硼对黑木相思幼苗生长发育的影响
2024-01-20陈朝黎白晓刚李湘阳曾炳山
陈朝黎,白晓刚,2,李湘阳,曾炳山,胡 冰*
(1.中国林业科学研究院热带林业研究所,热带林业研究国家林业和草原局重点实验室,广东 广州 510520;2.仲恺农业工程学院农业与生物学院,广东 广州 510225)
硼是植物必需的微量矿质元素,在植物的生长发育和生理代谢过程中发挥重要作用[1-2]。但植物对硼元素的适应范围较窄,土壤中有效硼含量过低或过高均会影响植物正常的生长发育[3],可能导致叶尖或叶缘失绿、黄化、焦枯[4],根系生长受阻、形态改变,植株矮小、死亡等症状。研究发现,硼胁迫抑制了桑树(Morus albaL.) 和柑橘(CitrusL.)的株高和根系的生长,降低了叶片的叶绿素含量[5-6]。并且,硼胁迫会诱导植物组织产生氧化应激损伤,导致脂膜过氧化和活性氧平衡系统改变,在6 mmol·L-1硼酸条件下,苹果砧木EM9(Malus domesticaBorkh) 的叶和茎中脂氧合酶(lipoxygenase,LOX) 活性增强, 丙二醛(malondialdehyde,MDA) 含量增加,脯氨酸(proline,Pro)含量减少[7]。氧化应激产物的积累导致氧化还原稳态受损,从而激活植物体内抗氧化系统中保护酶的活性和抗氧化剂含量的变化,硼胁迫导致植物中超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)的活性均增加,并诱导抗坏血酸(ascorbic acid,AsA)含量变化[8]。当植物处于硼胁迫环境中时其体内的代谢反应也会受到影响,如可溶性蛋白的合成[9-10]和次生代谢途径相关酶活性的变化[11]。此外,硼胁迫也会影响植物对其他营养元素的吸收和积累,施加硼营养促进了棉花对氮、磷、钾的吸收,而降低了棉花中钙和镁元素的含量[12];而缺硼会降低蚕豆(Vicia fabaL.) 对磷酸盐的吸收[13];研究火炬松(Pinus taedaL.) 细胞培养物的生长时,发现硼与钙、镁之间存在着显著的相互作用[14]。
黑木相思(Acacia melanoxylonR.Br.)属于含羞草亚科(Mimosaceae) 金合欢属(AcaciaMill.),是原产于澳大利亚的多年生高大乔木,其适应性广、抗逆性强、速生、材性优良,与根瘤共生固氮,可改良土壤、提升地力、保持水土,具有较好的经济和生态价值[15]。自20 世纪90 年代初引入我国,已在广东、广西、海南、福建等华南地区广泛引种栽培。华南地区大部分土壤的有效硼含量较低,达到缺硼(0.25~0.5 mg·kg-1)或严重缺硼等级(<0.25 mg·kg-1)[16],在林木培育过程中多需施以硼肥,因此,如何避免林木缺硼或硼过量胁迫,是南方人工林培育过程中亟待解决的问题。目前,在杨树(Populus)、桉树(Eucalyptus urophyllaS.T.Blake)及针叶树中进行了一些硼营养的研究[17-20],但林木的生长发育对硼胁迫的响应研究仍不够完善。本研究以黑木相思无性系SR17 幼苗为材料,分析了不同供硼量条件下幼苗的生长特性、生理生化特征的变化,旨在明确黑木相思对不同供硼量的响应模式,为阐明植物对硼胁迫的抵御机理奠定理论基础,为林木高产、优质栽培提供科学依据。
