黄杨斑蛾田猎姬蜂(膜翅目:姬蜂科)个体发育与生殖行为
2024-01-20田良恒张锐敏尹新明白素芬
王 于,田良恒,李 欣,张锐敏,尹新明,白素芬
(河南农业大学植物保护学院,河南 郑州 450002)
田猎姬蜂属(Agrothereutes)昆虫是林木害虫的重要寄生性天敌资源,迄今为止,在我国仅报道9 种,分别是短翅田猎姬蜂(A.abbreviatus(Fabricius))、枝田猎姬蜂(A.ramuli(Uchida))、大田猎姬蜂(A.macroincubitor(Uchida))、基田猎姬蜂(A.ruficoxalisSheng & Sun)、赣田猎姬蜂(A.ganicusSheng & Sun)、食心虫田猎姬蜂(A.grapholithae(Uchida))、三色田猎姬蜂(A.tricolor(Uchida))、叶蜂田猎姬蜂(A.aproceriusLi & Sheng) 和黄杨斑蛾田猎姬蜂(A.minousubaeNakanishi)[1~4],分布仅限于辽宁、吉林、江西、山东、浙江等地。国内有关田猎姬蜂的研究主要集中在新种或新记录种的记述[3-4]。此外,尚无相关深入的研究和报道。黄杨斑蛾田猎姬蜂 (Agrothereutes minousubaeNakanishi)(Hymenoptera: Ichneumonidae)是近年来发生频繁、危害严重的园林主要害虫大叶黄杨斑蛾(Prgeria sinicaMoore)预蛹和蛹期的重要外寄生蜂。在日本,该蜂是大叶黄杨斑蛾的专性寄生蜂,寄生率高达70%,是引起寄主死亡,控制其种群增长的关键因子[5]。
2019 年4 月,在郑州市河南农业大学校园内,受大叶黄杨斑蛾危害的大叶黄杨周围发现大量正在取食和飞舞的姬蜂,经采集并参照《浙江蜂类志》进行鉴定[6],疑为黄杨斑蛾田猎姬蜂。随后又与日本学者发表此新种时的形态特征进行比对[7],确定为该种。这是继日本、中国杭州分布外的新增分布地,为河南新记录种,河南新记录属。国内对黄杨斑蛾田猎姬蜂的研究仅限于形态特征的描述[6]。日本学者在发现此种的基础上,进一步开展黄杨斑蛾田猎姬蜂与寄主大叶黄杨斑蛾的年生活史及光周期和温度对该蜂滞育的影响[1,8]。然而,国内外有关该蜂的个体发育特征,特别是生殖行为方面研究仍属空白。园林害虫大叶黄杨斑蛾原分布于中国、日本、朝鲜和韩国,现已入侵至美国[9],在我国发生的普遍性和严重性加剧[10~12]。当前,国内对大叶黄杨斑蛾主要以化学药剂防治为主[13]。为实现园林植物的绿色保护,寻求安全有效的生物防治途径控制大叶黄杨斑蛾的危害势在必行。为此,开展基于个体发育特征的研究有助于对黄杨斑蛾田猎姬蜂的识别和保护,而生殖行为的明确和理解将有助于其生防作用的有效发挥[14-15]。
