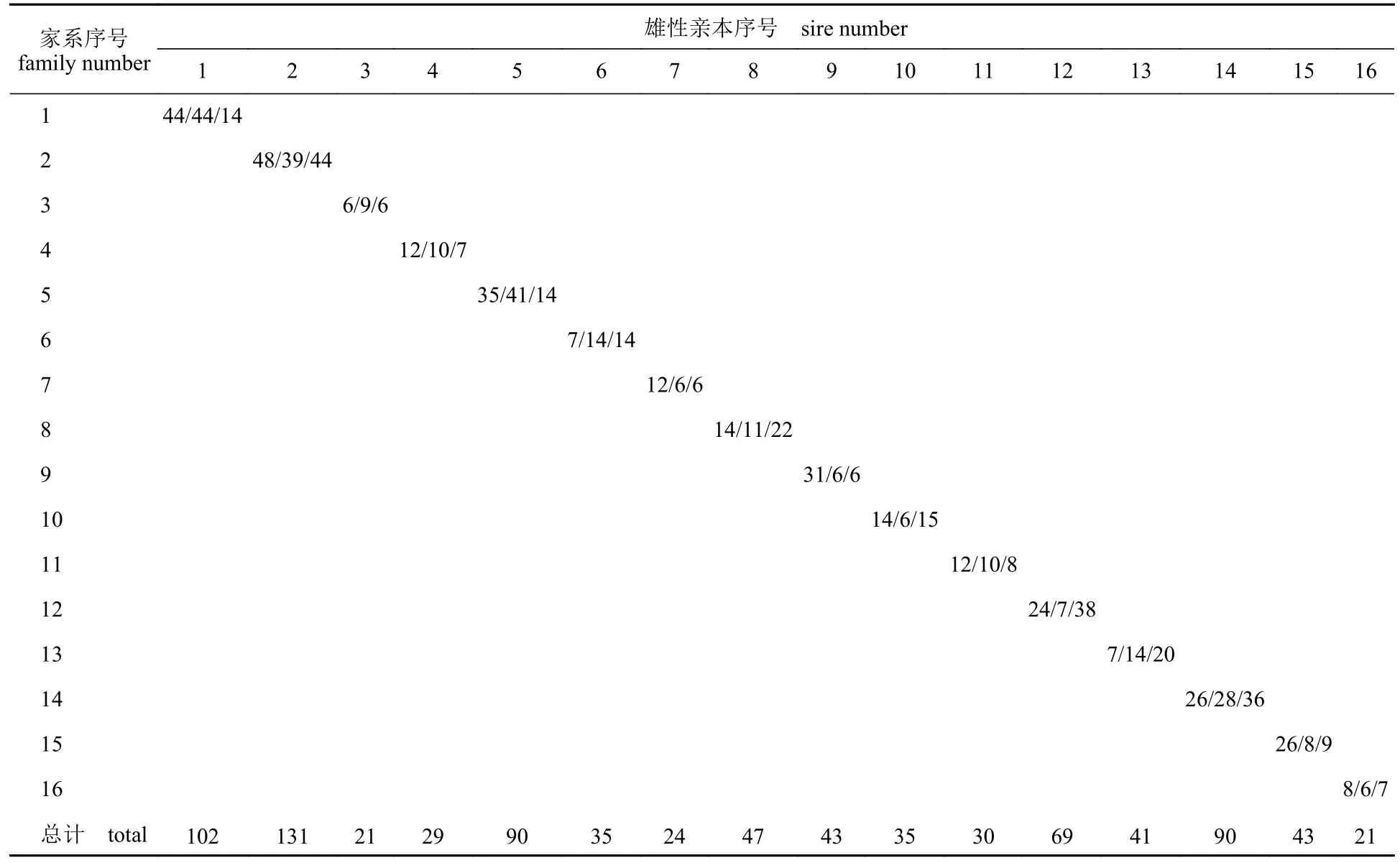
表2 基于微卫星分型的系谱鉴定结果Tab. 2 Number of progenies assigned to each of the 48 full-sib families based on microsatellite genotyping
家系重建后,统计了48个家系的壳色相关指标L*、a*、b*和ΔE。由12号母本和4号父本繁育的全同胞家系b*值的平均数最大为−85.93,而由47号母本和16号父本产生的后代的ΔE值平均数最小,为4.21 (图1)。描述性统计结果包括长牡蛎壳色参数和生长性状的平均值、标准差和变异系数(表3)。在生长性状参数中,总重量的变异系数最高,为39.62%,相比之下,壳高、壳长和壳宽的变异系数较低,为16.51%~21.10%,说明个体间总重量的差异更为明显。在4个颜色参数中,L*和ΔE的变异系数较大,说明子代个体间颜色参数中L*和ΔE的差异更为明显,L*的差异可能是个体间ΔE差异的来源;a*和b*的变异系数较小,为3.70%~5.70%。
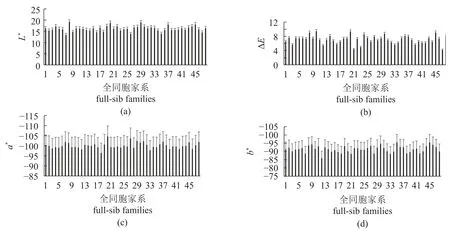
图1 长牡蛎壳色性状表型值(a)~(d)依次为L*、ΔE、a*和b*的参数值;每个全同胞家系的表型值用平均值±标准差表示,并用其母本编号命名。Fig. 1 Shell color-related trait performance in terms of L*, a*, b*, and ΔE of C. gigasParameter values of L*, ΔE, a* and b* are showed in figures (a)-(d), respectively; the shell color-related trait performances are represented by mean±SD.Each family is coded by its dam number.