1 材料与方法
1.1 试验材料与设计
以黑木相思无性系SR17 为研究材料,选取高度一致且长势相近的1 月龄水培幼苗,用超纯水冲洗干净根部后,定植于海绵固定的育苗盘中,盘长30 cm、宽24 cm、高10 cm,盘内添加1 L 不含硼的1/2 MS 营养液作为基础营养液,pH 为5.8,培养于中国林业科学研究院热带林业研究所育苗室 (23°20' N,113°19' E),室内日平均温度约25 ℃,平均湿度约70%,自然光照强度约6 000~7 000 Lux。以硼酸为硼源,设置0、0.1、1、2 mmol·L-1四个硼水平处理,分别添加至基础营养液中,处理2 个月,每2 天补充新鲜营养液,每周更换新鲜营养液,各处理分别记为B0、B0.1、B1、B2,每处理设4 个育苗盘为4 个重复,每重复设15 株。
1.2 生长指标的测定
测定培养0 d 和2 个月后各处理植株的株高、叶片数、根长等生长指标,每处理设4 个重复,每个重复15 株幼苗。
1.3 叶绿素含量的测定
分别称取各处理植株地上部分样品0.5 g,以80%丙酮溶液为提取液,利用分光光度计法测定645 nm、663 nm 波长下的吸光值,参照公式:叶绿素a=12.7 ×A663-2.69 ×A645;叶绿素b=22.9 ×A645-4.68 ×A663,总叶绿素含量=叶绿素a + 叶绿素b,计算样品的叶绿素含量[21],每处理从4 个育苗盘中分别随机选择1 株,4 株设为一重复,共6 个重复。
1.4 生理活性物质含量和氧化还原酶活性的测定
分别取各处理植株的地上部分为样品,利用改良型Bradford 法测定可溶性蛋白的含量[22];利用硫代巴比妥酸法测定MDA 含量[23];采用茚三酮法进行Pro 的提取与测定[24];利用抗坏血酸氧化酶催化AsA 氧化生成脱氧抗坏血酸(DHA)的原理测定AsA 的氧化速率从而测定AsA 含量[25];利用氮蓝四唑光还原法测定SOD 活性[26];采用紫外吸收法测定CAT 活性[27];利用愈创木酚法测定POD活性[28];参照Wang 等[29]的方法测定LOX 活性;采用L-苯丙氨酸法测定PAL 活性[30],每处理从4 个育苗盘中分别随机选择1 株,4 株设为一重复,共6 个重复。
1.5 营养元素含量的测定
分别取各处理植株地上部分为样品,至于烘箱中105 ℃杀青60 min,65 ℃烘干至质量恒定,进行氮、磷、钾、钙、镁、硼等营养元素含量的测定。参照NY/T2017-2011,利用全自动定氮仪测定氮的含量,利用分光光度法测定磷的含量,利用火焰原子吸收分光光度法测定钾的含量;参照DB12/T 846-2018,利用等离子发射光谱仪测定钙、镁的含量;参照LY/T1273-1999,利用干灰化-甲亚胺比色法测定硼的含量。每处理从4 个育苗盘中分别随机选择1 株,4 株设为一重复,共6 个重复。
