种公鸡繁殖性能调控机制的研究进展
2024-01-13孙研研陈继兰
孙研研,陈继兰
[中国农业科学院北京畜牧兽医研究所(畜禽生物育种全国重点实验室),北京 100193]
种公鸡于鸡繁育体系具有至关重要的作用。种鸡采用人工授精时,种公鸡和种母鸡的比例高达1∶80~1∶100。种鸡采用本交交配方式时,生产后期种公鸡交配能力和精液品质的下降也是产业重点关注的种鸡管理难点。因此,种公鸡的繁殖性能是维持种群高效繁育的关键。如何选育提高并维持种公鸡较高的繁殖性能成为科研和育种中的热点话题。
种公鸡繁殖性能受到多方面的影响。在遗传方面,主要精液品质的遗传参数估计结果表明其具有一定的遗传变异基础,通过全基因组关联分析、蛋白质组学和转录组学等多组学分析鉴定与精液量、精子活力等精液品质相关的候选基因和关联位点,为精液品质和繁殖性能的遗传选育提供分子技术手段。在营养因素方面,日粮中的蛋白含量低、能量不足、维生素及矿物质等营养元素的缺乏会直接导致种公鸡精液品质的降低。维生素、微量元素、植物油、矿物质、氨基酸及其衍生物等生物活性物质可在一定程度改善种公鸡精液品质。在饲养模式与环境条件方面,养殖模式和光照等环境因素也会影响种公鸡性成熟和繁殖性能的发挥。本文介绍了公鸡的生殖生理调控机理,总结了遗传、营养、饲养模式与环境条件等因素对公鸡精液品质和繁殖性能的影响,以期为种公鸡繁殖性能的选育、管理和性能调控提供参考。
1 公鸡的生殖生理特征
相对于哺乳动物,家禽拥有较为特殊的生殖系统,了解其生殖基础是提升其繁殖效率的重要前提。
1.1 公鸡的生殖系统
公鸡生殖系统由睾丸、附睾、输精管和交媾器4 部分组成。睾丸是精子发生和性激素分泌的主要器官。睾丸发育主要发生在2~15 周,在此阶段,睾丸重量没有明显的增加,主要是精原细胞增殖,精原细胞的数量决定了睾丸生成精子的能力。15 周龄后睾丸增重明显,曲细精管管腔内开始出现成熟精子。睾丸重和精液量在28~30周龄达到高峰,受精率最高。35 周龄后睾丸发生萎缩,精液量减少[1]。成年公鸡睾丸主要由大量的曲细精管组成,曲细精管紧密排列有精原细胞、精母细胞、精细胞、精子细胞和支持细胞,形成生精上皮。管腔内有精子,曲细精管外壁之间有狭长的间质细胞,支持细胞作为营养细胞为精子发生提供合适的微环境,支持精子发育。间质细胞能产生雄激素,参与睾丸发育和精子发生的调控。
附睾紧贴在睾丸两侧,由睾丸网、输出管和附睾管构成,参与精子的运输、成熟、重吸收睾丸网液和浓缩精液等生理过程[2]。输精管位于睾丸背部,前端与附睾相连,尾端弯曲膨大并埋于泄殖腔内,终端变直开口于泄殖腔两侧,并向泄殖腔内突出形成射精管。公鸡没有真正的阴茎,只有退化的交媾器,由位于肛门腹侧缘的生殖突起(阴茎体)、淋巴襞等结构组成。在交配或人工采精时,阴茎体合拢成纵沟翻出泄殖腔,精液流入纵沟而排出体外。
1.2 精子的生成与成熟
精子发生是一种复杂的细胞分化过程,包括精原细胞的有丝分裂增殖、初级精母细胞减数分裂为圆形精子细胞、圆形精子细胞向精子细胞的形态分化3 个阶段[3]。研究人员根据顶体发育和核结构的变化将家禽圆形精子细胞向精子细胞的形态变化过程又细分为12个步骤(图1)[4]。步骤1~3为高尔基体期,主要进行染色质凝结,形成顶体囊泡;步骤4~6 为加帽期,精子细胞核继续浓缩,形成核内管,顶体形成丝状结构的原基;步骤7~9 为顶体期,细胞核逐渐拉长,鞭毛被纤维鞘包裹;步骤10~12 为成熟期,细胞核继续拉长,形成成熟精子。