1 材料与方法
1.1 供试昆虫
自2019 年4 月至2021 年5 月,实地调查河南农业大学校园内大叶黄杨斑蛾及黄杨斑蛾田猎姬蜂发生情况。黄杨斑蛾田猎姬蜂成虫采自被大叶黄杨斑蛾危害的大叶黄杨周围,采集时间为2019 年4 月中旬至5 月上旬和2020 年6—7 月。自2020年6 月起,每周在上述地点进行大叶黄杨斑蛾茧采样(大叶黄杨树下落叶层),剥茧所得的黄杨斑蛾田猎姬蜂幼虫在实验室24 ± 1 ℃, RH 55%~65%,14 L∶10 D 条件下饲养。
1.2 田猎姬蜂个体发育过程观察
将采集的黄杨斑蛾田猎姬蜂成虫,以及经室内培养的老熟幼虫化蛹羽化得到的成蜂,以10%蜂蜜水饲养。交配后,每日提供10~20 头大叶黄杨斑蛾茧供每头雌蜂寄生,随后将被寄生的寄主放置在上述条件下培养。初产卵至幼虫孵化期间,每间隔12 h 在显微镜(LeicaM205A)下观察卵形态及颜色变化并拍照。待幼虫孵化后,选取10 头初孵幼虫,每天特定时间在体视显微镜(OLYMPUS-SZX7)下观察其形态变化并用相机(CanonPowershotA 650IS)拍照,记录蜂幼虫的蜕皮数,以确定幼虫的虫龄。老熟幼虫初化蛹时将其在寄主茧内所结的一层薄茧撕开,置于玻璃培养皿内,选取5 头在体视显微镜下观察其蛹期发育及色素沉积过程,直至羽化。用相机拍照胚后发育的形态。
1.3 成虫生殖系统解剖
选取上述条件饲养出的雌蜂(已交配未产卵)和雄蜂各10 头,先经-20 ℃短暂冻存后取出,在体视显微镜下用解剖镊夹住雌蜂腹部末端,轻轻拉
出生殖系统放入灭菌后的PBS 溶液中,在LEICA倒置相差显微镜及LeicaM205A 显微镜下拍照,并观察雌蜂卵巢管类型和统计卵巢管数量;雄蜂同上述处理,拍照内生殖器官和交配器结构。
1.4 生殖行为观察
1.4.1 求偶和交配 选取羽化当天或羽化1~2 d雌、雄蜂按1∶1 配对,共10 组,室温条件观察求偶和交配行为。交配装置为H=15 cm,R下底=7 cm,R上底=4.5 cm 的透明塑料盒。
1.4.2 产卵寄生 室温条件下,选10 头交配后1~2 d 的雌蜂,此时蜂体内孕育充足的成熟卵子。将蜂分别放入上述规格容器内(含5 头寄主茧和5 头寄主蛹),共10 组。在9:00—13:00时,使用摄像机(CanonPowershotA 650IS)观察记录雌蜂寄生行为。待观察结束后,将容器内寄主茧和蛹取出,并镜检是否被寄生。用含有10%蜂蜜水的棉花球饲喂雌蜂,放置于15 ± 1 ℃,RH50%~60%,14 L∶10 D 恒温培养箱(ZRG-250A-L,上海丙林电子科技有限公司)内饲养。次日重复相同的过程并持续观察记录1 周,随后统计子代蜂的性比。