1.6 数据处理与统计分析
利用Excel 2016 和 SPSS 22 对测定结果进行数据处理和ANOVA 单因素方差分析,利用Turkey HSD 进行事后多重比较。选择具有显著性差异的指标进行主成分分析,在分析前先对数据进行标准化处理,以消除量纲差异,处理方法采用x-ij =(xij-x-j)/sj,其中xij为各指标原始数据,xj为各处理原始数据平均值,sj为标准差。主成分分析以选择特征值大于1,累积贡献率达到90%以上为标准保留主成分。计算各主成分权重 (W)及综合得分(F), 权重计算公式:Wj=Pj/∑Pj,式中,Wj表示第j 个主成分的权重,Pj表示j 主成分的贡献率, 获得综合指标值F=∑(Fj× Wj)[31],Fj表示各主成分的得分。利用公式计算各指标数据标准化后的隶属函数值:Uj=(Xj-Xjmin)/(Xjmax-Xjmin),其中Xj为第 j 个综合指标;Xjmin 为第 j 个综合指标的最小值;Xjmax 为第 j 个综合指标的最大值。最后将隶属函数和权重结合,获得综合评价值(D),计算公式为D=∑(Uj× Wj)。
2 结果与分析
2.1 供硼量对黑木相思幼苗生长指标的影响
不同供硼量条件下,植株的生长表型有所不同(图1)。B0 的植株叶片(图1A) 比B0.1(图1B)稍黄;而B1 和B2 呈现出叶缘失绿的表型,且部分叶片黄化脱落(图1C、D)。根系方面,B0 形成了白色较粗的主根,侧根也较短而粗(图1E);而B1 和B2 侧根较细,且随着硼浓度增高而越发褐色(图1G、H)。由此说明,无硼的培养条件使黑木相思幼苗发生缺硼症状,叶片发黄,主根和侧根均加粗,且侧根变短;随着硼浓度的增加,幼苗受到了硼过量的毒害作用,叶缘失绿,老叶黄化脱落,侧根变细,根系褐化。

图1 不同供硼量对黑木相思幼苗叶和根的影响Fig.1 Effects of different boric acid content on the leave and roots of A.melanoxylon seedlings
由生长指标测定结果(图2) 得出,B0.1 植株株高增长量较B0 植株株高增长量大;但随着供硼量的继续增加,株高增加量反而减小,B2 株高增长量为7.05 cm,仅为B0.1 株高增长量的54.9%。根长生长与株高生长情况类似,B0.1 根长增长量较B0 根长增长量大;而B1 和B2 的根长增长量仅为B0.1 的59.6% 和41.7%。随着供硼量的增加,植株叶片数增加量逐渐减少。由此说明,B0.1 黑木相思幼苗生长较好;B0 形成缺硼表型,叶片发黄,侧根变短,主根和侧根加粗;B1 和B2 造成硼过量的毒害作用,叶缘失绿,根系褐化,株高、叶片数、根长生长均受到抑制。因此,供硼量是影响黑木相思生长发育的重要因素之一。