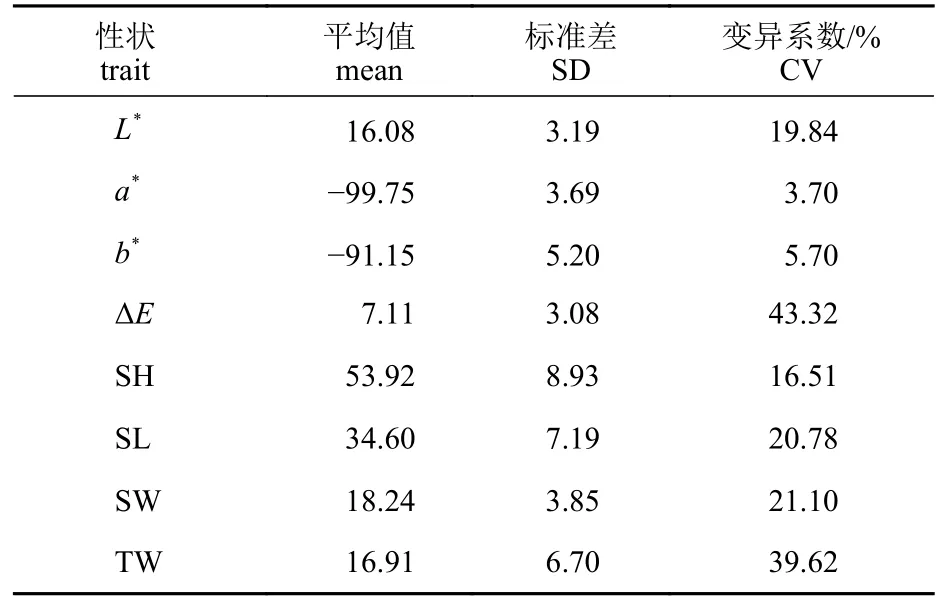
表3 长牡蛎壳色性状和生长性状参数的描述性统计结果Tab. 3 Descriptive statistics of the orange shell color-related parameters (L*, a*, b* and ΔE) and the growth-related traits of C. gigas
2.2 长牡蛎壳橙性状遗传力和遗传相关
基于家系重建后的系谱关系,利用动物模型和限制性最大似然法进行遗传力的估算。在4个颜色参数中,L*的遗传力估计值最低,为0.17±0.07。a*、b*和ΔE的遗传力估计值较高,依次为0.47±0.23、0.42±0.21和0.56±0.29 (表4)。壳色性状颜色参数内部之间相关性的差异既体现在相关性的正负上,也体现在相关性的大小上,L*和ΔE的遗传相关和表型相关为正相关,遗传相关和表型相关依次为0.51±0.30和0.08±0.04,表型相关值远小于遗传相关值;b*与L*、a*、ΔE均具有较大的负遗传相关,分别为−0.79±0.24、−0.64±0.31和−0.68±0.52。
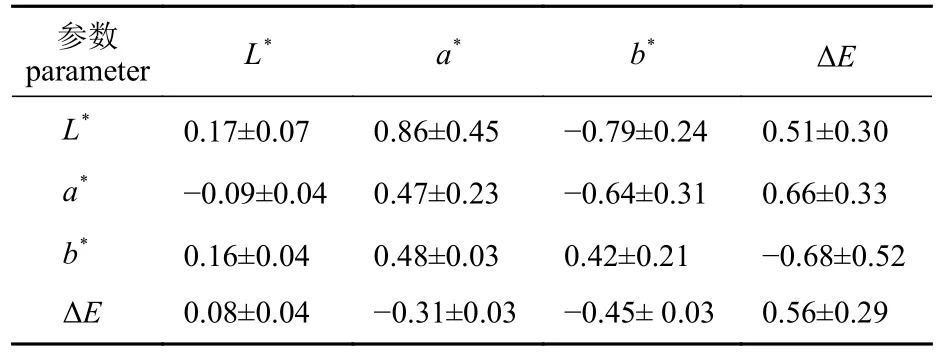
表4 长牡蛎壳色参数L*、a*、b*和ΔE的遗传力(对角线),遗传相关(对角线上方)和表型相关(对角线下方)Tab. 4 Heritabilies (in the diagonal), genetic correlations(above the diagonal) and phenotypic correlation (below the diagonal) of shell color parameters of C. gigas
2.3 长牡蛎壳橙性状与生长性状的遗传相关和表型相关
长牡蛎壳色性状和生长性状的表型和遗传相关参数列于表5。ΔE和生长性状的遗传相关和表型相关均为负值。其中,与壳高、壳长的负遗传相关程度高于其他颜色参数,为−0.29~−0.23。在L*和生长性状、a*和生长性状的遗传相关中,除L*和总重量的相关性以外,其余均具较低的相关性。在遗传相关方面,L*和总重量的负相关性最高为−0.33±0.25。在表型相关方面,a*与壳宽的相关估计值最高,为0.11±0.04;b*与总重量的相关估计值最低,为0.01±0.03。
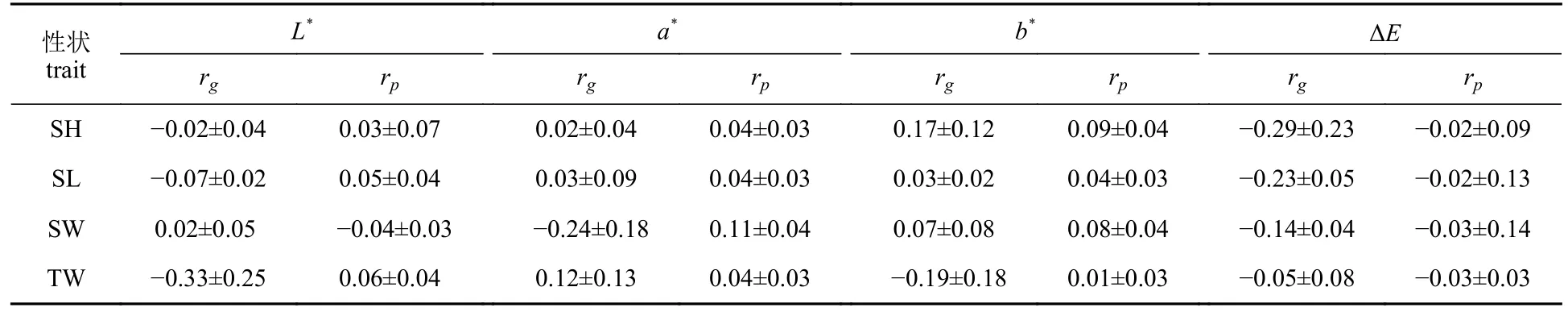
表5 长牡蛎壳橙快速生长品系壳色与生长性状的表型(rp)和遗传相关(rg)Tab. 5 Phenotypic and genetic correlation between the shell color and growth traits of the improved orange shell line C. gigas