2 结果与分析
2.1 黄杨斑蛾田猎姬蜂个体发育特征
2.1.1 胚胎发育 卵呈长椭圆形,略呈梭状,一端略大一端稍小,初产卵为乳白色,长径1.57 ±0.07 mm,短径0.35 ± 0.02 mm(图1A)。相差镜下,经强光照射,卵晶莹剔透,卵内物质分布均匀(图1B);12 h 后渐变为微淡黄色,卵内物质向中间聚集(图1C),24 h 后呈乳黄色(图1D),48 h 后卵壳内黄色团块状胚胎发育完成,卵较宽端即头部位置可见口器的褐色上颚(图1E),胚胎可上下轻微活动。产卵约50 h 后,幼虫孵化,从卵较宽端破壳而出(图1F)。从产卵到幼虫孵化,胚胎发育历经48~50 h。
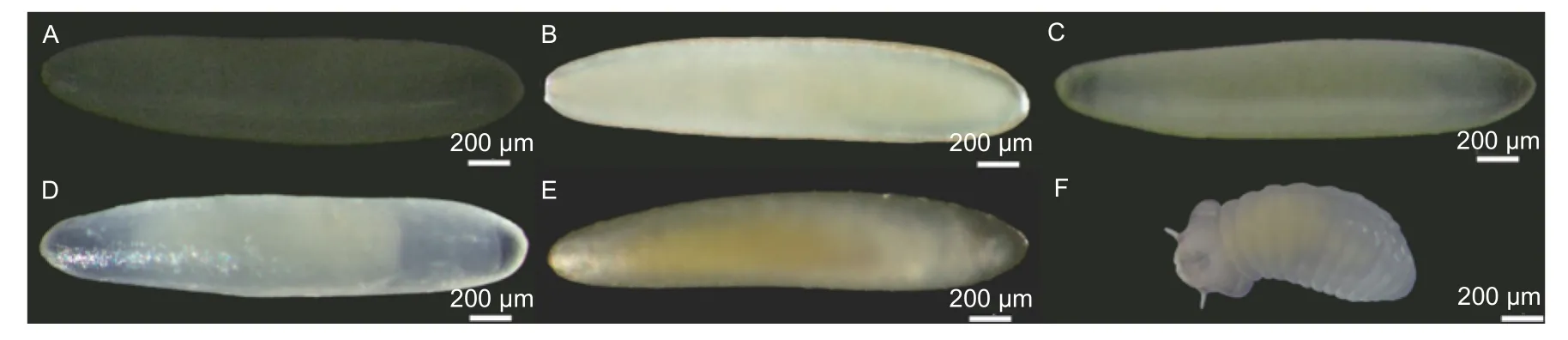
图1 卵的形态与初孵幼虫Fig.1 Morphological characters of egg and newly hatched larva
2.1.2 胚后发育 伴随取食,幼虫经历3 次蜕皮,共4 个虫龄。初孵幼虫头部具有骨化程度不强的头壳,触角较长,褐色的上颚发达。除头节外,体分13 节,其中胸部3 节,腹部10 节。头部明显宽于胸部和腹部,腹部末端细长,体壁薄而透明,消化道呈淡黄色。初孵幼虫体表无脂肪粒。取食4 h 后,1 龄幼虫头壳宽为0.37 ± 0.02 mm,体长1.35 ± 0.07 mm,体宽0.43 ± 0.01 mm,腹部靠后两侧出现有脂肪体颗粒(图2A)。孵化24 h后,消化道已变成棕红色,脂肪体颗粒显著增多。2 龄幼虫体呈纺锤形,前后两端小,而中部膨大。头壳宽为0.50 ± 0.01 mm,体长2.57 ± 0.02 mm,体宽1.08 ± 0.02 mm,体壁透明,有腺体分布,消化道颜色明显,呈红褐色。第2 腹节之后各腹节脂肪体颗粒密布(图2B)。3 龄幼虫头壳宽为0.69 ± 0.01 mm,体长3.05 ± 0.04 mm,体宽1.18 ± 0.04 mm,体肥硕,体壁较2 龄厚,取食量增大,皮下白色脂肪体颗粒变大(图2C)。4 龄幼虫头壳宽为0.92 ± 0.01 mm,体长4.97 ± 0.12 mm,体宽1.97 ± 0.13 mm,体粗短,两端钝圆,头常缩于前胸内,体壁变厚呈乳白色,分节十分明显,消化道发黑,吐丝结茧后体壁变为微淡黄色(图2D,E)。每次蜕皮时间间隔约为2~3 d,整个幼虫期为9~10 d。吐丝结茧化蛹前的老熟幼虫头壳特征明显,由一对触角、口侧片、一对上颚、吐丝器、口后片、口后片距、茎节骨片、下唇和下唇骨片组成(图2F)。这些特征可作为识别姬蜂幼虫种类的重要鉴定依据。