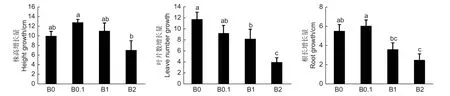
图2 不同供硼量培养后黑木相思幼苗的生长指标Fig.2 Growth indexes of A.melanoxylon seedlings cultured with different boric acid content nutrient solution
2.2 供硼量对黑木相思幼苗叶绿素含量的影响
图3 表明,B0 植株体内叶绿素a 和叶绿素b 含量均低于B0.1 植株;B0.1 植株的叶绿素a 和叶绿素b 含量最高;而B1 和B2 植株的叶绿素a 和叶绿素b 的含量逐渐降低。由此推断,0、1 和2 mmol·L-1的供硼量均引起黑木相思地上部分叶绿素含量降低,抑制叶片光合作用,从而影响植株的正常生长发育。

图3 不同供硼量培养后黑木相思幼苗的叶绿素含量Fig.3 Chlorophyll content of A.melanoxylon seedlings cultured with different boric acid content nutrient solution
2.3 供硼量对黑木相思幼苗生理活性物质含量的影响
植物体内的可溶性蛋白的含量是反映植物总代谢的重要指标,如图4 所示,随着供硼量的增加,黑木相思幼苗体内可溶性蛋白含量越高,植株体内的代谢反应越活跃。AsA 是植物细胞中保护叶绿体的重要的抗氧化剂,AsA 的含量在B0.1 植株体内最高,随着供硼量的增加,AsA 的含量急剧降低,说明硼过量条件促进了黑木相思体内的氧化反应造成叶绿体损伤。MDA 是过氧脂质分解出的产物之一,其含量可以反应植物体内脂质氧化的水平,在B0、B1 和B2 植株体内MDA 含量较B0.1 植株的高,说明硼胁迫引起黑木相思体内脂质氧化水平升高。Pro 是植物抵御逆境时的渗透调节物质之一,用于保持细胞质基质与环境的渗透平衡,防止水分散失,Pro 含量在B0、B1 和B2 植株体内较高,说明缺硼和硼过量条件造成了黑木相思的渗透胁迫。
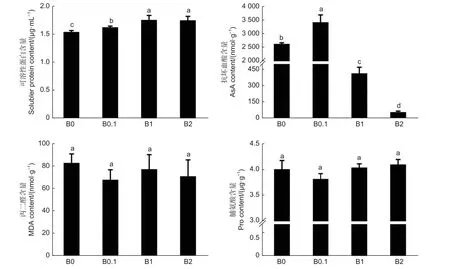
图4 不同供硼量培养后黑木相思生理活性物质的含量Fig.4 Physiologically active substance content of A.melanoxylon seedlings cultured with different boric acid content nutrient solution
2.4 供硼量对黑木相思幼苗氧化还原酶活性的影响