3 讨论
3.1 长牡蛎系谱鉴定结果分析
遗传力作为水产动物选择育种中的重要遗传参数,在育种值估计、选择方法的比较、育种计划的制定等方面具有重要作用,而使用正确的系谱信息是准确评估遗传参数的前提。基于微卫星分型技术的家系重建方法,凭借其精确的鉴别力、检测的高稳定性等优点,已广泛应用到了尖吻鲈(Lates calcarifer)、鲢(Hypophthalmichthys molitrix)、鲤(Cyprinus carpio)等鱼类[21-23],以及长牡蛎[16]、耳鲍(Haliotis asinine)[24]等贝类混养家系的系谱关系鉴定中。这种将分属于不同全同胞和半同胞家系的个体养殖在共同环境中的方式,不仅降低了育种和管理成本,还排除了传统的家系分养过程引入的环境组分干扰,提升了遗传参数估测的准确性[25]。本研究运用12对微卫星位点鉴定了长牡蛎壳橙快速生长品系G1的子代,收获的863个个体中有851个鉴定到其父母本,成功率达98.61%,高于时嘉赓等[26]在皱纹盘鲍(H. discus hannai)中使用8个微卫星位点和邢德等[13]在长牡蛎中使用11个微卫星位点所得的亲子鉴定成功率,表明本研究选用的微卫星位点,其数量和多态性信息含量可以实现对大样本的家系重建。此外,在实验设计上,巢式交配设计严格控制了每一个雄性个体只能与特定的3个雌性交配,这也是提高家系鉴定成功率的原因。值得注意的是,根据家系重建的结果可以发现,混养群体中不同家系间的亲本贡献率差异较大,可能会导致选育群体有效种群大小的下降,进而产生近交。因此,在选育过程中利用分子标记监测群体的系谱信息及遗传变化,对于选育群体的种质管理有重要意义。
3.2 长牡蛎壳橙性状遗传力分析
随着长牡蛎活体、半壳等销售状态在消费市场的兴起,长牡蛎外壳的颜色、形状等壳性状开始影响消费者的消费偏好[27]。培育长牡蛎壳橙快速生长品系,不但可以满足市场多样化的需求,还可以为长牡蛎壳色性状遗传模式的研究提供材料。目前,关于长牡蛎橙壳色性状的遗传模式及相关QTL定位等研究已取得了一系列进展[28-29]。本研究通过结合使用系谱鉴定技术和计算机视觉颜色量化系统估算了10月龄长牡蛎橙壳色性状的遗传力,颜色参数L*、a*、b*、ΔE的遗传力分别为0.17、0.47、0.42和0.56。在以前的研究中,研究者将遗传力(h²)<0.15划分为低遗传力水平,0.15<h²<0.30划分为中等遗传力,h²>0.30划分为高遗传力[30]。可见,与壳色明暗程度相关的L*为低遗传力,该颜色参数可能受海区环境因素的影响较大。a*、b*、ΔE等颜色参数均为高遗传力,这表明长牡蛎橙壳色性状受遗传因素控制程度较高,以橙壳色为目标性状的选育具有较大的潜力。此外,通过家系混养的方式排除了环境效应,将非遗传方差从表型总方差去除,也可能会导致加性遗传方差增高。Xu等[14]和Evans等[4]对长牡蛎壳色遗传参数的评估中也得到了较高的遗传力。在其他贝类中,王照旗等[31]评估的三角帆蚌紫色选育系1龄阶段内壳色颜色参数L*、a*、b*、ΔE的遗传力也是中高等水平,均证明了以壳色为选育目标可以取得较好的选育效果。