图2 幼虫期Fig.2 Larval stage
老熟幼虫变为短粗继而细长状的预蛹,乳白色,复眼从无色渐变为红色,腹部脂肪体颗粒密集(图3A,B)。约3 d 后老熟幼虫蜕皮化蛹,蛹为离蛹。在预蛹和蛹期虫体发育迅速,以色素的沉积最为明显。初蛹乳白色,复眼和单眼红棕色,雄性外生殖器于腹部末端突出,雌性的产卵器弯曲粘连在腹背部上方,雌、雄蜂分化明显(图3C,D)。蛹的中期,以胸部变黑为主(图3E,F);蛹的后期,以触角、腹部和翅芽变黑为特征(图3G,H)。总之,在蛹的发育过程中,先是复眼和单眼的颜色逐渐由红色变至黑色;接着中胸和上颚变褐变黑;随后前、后胸背板和腹板变黑,头部变黑;接下来,触角由无色透明状变为浅褐色至黑色,腹部后端变黑;最后翅由透明变为黑色,翅脉可见,意味着即将羽化为成虫。当蛹完成色素的沉积和形态转化之后,翅发育完成,破茧而出,羽化为成蜂。预蛹和蛹期共计12~15 d。

图3 预蛹和蛹Fig.3 Prepupal and pupal stage
通常,雄蜂比雌蜂早羽化2~3 d。成蜂体型较大,具明显性二型现象(图4A~D)。雌蜂体长9.00 ~11.00 mm,黑色(图4A,B)。触角31~33 节,鞭节第5~9 节、腹部第7背板端部中央具白色大斑(图4A,F),产卵管鞘粗壮,长为2.50 ± 0.05 mm,为第1 腹节的1.6 倍;产卵针细长形似针头,长3.70 ± 0.02 mm,其腹瓣亚端具有7 条纵脊( 图4E )。 雄蜂黑色, 体长8.50~10.30 mm,稍小于雌蜂,但比雌蜂更具光泽(图4C,D)。触角32~35 节,在鞭节第15~20 节有角下瘤(图4G);小翅室大,呈五边形,横窄,前翅第2 回脉位于小翅室内侧,约在内侧0.3 处相接(图4H)。

图4 成虫特征Fig.4 Morphological characters of adult wasps
雌蜂从产卵到羽化为成蜂完成一个世代共历经23~27 d。黄杨斑蛾田猎姬蜂生长发育所需营养完全由寄主蛹提供,一头寄主茧里只能出一头蜂,一般来说,蛹体越大,羽化出的蜂也相对较大。
2.2 生殖系统构造
2.2.1 雌蜂 雌蜂生殖系统构造主要由体内的1 对卵巢、1 对侧输卵管、中输卵管、1 个受精囊和外部的产卵器组成(图5A,B)。此外还有杜氏腺、毒囊和毒腺等与产卵寄生相关的结构。卵巢位于消化道两侧向后背方延伸,占据了腹部内绝大部分的空间。每侧卵巢由4~6 根卵巢管组成,每雌卵巢管数9~11 根,以10 根居多(图5A)。当卵子发生完成时,每根卵巢管基部至少含一粒卵黄蛋白丰富的成熟卵。每根卵巢管的滋养细胞与卵母细胞交替排列,属典型的多滋式。产卵结束后,卵巢管萎缩。侧输卵管位于卵巢萼下部,两侧的侧输卵管汇入中输卵管,侧输卵管及中输卵管很短(图5A)。中输卵管后部接入生殖腔,生殖腔前端膨大呈球形,其余部分管状,经产卵孔向体外开口。