SOD 和POD 是植物体内氧自由基和H2O2的重要清除剂,保护植物免受氧化反应毒害。如图5 所示,在B0、B1 和B2 幼苗体内SOD 和POD 的活性较高,推断缺硼和硼过量条件使黑木相思幼苗发生了氧自由基和H2O2的积累。CAT 是植物体内清除H2O2的主要酶之一,随着供硼量增加,幼苗体内产生的H2O2越多,CAT 的活性越高。PAL 是植物体内次生物质苯丙烷类代谢的关键酶,PAL 活性在B0、B1 和B2 幼苗中升高,说明缺硼和硼过量条件促进了黑木相思体内次生代谢物的合成。LOX 是一种催化膜脂过氧化的酶,其活性越高使得植物体内的过氧化程度越大,对植株产生毒害作用增加。LOX 活性在B0、B1 和B2 幼苗体内增加,说明缺硼和硼过量条件使黑木相思幼苗产生了较剧烈的过氧化反应。以上结果表明,缺硼或硼过量条件影响了黑木相思幼苗生化水平的稳定,促进了LOX 酶活性的增加,使细胞膜脂产生过氧化反应;促进了SOD、POD、CAT 等抗氧化酶和次生代谢物合成酶PAL 的活性增加,以抵御过氧化反应的毒害作用,协调控制黑木相思的生长发育。

图5 不同供硼量培养后黑木相思幼苗氧化还原酶的活性Fig.5 Oxidoreductase activity of A.melanoxylon seedlings cultured with different boric acid content nutrient solution
2.5 供硼量对黑木相思幼苗营养元素含量的影响
在植物体内,不同营养元素之间会产生促进或拮抗的作用,这些相互作用完成植物生理生化过程,调节着植物的营养状况,影响着植物的生长发育。结果表明(图6),植株体内硼元素的含量随着培养液中供硼量的增加而增加,尤其在过量硼供应条件下(B1 和B2) 发生硼元素的大量积累,分别为B0.1 硼含量的10.94 倍和12.95 倍;随着供硼量的增加,磷和钾元素的含量逐渐增加;而氮和镁元素的含量无明显变化;钙元素在生长状态较好的植株(B0.1 ) 中的含量较高,随着硼的过量供应,钙含量逐渐降低,B1 和B2 的钙含量分别较B0.1 的降低15.38%和23.08%。由此说明,黑木相思体内硼元素与不同元素之间的相互作用有所不同,硼元素的积累可以促进磷和钾元素的积累,即硼元素与磷和钾元素具有协同作用;硼元素与氮和镁元素无明显相互作用;在硼元素促进植株生长的条件下,可以促进钙元素的积累,但硼过量抑制植株生长时,也抑制了钙元素的积累。
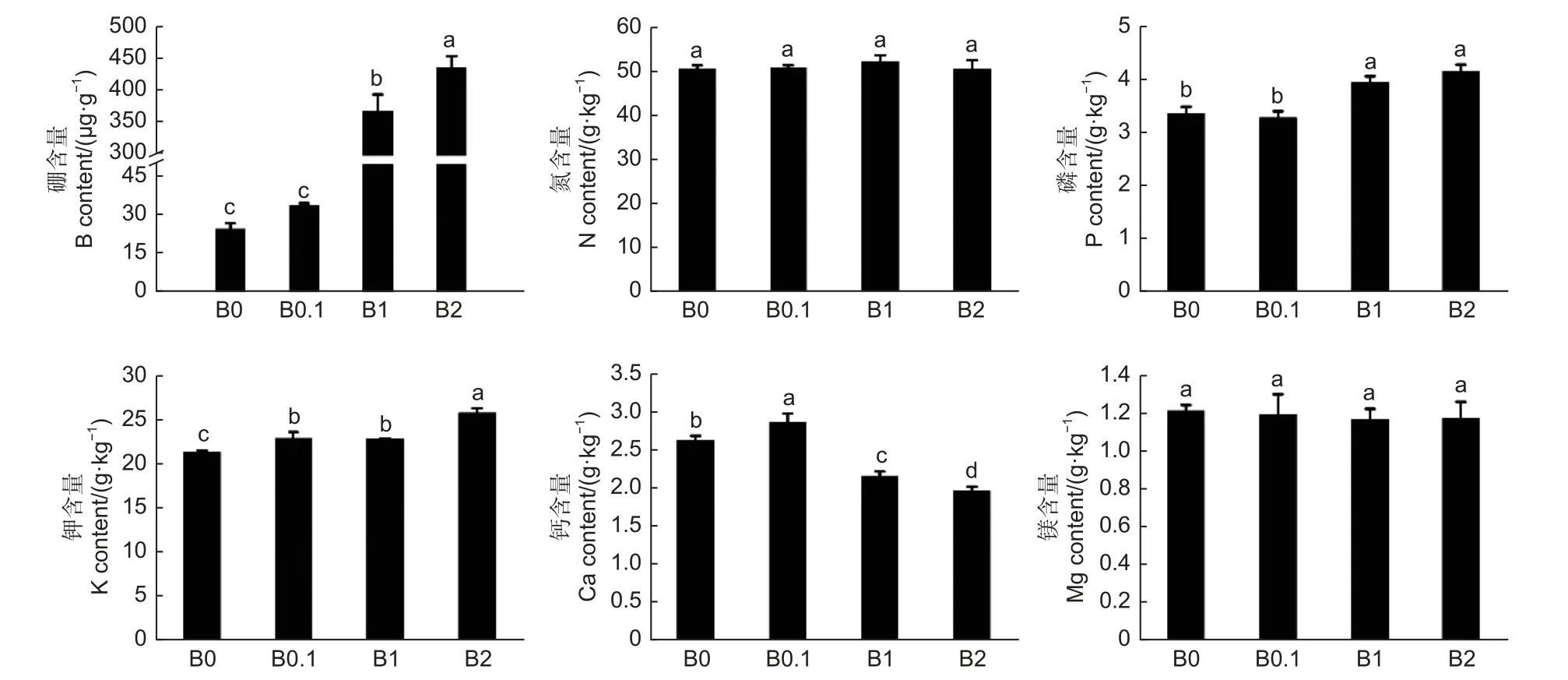
图6 不同供硼量培养后黑木相思幼苗营养元素的含量Fig.6 Mineral element content of A.melanoxylon seedlings cultured with different boric acid content nutrient solution
2.6 主成分分析与隶属函数综合分析
为了更明确地分析和评价各指标对不同供硼量的响应程度,将具有显著性差异的17 个指标进行主成分分析。结果得出(表1),前两个综合指标的贡献率分别为75.14%、19.66%,累计贡献率达94.80%,表明提取前两个主成分可以代表大部分指标的信息。以特征向量系数作为评价标准(表2),第一主成分特征值为12.77,其中钾元素含量、叶片数增长量、株高增长量、LOX 活性、AsA 含量、钙元素含量、硼元素含量、POD 活性、Pro含量、SOD 活性、根长增长量、蛋白浓度、PAL活性的荷载量较高;第二主成分特征值为3.34,其中叶绿素b 含量的荷载量较高。因此,可取以上14 个指标作为评价黑木相思对不同供硼量条件适应性的主要指标。