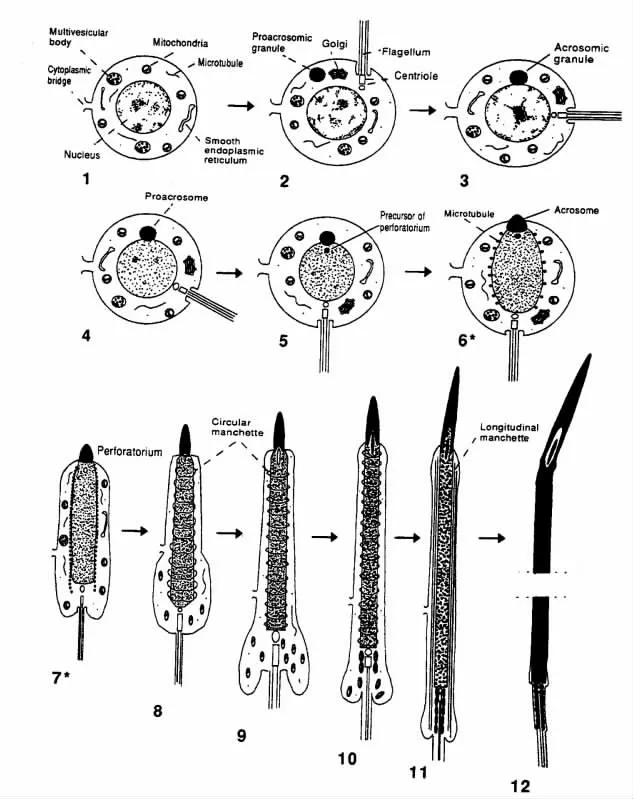
图1 家禽圆形精子细胞向精子细胞发育过程[4]
刚从睾丸排出的精子功能尚不成熟。当哺乳动物精子通过附睾时进一步成熟,获得受精能力。家禽附睾发生一定退化,但近年来越来越多研究表明家禽附睾和输精管对精子成熟有调控作用[2,5]。
精子发生呈周期性、连续不断的特征。鸡的精子发生周期仅为14d,显著低于小鼠的35d 和人的64d。一方面可能是由于禽类精原细胞增殖过程中的有丝分裂少于哺乳动物[6],另一方面家禽精子成熟主要在输精管内,精子自睾丸经输精管到泄殖腔只需要24h,成熟时间也显著短于哺乳动物。
1.3 鸡的精子和精液特征
精子一般分为头部、颈部和尾部三部分,尾部又分为中段、主段和末段。与人类和家畜等哺乳动物精子的扁圆形头部不同,鸡精子头部呈长圆柱状,最大直径仅为0.5~0.7μm,尾部长达80μm[7]。顶体位于精子头部的顶端,内有大量与受精相关的酶。精子细胞核染色质高度浓缩,线粒体聚集于中段形成线粒体鞘,是精子运动的主要能量来源(图2)。

图2 鸡精子及各段超微结构电镜图[1]
精液中除了精子还有精浆,鸡精浆占精液体积的90%以上。由于缺乏精囊腺、前列腺和尿道球腺等哺乳动物的副性腺,附睾不发达,禽类精浆主要成分与哺乳动物有较大差异,主要包括水、糖类、脂类、蛋白质、有机酸、有机碱和无机离子等。精浆具有为精子提供充足的能量、吸收精子代谢产物、缓冲内外环境刺激、维持精子生存环境稳态、辅助精子进入雌性生殖道等多重功能。
2 影响公鸡繁殖性能的因素
精液品质与受精率呈显著相关[8],是对种公鸡繁殖能力的间接度量指标。因此,影响睾丸发育、精子生成等诸多因素均可调控公鸡繁殖性能,主要包括遗传、营养和管理等因素。
2.1 遗传因素
遗传参数估计的结果表明,主要精液品质性状是可遗传性状,受遗传变异的影响。Hu 等[9]估计地方品种北京油鸡43 周龄主要精液品质性状遗传力,发现精子活率、精子活力和精子畸形率3 个性状为高遗传力(h2=0.52、0.85、0.60),精液量、精液颜色和精子密度为中等遗传力性状(h2=0.28、0.19、0.12),精液pH 值的遗传力较低(h2=0.03)。Bongalhardo 等[10]估计白来航公鸡26周龄精液量、精子密度和精子活力性状遗传力分别为0.27、0.34 和0.26。Kabir 等[11]估计罗德公鸡精子活力、精子活率、精液量、精液颜色、精子密度和精子畸形率的遗传力分别为0.82、0.33、0.45、0.55、0.52 和0.42。Gebriel 等[12]估计埃及地方品种鸡46 周精液量、精子密度、精子活力、精子活率、精子畸形率和精液pH 值的遗传力分别为0.28、0.11、0.32、0.25、0.12 和0.25。整体来看,公鸡精液品质性状属于中等偏高遗传力性状,其中精子活力、精液量、精子活率在不同品种中都具有较高的遗传力,但精子密度、精液pH的遗传力普遍较低。