图5 雌蜂生殖器官Fig.5 Female reproductive organs of Agrothereutes minousubae
产卵器很发达,由3 对瓣状结构组成(图5B)。1对背瓣组成产卵器鞘,内瓣和腹瓣组成产卵构造为产卵针,锯齿状结构以利于刺破寄主茧壳(图5B)。
2.2.2 雄蜂 雄蜂生殖系统由内部的1 对愈合的精巢、2 根细长的输精管、1 对贮精囊和射精管,以及外部的交配器组成(图6A)。精巢呈淡黄色圆球状。精巢基部侧下端各伸出1 条细长的输精管,此为输精管上部,其末端接入同侧贮精囊近基部。输精管下部从贮精囊下端伸出,较粗,下端汇入射精管。
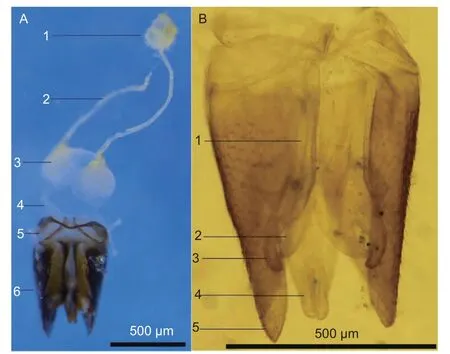
图6 雄蜂生殖器官Fig.6 Male reproductive organs of Agrothereutes minousubae
交配器由阳具和抱握器组成,其中阳具由阳茎和阳茎基环组成,阳茎由阳茎内突、阳茎基腹铗、阳茎端和阳茎基侧突组成。抱器背突形成抱握器(图6B)。
2.3 生殖行为
交配前雄蜂会有求偶行为,雌蜂若接受雄蜂,便与之交配,雄蜂完成授精过程。雌蜂无论是否交配均可产卵寄生大叶黄杨斑蛾的蛹,交配后雌蜂所产后代的性比接近1∶1。
2.3.1 求偶行为 将羽化当天或羽化1~2 d 的雌、雄蜂进行配对时,雄蜂很快表现出明显的振翅求偶行为,通常在雌蜂后面追逐,若雌蜂不接受,则雄蜂飞离原地,表明求偶失败。再将该雄蜂放入其他有雌蜂的装置中,继续观察其求偶行为,观察并记录,直至求偶并交配成功(图7, 图8)。

图7 交配中的田猎姬蜂Fig.7 A pair of Agrothereutes minousubae in mating
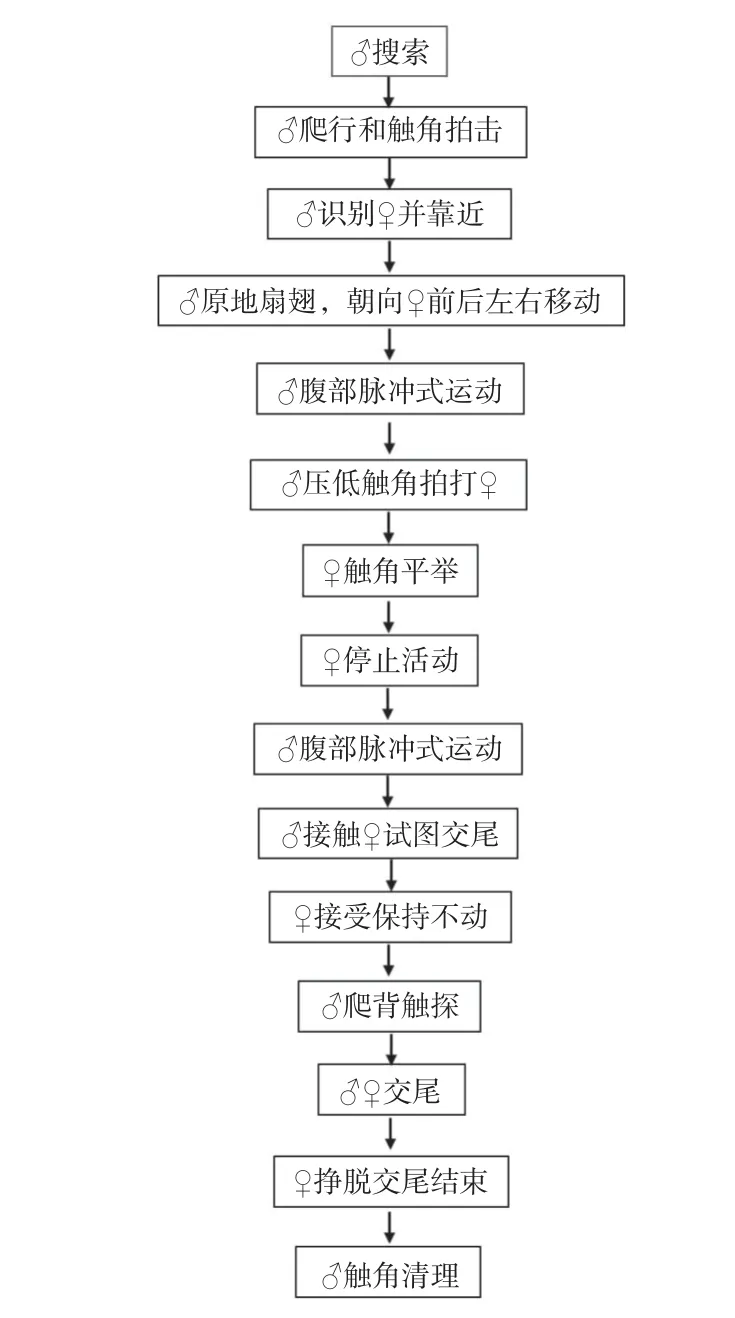
图8 求偶与交配行为Fig.8 Courtship and mating behivour sequence of Agrothereutes minousubae