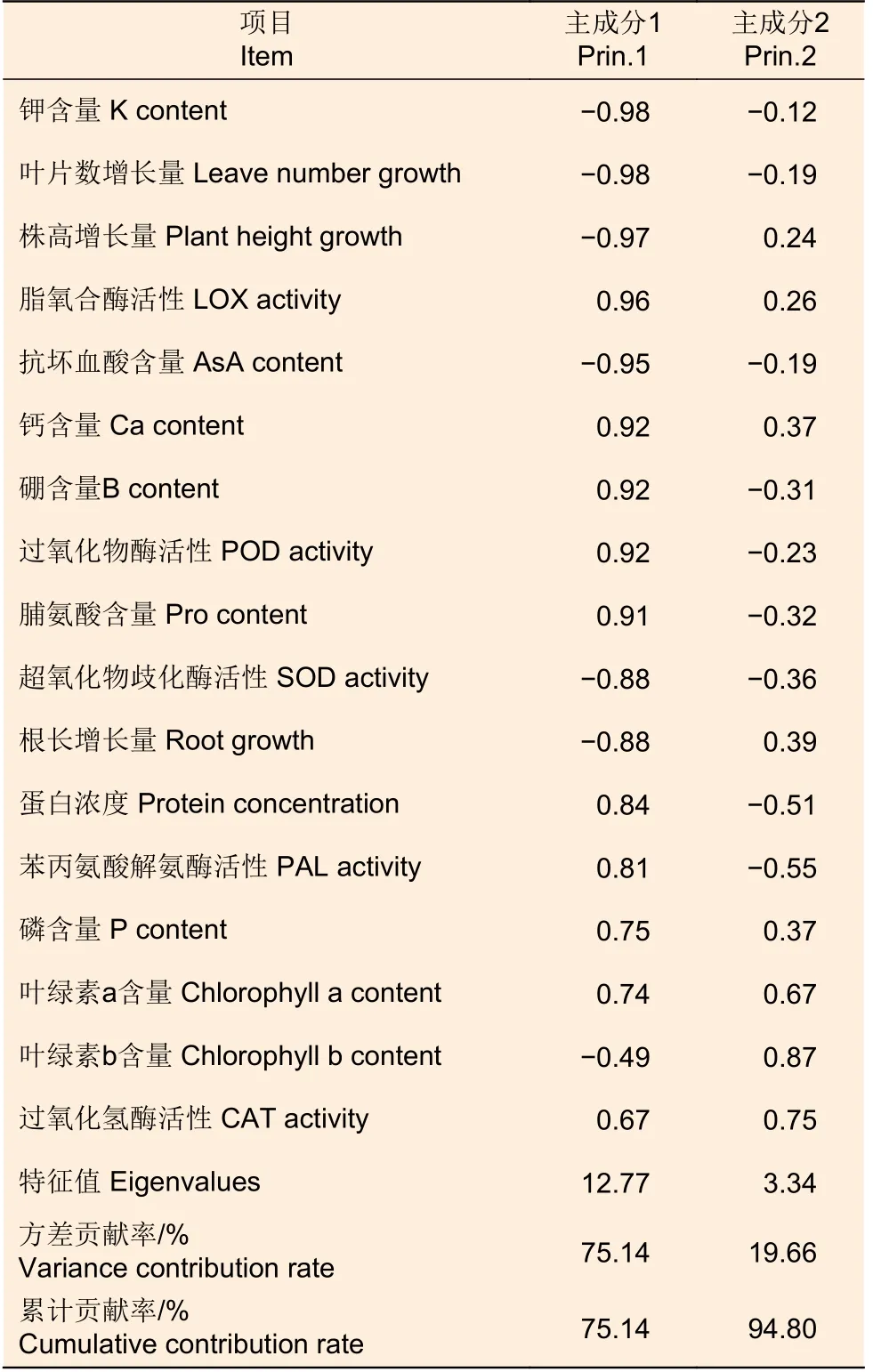
表1 综合评价主成分指标系数及贡献率Table 1 Principal components eigenvectors and cumulative contribution rates
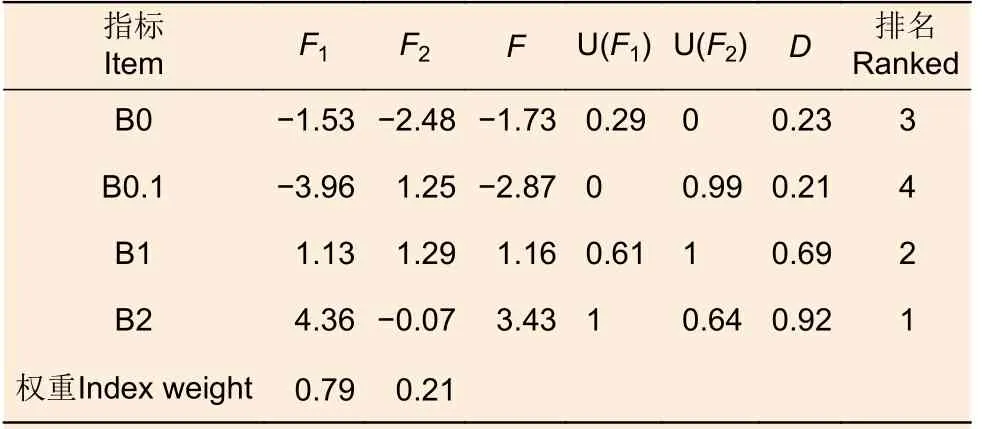
表2 不同供硼量条件下各指标的主成分和隶属函数分析及综合评价Table 2 Comparison and comprehensive evaluation of the subordinate function values under different boric acid supply
根据主成分特征向量系数及贡献率(表2),获得前两个主成分的因子方程:
F1=-0.27Q1-0.27Q2-0.27Q3+0.27Q4-0.27Q5+0.26Q6+0.26Q7+0.26Q8+0.25Q9-0.25Q10-0.25Q11+0.24Q12+0.23Q13+0.21Q14+0.21Q15-0.14Q16+0.19Q17;
F2=-0.06Q1-0.10Q2+0.13Q3+0.14Q4-0.10Q5+0.20Q6-0.16Q7-0.12Q8-0.17Q9-0.19Q10+0.21Q11-0.27Q12-0.29Q13+0.19Q14+0.35Q15+0.46Q16+0.39Q17(式中Qi 为各指标标准化后的值)。
主成分综合得分公式为F=0.79F1+ 0.21F2,分别计算不同供硼量条件下,前两个主成分的综合得分(F),得出黑木相思的生长发育受胁迫程度排序为B2>B1 >B0 >B0.1。另外,利用权重(W)及隶属函数值求得不同供硼量条件下黑木相思幼苗生长发育的综合评价值(D),结果表明(表2),对黑木相思幼苗生长发育的胁迫程度同为B2>B1 >B0>B0.1。因此,黑木相思幼苗相对适宜的供硼量为0.1 mmol·L-1,而 0、1 和2 mmol·L-1的供硼量均影响黑木相思幼苗的生长发育。
3 讨论
充足的硼营养对植物生长发育至关重要,研究黑木相思受硼胁迫时生长特性及生理生化特征的变化,对解析林木对硼胁迫的抵御机理,定向培育优良品种,促进林木生产力具有重要意义。本研究发现,缺硼和硼过量条件均抑制了黑木相思幼苗的生长,导致植株矮小,根系变短加粗。以往的研究也证实了硼胁迫会导致植物生长减缓,如缺硼胁迫导致枳橙(Poncirus trifoliate(L.) Raf.)节间缩短,根系明显变粗和褐化,且根尖膨大[32];硼过量时葡萄(Vitis viniferaL.)