基于精液品质性状的可遗传基础,研究人员采用不同的技术手段解析精液品质的遗传调控机制,其中最为关注的是精液量和精子活力性状。Zhang 等[13]通过全基因组关联分析检测到7 号染色体上13.82~16.12Mb 与白羽肉种鸡精液量性状显著关联的位点,并筛选到脂肪酸结合蛋白1(Fatty Acid Binding Protein 1,FABP1)、氧固醇结合蛋白样6(Oxysterol Binding Protein Like 6,OSBPL6)和精子特异性抗原2(Sperm Specific Antigen 2,SSFA2)等重要候选基因。精子的顶体膜和其他组分中含有多种精子相关抗原蛋白,其中精子相关抗原6(Sperm Associated Antigen 6,SPAG6)蛋白最早在衣藻的纤毛中发现,参与纤毛形成和运动等过程。随后在动物的睾丸中也鉴定到SPAG6 蛋白,并确定为一种精子鞭毛中央轴丝微管控制鞭毛运动和稳定性的重要蛋白。Sun 等[1]通过蛋白质组学鉴定到弱精子症(精子活力低)与正常公鸡睾丸中差异表达的SPAG6 等93 个蛋白,是细胞骨架和鞭毛的成分,并参与精子受精、蛋白质转运和细胞蛋白质定位相关的生物学过程,并进一步证实了SPAG6 在弱精子症睾丸中表达下调,且其降低趋势与睾丸发育和衰退速度一致。而SRY 盒转录因子5(SRY-box Transcription Factor 5,SOX5)也参与调控了SPAG6的下调表达。通过对北京油鸡SPAG6 基因多态性与精液品质和性发育相关性状进行关联分析发现,SPAG6 基因SNP 突变与其精液品质显著相关[14]。
近年来,非编码RNA(包括lncRNA,miRNA 和piRNA 等)对精液品质的调控作用也成为研究热点[15,16]。Liu 等[17]在地方品种北京油鸡公鸡睾丸样本中鉴定到了518 个已知miRNA,同时预测到了106 个新miRNA,gga-miR-155 在低精子活力公鸡睾丸的表达量是高精子活力个体的2.1倍。通过生物信息学工具进行靶基因的预测发现,gga-miR-155 能通过靶向结合钾电压门控通道亚家族A 成员1(Potassium voltage-gated channel subfamily A member 1,KCNA1)基因参与调控鸡精子活力。Xing 等[18]在海兰褐父母代的弱精子症和正常公鸡中鉴定出302 个差异表达基因和13 个差异表达miRNA,生物信息学分析发现这些差异表达基因与类固醇激素的生物合成有关。Li等[19]对比了高精子活力和低精子活力北京油鸡精浆蛋白质表达谱的差异,发现8 个与糖酵解过程相关的蛋白酶,包括3-磷酸甘油醛脱氢酶(Glyceraldehyde-3-phosphate Dehydrogenase,GAPDH)和 己糖激酶3(Hexokinase 3,HK3)以及3 个线粒体功能相关的蛋白质,包括线粒体磷酸激酶(Creatine Kinase,Mitochondrial 2,CKMT2)、细胞色素c(Cytochrome c,CYC)和精子发生相关蛋白18(Spermatogenesis Associated 18,SPATA18),在低精子活力组高表达的这些蛋白质可能来源于精子鞭毛结构损伤,其在精浆中的高表达可能意味着精子的结构损伤是造成活力下降的主要原因。
除了睾丸中基因和蛋白的表达对精液品质有调控作用外,精子排出睾丸后的成熟过程依然发挥重要的调控作用[20]。其中外泌体是由细胞分泌的直径为30~150nm 的小囊泡,可以介导细胞间的物质转运,在配子发育和成熟等生理过程中发挥重要调控作用[20]。Cordeiro 等[21]和Luo 等[22]鉴定了鸡精浆外泌体的蛋白质表达谱,将高精子活力组的外泌体孵育低精子活力组的精子,可在一定程度上改善精子运动和受精[21]。Han 等[23]鉴定了鸡精浆外泌体的miRNA 表达谱,发现高表达的miRNA 可以靶向调控精子和母鸡贮精腺细胞中的mRNA,通过调控精子成熟和雌性生殖道免疫抑制等过程影响精子活力。