2.3.2 交配行为 交配前,雄蜂用触角不断拨动雌蜂触角内侧近端部位置,以此向雌蜂传递信息,这一过程大概持续15 s;当雌蜂感受到雄蜂发出的信号并接受时,保持身体不动。此时,雄蜂伏在雌蜂体背,用前足和中足紧紧抱住雌蜂,后足支撑身体,腹部弯曲呈弓形,交配持续时间约为3~7 min,平均交配时间约5 ± 2 min。雌蜂先挣脱离开预示交尾结束。交配结束后,雌蜂飞离,雄蜂进行触角清理行为,为下一次求偶交配行为做准备。若雄蜂求偶失败,则无法靠近雌蜂;或雌蜂离开;或当雄蜂试图交尾时,雌蜂挣脱雄蜂的爬背并逃离。通常,一头雄蜂一生可与多头雌蜂交配,但雌蜂一生只交配一次。
交配后雌蜂即可产卵寄生,雌蜂初接触寄主时通常会有假寄生行为发生,待交配后2~3 d 且熟悉寄主后,便可大量产卵寄生,此外,雌蜂未与雄蜂交配也可自行产卵,营孤雌生殖,所产后代全为雄性。
2.3.3 产卵寄生行为 雌蜂实施一次成功的产卵寄生行为要经历搜索、定位、识别、蜇刺、产卵、清理等一系列过程。首先雌蜂在对寄主的搜索过程中,触角始终处于非常活跃的状态,不断敲击拍打;当找寻到寄主后,先用触角不断拍打寄主茧,随后鞭节弯曲呈直角,利用触角端部感受器感知寄主状态,同时利用产卵器刺破寄主的茧壳刺探寄主是否适宜产卵;雌蜂实施寄生前,常伴随有产卵器穿刺寄主茧壳的动作,但无卵产出。这表明该蜂通过此行为进行寄主的识别,以判断寄主是否适合产卵寄生。经此学习经历,随后便可正常产卵寄生(图9)。

图9 产卵寄生行为Fig.9 Ovipositional behavior of Agrothereutes minousubae
产卵寄生时,雌蜂左右翅重叠,腹部拱起,将产卵器刺入寄主茧内,腹部末端有规律地伸缩(图10)。由于寄主茧较厚,蜂产1 粒卵的时间5~40 min 不等,将卵产附于寄主蛹体表(图11A),产卵位置以蛹侧面居多。也偶见过寄生行为,即将2 粒或以上的卵产在同一寄主蛹上,数量高达5 粒。同时,伴随杀卵现象,即先产的卵被随后产卵的雌蜂产卵器刺破。但无论过寄生程度如何,1 个大叶黄杨斑蛾茧最多只能育出1 头蜂。产卵结束后,雌蜂会用足对触角和产卵器进行清洗和梳理,为下次产卵寄生做准备。该姬蜂作为大卵型外寄生蜂,日产卵量可达7 粒,一生可产20 粒卵,产下的卵有寄主茧保护。

图10 产卵于大叶黄杨斑蛾茧内Fig.10 Agrothereutes minousubae parasitizing on the cocoon of Prgeria sinica
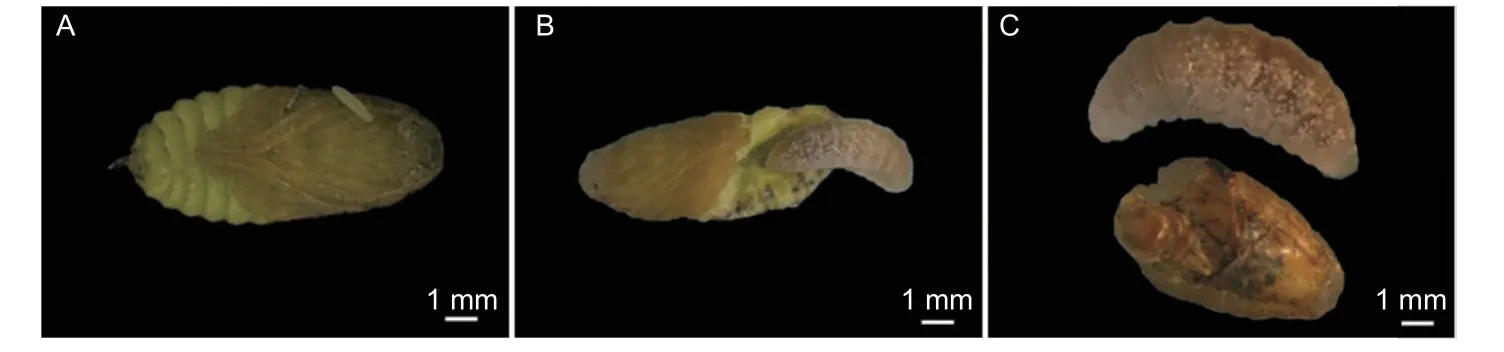
图11 产卵部位与幼虫取食Fig.11 Egg depositing position and of Agrothereutes minousubae