的叶片和根系生长均受到显著抑制[33];麻风树(Jatropha curcasL.)植株整体生长降低,且根系的生长减少程度高于叶片和茎部[34]。供硼量对黑木相思的叶片数增长量有显著影响,这可能是因为硼元素在维持细胞壁结构完整性中起着至关重要的作用[35]。缺硼条件下,植株顶端分生组织细胞壁完整性受损[36-37],硼过量的环境又造成细胞壁过度增厚[5],细胞无法完成正常的分化进程,引起器官的结构发育不良,进而导致植株顶端分生组织生长异常,叶片数增长受到影响。另外,硼胁迫还导致黑木相思幼苗叶片失绿黄化,叶绿素含量显著降低。这是因为硼胁迫下细胞膜透性增加,叶绿素初始前体氨基乙酰丙酸积累受到影响[38],造成叶绿素含量降低,叶片黄化。Hegazi 等[39]对橄榄(Olea europaeaL.) 喷施叶面硼酸也证明了叶绿素a、叶绿素b、总叶绿素含量与硼浓度显著相关。因此,硼胁迫会破坏植株的细胞结构,抑制植株株高和根系生长,且降低叶绿素含量,抑制光合作用,从而影响植株的正常生长。
植物遭遇硼胁迫通常会导致体内活性氧积累,从而诱导细胞膜的氧化损伤。本研究发现,0、1 和2 mmol·L-1供硼量下,黑木相思地上部LOX活性和MDA 含量均高于0.1 mmol·L-1供硼量下的,这表明缺硼和硼过量环境均对植株造成了胁迫,导致植株的细胞膜透性增加,脂质过氧化反应增强。同时,本研究中AsA 的含量在0、1 和2 mmol·L-1供硼量下均低于0.1 mmol·L-1供硼量下的,表明缺硼和硼过量环境中过量的活性氧消耗了植物体内的AsA,导致AsA 含量降低。SOD、CAT 和POD 均为重要的抗氧化酶,本研究中SOD和POD 活性在硼胁迫条件下显著升高;CAT 活性在缺硼条件下降低,硼过量条件下升高。这表明硼胁迫条件诱发了植物细胞的氧化应激保护机制,而不同的抗氧化酶在面对胁迫反应时的敏感程度不尽相同,活性氧的清除系统由多种抗氧化酶共同对抗氧自由基组成,以抵御对植株产生伤害。
植物体内的硼元素含量可以通过复杂的作用网络影响植物对其他营养元素的吸收和运输。本研究发现,随着营养液中硼酸浓度的增加,黑木相思地上部硼含量也随之增加,表明植株对硼含量的积累与环境中硼酸浓度有关,过多的硼酸供应将导致植株体内的硼元素含量累积超过植株自身需要,从而造成硼胁迫。磷是核酸、磷脂、核苷酸的关键成分,在代谢中发挥重要作用[40];钾是40 多种酶的辅助因子,是形成细胞膨胀和维持细胞内电中性的主要阳离子[41]。本研究发现,随着黑木相思地上部分硼含量的积累,磷和钾元素的吸收也相应增加。前人关于硼和磷[42-43]、硼和钾[44]的联合施用对作物生长发育影响已进行大量研究。早在1974年,Robertson 和Loughman[13]就已经证明,缺硼会阻碍磷的吸收,而充足的硼供应会增加植物对磷的吸收。这可能是因为硼含量对植物体内质膜的特性及质膜结合质子泵核苷酸(ATP) 酶的活性产生影响,直接或间接地调节质子运输,缺硼降低了植物对磷和钾的吸收和运输,而硼供应增强了膜结合ATP 酶的活性,可以促进磷和钾元素的积累[45]。