2.2 营养因素
2.2.1 能量
能量是保证种公鸡正常生长发育的基础。长期饲喂低能量日粮会导致种公鸡精子密度和活力下降。饲喂高能日粮容易导致公鸡体重超标,本交笼的鸡交配困难,精液品质下降,受精率降低[24]。不同品种公鸡在不同生长发育时期的能量需求存在差异。姚会等[24]在海兰褐种公鸡上的试验表明,11.7~11.9MJ/kg 为22 周龄公鸡的适宜能量水平,其精子活力和精液量最佳[24]。王竹伟等[25]指出50 周龄北京油鸡公鸡的适宜能量水平为11.72MJ/kg。高小芳等[26]研究表明,32周龄广西三黄鸡种公鸡的适宜能量水平为11.3~11.7MJ/kg。
2.2.2 脂肪酸
鸡精子质膜中含有大量花生四烯酸、二十二碳四烯酸等多不饱和脂肪酸(Polyunsaturated Fatty Acid,PUFAs),与膜流动性、顶体反应和精子活力紧密相关,主要体现在PUFAs 可以穿透精子细胞膜,提高精子质膜的可扩展性,保持其结构和功能的完整性,增强顶体膜的渗透阻力[27]。由于家禽体内缺少脂肪酸去饱和酶,PUFAs 或其前体主要来源是饲料,因此,饲料中添加PUFAs 对提升公鸡精液质量具有重要意义[28]。当在公鸡的日粮中添加富含2%n-3 PUFA 的鱼油40d 后,精子活力、质膜完整性及脂质过氧化反应等精液品质均优于未添加组[29]。47 周龄Ross 种鸡饲料中持续13 周添加2%鱼油也可以改善其精液质量和繁殖性能,添加鱼油组的睾丸激素、精子活力和运动性以及受精率均显著提高[30]。但由于鱼油成本高,在饲料加工和生产过程中也容易变质[31],一些替代品也在不断挖掘。如亚麻籽油作为α-亚麻酸的丰富来源,在酶的作用下可以通过去饱和和延伸过程将其转化为二十碳五烯酸(Eicosapentaenoic acid,EPA)和二十二碳六烯酸(Docosahexaenoic Acid,DHA)[32]。在公鸡 日粮中添加2%的亚麻籽油可以改善老龄公鸡的精液质量和繁殖性能,同时可以增加其EPA 和DHA 含量[33]。而且亚麻籽油增强精液量、精子活力和精子总数是通过上调参与类固醇生成的限速酶(如类固醇生成急性调节蛋白)的mRNA 表达水平实现的[34]。
2.2.3 氨基酸及其代谢物
一些重要的氨基酸及其代谢产物参与了睾丸发育、精子生成和精子抗氧化等功能的调节,当日粮中氨基酸不能满足机体需要时,可能会导致繁殖性能下降。在55 周龄肉种鸡日粮中添加200~300mg/kg D-天门冬氨酸可以提升睾酮(Testosterone,T)水平以及精子密度、质膜完整性、精子活力和受精能力[35,36]。精氨酸主要参与精子蛋白质鱼精蛋白的合成,能有效提高精子质量,增强精子运动能力。除此之外,精氨酸还可以促进生长激素、雄性激素正常分泌,以及精子正常生成。家禽无法通过鸟氨酸循环等生化途径合成精氨酸。有研究表明,连续3 周在52 周龄Ross 肉种鸡日粮添加0.68%的L-精氨酸可以提升T 水平、精子质量和生精能力[37]。胍基乙酸是由精氨酸生物合成的,用来产生肌酸。肌酸被磷酸化后产生磷酸肌酸并产生能量[38]。