待幼虫孵出后开始取食蛹,蜂幼虫用1 对较发达的上颚在蛹体腹部位置先刮破一小口,吸食寄主蛹血淋巴等作为自身发育的营养来源(图11B),取食持续两周左右,将蛹内物质吸食尽只剩表皮,自身也发育为老熟幼虫(图11C)。
停止取食后,老熟幼虫开始在寄主茧内吐丝结茧,并以老熟幼虫的虫态滞育,待温度和光周期适宜时,滞育被打破,老熟幼虫开始化蛹,适时咬破寄主茧羽化钻出。
3 讨论
3.1 黄杨斑蛾田猎姬蜂胚胎发育需48~50 h,幼虫有4 个虫龄,完成个体发育历经23~27 d
昆虫的个体发育始于卵期的胚胎发育,早期胚胎发育模式基本一致,由合核分裂开始,但在后期胚盘的形成以及体节分化等阶段表现出多样性。而就完成胚胎发育的时间而言,通常长胚带昆虫即滋养型要比短胚带昆虫即无滋型的胚胎发育快得多。除了产卵于寄主的位置不同,内寄生蜂胚胎发育可同时利用自身卵内营养物质积累和寄主血淋巴的营养完成[16],而外寄生蜂的胚胎发育则完全依赖自身卵内营养物质的贮备。川皮硬肿腿蜂(Scleroderma sichuanensisXiao)在26 ℃培养条件下,产卵60~70 h 后,幼虫孵化[17]。而在24 ℃下,管氏肿腿蜂(S.guaniXiao et Wu)的胚胎发育却历经140 h[18],而同为外寄生蜂的黄杨斑蛾田猎姬蜂胚胎发育仅用48~50 h。为何不同寄生蜂种类卵期发育时间存在明显差异,其机制有待深入研究。因外寄生蜂胚胎发育的观察需卵量较大,而且需要收集不同发育时期的卵来进行固定、染色,甚至切片,故而本研究只明确了黄杨斑蛾田猎姬蜂从产卵到幼虫孵化完成胚胎发育所需时间。相对于绝大多数寄生蜂,黄杨斑蛾田猎姬蜂卵较大,长约1.57 ±0.07 mm,通过其卵内物质与胚胎发育期关系的研究对揭示寄生蜂胚胎发育期差异而言是一个理想的材料。已有的外寄生蜂胚胎发育研究表明,受精卵首先经历合核分裂、胚盘形成;接着胚带形成及胚层分化;与此同时,胚胎分节、原头原躯化、器官与系统形成,最后附肢形成、胚动、胚胎闭合,直到幼虫孵化[17~20]。
黄杨斑蛾田猎姬蜂幼虫经历3 次蜕皮,有4 个虫龄,特别对末龄老熟幼虫头壳的分类特征进行了细致观察,可与其他姬蜂加以区别。蛹期经历色素的快速沉积转运,其转化过程与半闭弯尾姬蜂(Diadegma semiclausum(Hellén))蛹期色素变化基本一致[21]。黄杨斑蛾田猎姬蜂成虫性二型现象明显,其前翅小翅室大,呈五边形,横窄,前翅第2 回脉位于小翅室内侧,约在内侧0.3 处相接。小翅室和第2回脉特征与赣田猎姬蜂和基田猎姬蜂有明显差异[3],这些可作为田猎姬蜂属不同种类分类的重要鉴定依据。而角下瘤不仅是雄性的识别特征,更是在信息素释放、参与求偶、发挥不同的生态功能中起重要作用[22-23]。至于黄杨斑蛾田猎姬蜂角下瘤的超微形态结构和生理功能还有待研究。