过去普遍认为硼钙间存在拮抗作用[46],但随着研究的深入,研究者发现硼钙之间存在着错综复杂的关系[47],钙和硼功能类似,可以结合在果胶的不同位点上,二者共同作用维持细胞壁结构的稳定性。本研究中,在0、1 和2 mmol·L-1供硼量下,黑木相思地上部分钙元素含量均显著低于0.1 mmol·L-1供硼量下的。缺硼环境下植株地上部分钙含量较低,可能是因为钙是不易移动的元素,植株吸收的钙被根系固定,较少被地上部生长利用;而在硼浓度较高环境下钙含量较低,可能是因为过量的硼供应导致钙在细胞壁中的结合位点减少,因此钙吸收量减少。不同硼胁迫环境下,不同物种体内各元素的积累模式会有所不同,如在硼过量情况下,麻风树叶片中钾、镁的浓度增加,而磷浓度略有下降[34],与我们的研究结果有所不同。因此,不同供硼量条件下,植物对氮、磷、钾、钙、镁等营养元素的吸收及平衡机制,有待深入探讨。
主成分分析作为基础的数学分析方法,应用十分广泛。在具有多个变量时,变量之间的相关性会对分析造成一定的复杂性。本研究将17 个具显著性差异的指标结果经过主成分分析,转化成两个不相关变量(累计贡献率为94.80%),以减弱指标之间的相关性带来的影响。根据特征向量系数得出,钾元素含量、叶片数增长量、株高增长量、LOX 活性、AsA 含量、钙元素含量、硼元素含量、POD 活性、Pro 含量、SOD 活性、根长增长量、蛋白浓度、PAL 活性及叶绿素b 含量的荷载量较高。因此,可将这14 个指标作为硼胁迫影响黑木相思生长发育的主要指标,为进一步研究黑木相思抵御硼胁迫的机理提供理论参考。另外,通过主成分综合得分和隶属函数值均得出,黑木相思的生长发育受胁迫程度排序为供硼量B2>B1>B0>B0.1,最终判定0.1 mmol·L-1供硼量时黑木相思幼苗生长发育表现最好。
4 结论
总之,本研究表明硼胁迫抑制了黑木相思茎的伸长和根系的发育,使得叶片失绿、黄化甚至脱落;降低了黑木相思地上部分叶绿素的含量,从而抑制了光合作用;降低了黑木相思体内AsA 的含量,引起了氧化反应,增加了MDA 和Pro 含量,引起体内脂质氧化水平升高和渗透胁迫;受硼胁迫后,黑木相思体内SOD、POD、CAT、LOX 和PAL 的活性较高,引发了氧自由基和H2O2的积累与清除以及次生代谢物的合成。另外,黑木相思体内硼元素的积累可以促进磷和钾元素的积累;在硼元素促进植株生长的条件下,也可以促进钙元素的积累。综合分析表明,钾元素含量、叶片数增长量、株高增长量、LOX 活性、AsA 含量、钙元素含量、硼元素含量、POD 活性、Pro 含量、SOD活性、根长增长量、蛋白浓度、PAL 活性及叶绿素b 含量这14 个指标可作为硼胁迫影响黑木相思生长发育的主要指标。本研究揭示了黑木相思对硼胁迫环境的响应模式,为黑木相思硼营养诊断和栽培管理提供了科学依据,同时也为阐明林木对硼胁迫的抵御机理奠定了理论基础。