当添加1200mg/kg 的胍基乙酸时,不仅可以提高肌酸合成效率,保障精子代谢能量,同时可以提升精液量和精子活力,也可以作为抗氧化剂保护精子质膜完整性,有效改善生精小管上皮的厚度,增加精原细胞数量,显著提高公鸡的繁殖能力[39,40]。L-肉碱由赖氨酸和蛋氨酸生成,可向线粒体运输长链脂肪酸参与脂肪代谢,也可以通过调节过氧化氢酶和谷胱甘肽过氧化物酶(Glutathione Peroxidase,GSH-Px)等抗氧化酶的活性来防止脂质氧化损伤[41,42]。在白来航公鸡从出雏开始在饲料中长期添加125mg/kg 的L-肉碱可以稳定的提高精子密度[43,44]。而在36 周龄种鸡中饲料中添加125mg/kg 的L-肉碱可以显著提升T 水平和受精率[42]。
2.2.4 维生素
维生素E 又名生育酚,作为重要脂溶抗氧化剂在PUFAs 过氧化反应中起着至关重要的保护作用[45]。在饲料中额外添加天然维生素E 可以显著影响精子密度、活力、活率及受精率[46]。罗曼褐种鸡日粮中添加100mg/kg 的维生素E 可以增加精液量,精子密度和精子活力[47],地方品种种鸡日粮中添加160mg/kg 维生素E 可以提高受精率[48],而Ross 种公鸡需要将添加量提高到200mg/kg 才能有类似的效果[33]。除了单独添加维生素E,维生素E 与鱼油、亚麻籽油以及大豆磷脂一起添加进饲料中均可以提高精液质量及繁殖性能[6,9,13]。
2.2.5 微量元素
硒作为与禽类生殖功能联系最密切的一类重要微量元素,存在于所有家禽睾丸与精子中,是GSH-Px 的组分,发挥抗氧化作用[47,49,50]。在30 周龄洛岛红公鸡饲料中添加0.3mg/kg 的硒可以提高睾丸、精子和肝脏中GSH-Px 的活性,对精子发生、精子质量和受精率提升等均有效果,在火鸡、鹅、鹌鹑和鸽等其他家禽中的效果也得到充分验证[51-53]。
2.3 饲养模式与环境条件
2.3.1 饲养模式
种鸡的饲养模式主要有单笼、本交笼和平养。饲养模式对公鸡精液量和质量也具有一定的影响。Du 等[54]对比70~210 日龄罗曼父母代种鸡在单笼(50cm 长、40cm 宽、45cm 高)、大 笼(200cm 长、100cm 宽、80cm 高,无母鸡)和本交笼(200cm 长、100cm 宽、80cm 高,公母配比9∶81)中饲养对公鸡性成熟和精液品质等的影响,发现本交笼公鸡精液量、精子密度和精子活率高于其他两组,而且受精蛋孵化率也显著提高,这可能与本交笼公鸡在母鸡的视觉刺激和激素刺激下,其促进垂体分泌促卵泡激素(Follicle-Stimulating Hormone,FSH)、促黄体生成素(Luteinizing Hormone,LH)和T 等激素水平较高有关[54,55]。随着劳动力紧缺和人工成本的不断上升,种鸡场人工授精的成本不断提升。蛋鸡本交笼养模式作为一种新的养殖模式,因具有节约成本,以及便于实现饲养管理自动化、标准化和福利化等优点,正迎来巨大的发展契机[56]。本交笼养模式下每个笼为一个饲养单元,每笼目前常见的饲养量为50 或者100 只鸡,公母比例为1∶9~1∶10。有研究表明,在50 和100 只种鸡的本交笼中,同一笼内不同公鸡的遗传贡献率差异显著,剔除遗传贡献率最低的1 只公鸡,不影响受精率,且可以降低饲养成本。公鸡有效精子数及血清中的FSH、三碘甲状腺原氨酸(Triiodothyronine,T3)水平与公鸡的遗传贡献率的相关系数分别为0.79,0.80 和-0.80,可以作为潜在的选择优秀种公鸡的指标[57]。