黄杨斑蛾田猎姬蜂个体发育的研究为进一步深入探讨该蜂的遗传学机制提供了形态学基础,也为今后开展对该天敌的识别、保护和引种提供了基础信息。
3.2 雌蜂卵巢管数量9~11 根,为多滋式,产卵器细长针状。雄蜂精巢愈合,交配器发达
天敌寄生蜂生殖力大小决定其控害能力,而生殖力与卵巢管数量关系密切。已有的研究表明,多数情况下,卵巢管数量越多其生殖力越强[24]。黄杨斑蛾田猎姬蜂卵巢管数量9~11 根,为多滋式,生殖力较强。相比于雌蜂,目前对雄蜂生殖系统构造的研究较少。精巢和贮精囊结构的研究可为深入研究雄蜂交配次数与性别决定提供重要的理论依据。交配器的形态学研究可为同属不同种类,特别是近缘种的区分提供鉴定依据。
3.3 雌蜂一经羽化即可交配,羽化后2~3 d 产卵,日产卵量达7 粒
寄生蜂生殖行为复杂多样,直接影响控害能力的发挥和其自身种群的维持和增长。因此,研究寄生蜂生殖行为,如求偶行为、交配行为和产卵行为等可为更好地利用寄生蜂这一天敌资源发挥其控害作用提供理论基础。大多数寄生蜂体型较小,甚至微型,不足1 mm,而黄杨斑蛾田猎姬蜂体型较大,这有利于生殖行为的准确观察和记录,不失为一种研究寄生蜂生殖行为学的极好材料。田间调查发现,雄蜂比雌蜂早2~3 d 羽化,这与室内个体发育过程相吻合。雌蜂一经羽化即可交配,通常一头雌蜂一生只交配一次,而一头雄蜂可以和多个雌蜂交配。羽化后2~3 d 雌蜂产卵,日产卵量达7 粒。黄杨斑蛾田猎姬蜂存在过寄生行为与杀卵现象,这在其他外寄生蜂中也有报道[23],对其产生的机制的还有待探明。因行为学内容本文采取人眼观察记录,结合相机录像方式,为保证精确,且雌蜂只交配一次的特性,重点观察了雌雄蜂单对的求偶和交配行为。然而,寄生蜂求偶与交配行为是非常复杂的过程,当1∶n,n∶1,n∶ n 等情况下涉及性别竞争、交配干扰时雌雄蜂的交配行为如何发生还有待深入研究。
4 结论
本文通过对河南新记录种黄杨斑蛾田猎姬蜂个体发育、生殖系统构造及生殖行为的系统性初步研究,结合室内饲养和室外调查,丰富了该蜂生物学特性的基本信息。得出以下结论:黄杨斑蛾田猎姬蜂为大卵型、卵育型外寄生蜂,胚胎发育需48~50 h;幼虫有4 个虫龄,幼虫期历经9~10 d;预蛹期及蛹期12~15 d;待成蜂羽化完成一个世代历经23~27 d。雌雄蜂生殖系统构造属典型的膜翅目昆虫类型。同时本文观察并补充了黄杨斑蛾田猎姬蜂求偶、交配及产卵寄生行为学方面的空白,为更好地实施人工保护和发挥该蜂对大叶黄杨斑蛾的控害潜能提供了依据,为实现园林植物的绿色保护奠定了基础。