3.3 长牡蛎壳橙性状与生长性状的关联性分析
选择育种是指通过挑选表型性状优良的个体繁育子代,并进行多代反复地选择与淘汰,从而选育出新品系或品种的方法。在水产动物选育过程中,需要量化表型性状以筛选最佳亲本,而有些性状的量化过程会致使个体机械损伤,导致其丧失作为亲本的有利条件,倘若容易测量的性状与不易测量的性状间具相关性,则可对后者进行间接选育。因此,在连续多代选育的过程中,需要开展目标性状关联分析以提高育种效率。表型相关主要分为遗传相关和环境相关两部分,其中,遗传相关是由基因间连锁不平衡性和基因多效性形成的[9]。通常,性状间的正遗传相关值与间接选育的效果成正比。
在海产经济贝类中,已有研究报道壳色性状和生长性状的相关程度因壳色而异。例如,邢德等[13]发现长牡蛎壳黑品系L*与生长参数遗传相关值为0.49~0.91,两性状间可进行互相选择。而Wan等[16]估测壳金长牡蛎颜色参数与各生长性状之间的遗传相关范围为−0.04~0.26,表型相关范围为−0.10~0.13,无法利用生长性状实现对壳金性状的间接选育,在三角帆蚌内壳色与生长性状的关联性分析中也得到了相同的结果[31]。本研究中,长牡蛎壳橙快速生长品系壳色性状与生长性状之间的遗传相关和表型相关普遍较低,范围介于−0.33~0.17,不同研究中长牡蛎壳色和生长性状的遗传相关存在较大的差异可能是生长环境和生长时期的不同所导致。不同养殖海区温度、盐度、海流和饵料组成不同,生物间资源分配存在差异。在不同的生长时期,母本效应大小对遗传参数的评估结果也有影响[32]。此外,不同壳色选育群体的壳色遗传模式不同,这也可能是引起不同壳色品系颜色性状的关联分析出现差异的原因。橙壳色性状与生长性状间的关联性结果说明,在长牡蛎壳橙快速生长品系的育种实践中,可通过将壳色性状和生长性状作为目标性状协同选育的方法,取得遗传进展,提高育种效率。
4 结论
本研究分析了10月龄长牡蛎壳橙快速生长品系壳色性状的表型变异、遗传力及与生长性状的关联性,进一步确定了在共同的养殖环境中以分子标记系谱鉴定为基础的家系重建方法的可行性。长牡蛎壳色性状颜色参数多具中高遗传力,以其作为选育的目标性状,有望达到良好的改良效果。长牡蛎壳色性状与生长性状没有明显的相关性,无法单一地通过壳色性状的选育来实现对生长性状的改良,将这两种性状视为目标性状协同选育是可行的措施。本研究结果为长牡蛎壳橙快速生长品系的选育提供了重要参考。
(作者声明本文无实际或潜在的利益冲突)