2.3.2 温度
种公鸡养殖环境的温度条件也会对其精液品质和繁殖力产生一定的影响。高温是一种重要的应激源,当环境温度超过27℃时,鸡群会产生热应激,特别是在湿度较大时热应激更大。热应激一方面严重抑制鸡采食行为和机体代谢,摄入营养不足会导致公鸡繁殖性能下降。另一方面,高温环境直接导致公鸡体温和睾丸温度升高,生精细胞数量减少[58],使精子发生的成熟和储存受到影响,精子DNA 完整性受到破坏[59];睾丸重和精液量降低,精液成分也发生变化,导致精子密度、活力和活率的下降[60,61],在雌性生殖道的存活时间显著降低,凋亡增加[61,62]。此外,睾丸中的支持细胞对生殖细胞的生长发育很重要,FSH 可以刺激支持细胞分泌结合T 的雄激素结合蛋白,热应激可能通过降低FSH 对支持细胞产生负面影响,从而抑制精子发生[61]。温度对公鸡繁殖性能的影响不只局限于性成熟后。有研究表明,在蛋种鸡孵化的第15~17 天每天进行3h 的40℃热刺激,与对照组相比,成年后公鸡的精子活率、抗氧化能力显著提高,热应激蛋白基因的表达量显著降低[63]。
2.3.3 光照
光信号以周期变化、强度和光波长等属性被光感受器所感知,并转变成生物学信号,调节动物的生理和行为。家禽的繁殖活动受 “下丘脑-垂体-性腺轴” 的调控。下丘脑分泌促性腺激素释放激素(Gonadotropin-releasing Hormone,GnRH),促进垂体分泌FSH 和LH。LH 作用于间质细胞膜上的LH 受体,促进T 合成。T 与支持细胞的雄激素受体结合,刺激雄激素分泌,进而刺激精子发生和性征的维持。FSH 则与支持细胞结合,激活腺苷环化酶,促进环腺苷酸(Cyclic Adenosine Monophosphatec,AMP)形成,激活蛋白激酶的活性,促进支持细胞合成和分泌促进曲细精管发育和精子发生的活性物质。禽类对光照敏感,光照可以直接作用于下丘脑或者通过视网膜感受器产生光信号传递至下丘脑[64],引起血液LH 和FSH 浓度变化,影响生殖系统功能。因此,光照也是影响种公鸡繁殖性能的一个重要环境因素[65]。在实际生产中,种公鸡与种母鸡饲养于同一环境中,一般是育成期每天8h 光照,20 周龄左右延长至每天12h,随后每周逐步增加光照时长,直至25 周龄前后达到每天光照15h 左右,从而刺激其性成熟一致[66]。但也有研究发现,育成期每天光照4h 的公鸡性成熟、睾丸重量和精子量与每天光照8h 的公鸡无显著差异[67]。Shi 等[68]对比16、18、20 和22 周光照刺激对白来航和北京油鸡种公鸡性成熟和精液品质的影响,发现不同组性成熟时间显著差异,但成年期精液品质和受精率无显著差异。在光波长方面的研究发现红光可以促进睾丸增重[69]。在同等能量水平下,长波长的光比短波长的光所含的光子更多,这可能是长波长的红光对繁殖性状的刺激作用较短波长的蓝光和绿光更显著的主要原因[70]。
3 小结
不同的品种品系因遗传背景导致精液品质和繁殖性能存在一定差异,在鉴定基因组和表观遗传调控元件基础上挖掘可以用于分子选育的有效标记至关重要。在营养调控技术方面,饲料营养成分或功能性添加剂对精液品质的影响主要是通过改善精子细胞膜功能和抗氧化能力来实现,有必要的更进一步研究营养元素或抗氧化添加剂等在不同品种和不同周龄群体的最适添加量。此外,种公鸡育雏育成期的养殖方式和环境管理对体成熟、性成熟和繁殖性能的维持都具有重要作用,不容忽视。
