秋季额木尔河江鳕的摄食生态
2024-01-09邵慧丽张泽鹏金洪宇迟淼邢悦李慎辉李雷
邵慧丽,张泽鹏,金洪宇,迟淼,邢悦,李慎辉,李雷
(1.中国水产科学研究院黑龙江水产研究所,农业农村部黑龙江流域渔业资源环境科学观测实验站,黑龙江 哈尔滨 150070;2.国家渔业资源环境抚远观测实验站,黑龙江 哈尔滨 150070;3.大连海洋大学水产与生命学院,辽宁 大连 116023)
额木尔河又作“额穆尔河”是黑龙江上游右岸支流,发源于伊勒呼里山北麓,由南向北集老槽河、大林河、古莲河等支流,横贯黑龙江省大兴安岭地区漠河市全境,在兴安镇大河西村附近注入黑龙江。额木尔河全长469 km,河宽20~150 m,水深2~5 m,流域面积16 280 km2,是黑龙江上游第二大支流。江鳕(Lota lota),地方名山鲶鱼、山怀子,属鳕形目(Gadiformes)鳕科(Gadidae),是鳕科鱼类中唯一的淡水种类,肉质鲜美、营养价值高、生长快以及适应能力强[1],广泛分布于45°N 以北的欧亚及北美的内陆水域和海湾处,是高纬度水域珍贵经济鱼类;在我国仅分布于额尔齐斯河、黑龙江水系及鸭绿江上游[2,3]。江鳕是典型的冷水性鱼类,在冰下进行生殖洄游和水温近0 ℃时产卵[4]。江鳕抗冻蛋白属III 型抗冻蛋白[5],是防寒抗冻的特种生物[4]。江鳕喜栖居于水质清澈的沙底或有水草生长的水域[6]。
近年来,环境污染、过度捕捞和繁殖场破坏等因素导致江鳕自然资源量锐减[7,8]。摄食行为是鱼类获取物质和能量的基本生命特征,是个体生存繁衍和种群增长的基础[9]。探究江鳕的摄食习性是保护江鳕的首要任务。目前,我国有关江鳕的研究最早出于施白南和高岫[10],此后陆续研究了不同水域江鳕的生长、生殖和食性,以及人工繁殖、分子遗传和病害等[1,6,11,12],对黑龙江支流额木尔河江鳕的摄食生态尚未见到相关研究。本文通过研究额木尔河江鳕的食物组成和摄食强度,及其随体长和空间的变化特征,以期为合理利用及保护江鳕资源提供科学依据和理论基础。
1 材料与方法
1.1 样本采集
2022 年10 月于额木尔河漠河市西林吉镇段、兴安镇段及支流大林河、老漕河和龙河(图1),用网目为1 cm×1 cm、2 cm×2 cm 和3 cm×3 cm 的定置刺网和网目4.0 mm 的地笼,单次放置时间为3 h 左右。采集样品,共获148 尾江鳕。
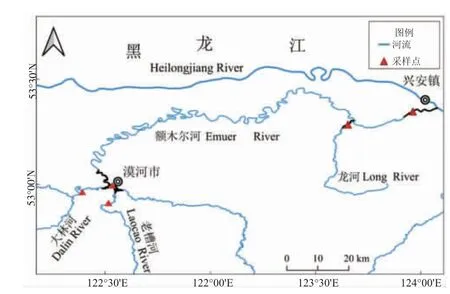
图1 额木尔河江鳕采样点Fig.1 Sampling sites of Lota lota in Emuer River
测定鱼类样本体长、全长、体质量等常规生物学指标,长度精确到0.1 mm,体质量精确到0.01 g。采集到的江鳕样本体长范围为107.0~372.0 mm,体质量范围为8.91~372.36 g。样本解剖后,取胃含物,储存在10%福尔马林溶液中,待胃含物的鉴定。
1.2 胃含物分析
将保存的样本带回实验室,取出胃含物,用NSZ-405 双筒解剖镜观察。在解剖镜下识别和计数胃含物中饵料生物的种类,根据形态特征加以鉴定到最低分类单元,用精度为0.000 1 g 的电子天平称重。称重前用滤纸吸干饵料生物表面水分。种类鉴定参照《黑龙江鱼类》[13]、《东北地区淡水鱼类》[14]和《水生昆虫》[15]。
1.3 数据处理
江鳕样本采集区域分为干流漠河市西林吉镇段(51 尾)、干流兴安镇段(65 尾)、支流大林河段(12 尾)、老漕河(0 尾)及龙河段(20 尾)5 个地点。由于老漕河未采集到样本,在数据分析中除去了此地点。
计算饵料生物数量百分比(N)、质量百分比(W)和出现频率(F)[16],进而采用相对重要性指数(IRI)[17]评价饵料生物的重要性。饱满指数和空胃率是反映鱼类摄食强度的重要指标[18,19],本研究利用胃饱满指数和空胃率评估江鳕摄食强度随体长和空间的变化。利用Primer 6.0 软件对数据进行聚类分析,所用指数为Bray-Curtis 相似性系数[20]。利用SPSS 26.0软件分析空胃率变化,使用卡方检验分析平均饱满指数变化。
计算公式如下:
其中,计算食物糜的数量百分比时,每尾鱼按数量为1 计算,以便于统计食物糜的质量百分比和出现频率等其他指标。
2 结果与分析
2.1 食物组成
江鳕的食物组成包括鱼类、水生昆虫、甲壳类、两栖类、圆口纲和其他等6 大类群。水生昆虫相对重要性指数(IRI=7 098.81)最高,是最重要的食物类群,其次是鱼类(IRI=5 480.44),甲壳类最低(IRI=0.09)。在水生昆虫中,网襀科(Perlodidae sp.,1 090.14)相对重要性指数最高;鱼类中,最高的是真(Phoxinus linnaeus,894.77);甲壳类中,仅有长臂虾科(Palaemonidae sp.,0.09)。按照质量百分比(W),鱼类是最大的类群(76.89%),其次是两栖类(8.25%)和水生昆虫(7.05%)。在鱼类中,真最多,其质量百分比为37.43%;在两栖类中,仅有林蛙(Rana sp.),其质量百分比为8.25%;在水生昆虫中,网襀科的质量百分比最高,为2.87%。按照数量百分比(N),水生昆虫是最重要的食物类群(80.32%),其次是鱼类(12.79%)。水生昆虫中,网襀科数量百分比最高(24.20%);鱼类中,真的数量百分比最高(4.13%)。按照出现频率(F),水生昆虫的出现频率最高(81.25%),其次是鱼类(61.11%),甲壳类和两栖类最低(均为0.69%)。水生昆虫中,网襀科的出现频率最高(40.28%);鱼类中,真(21.53%)最高(表1)。
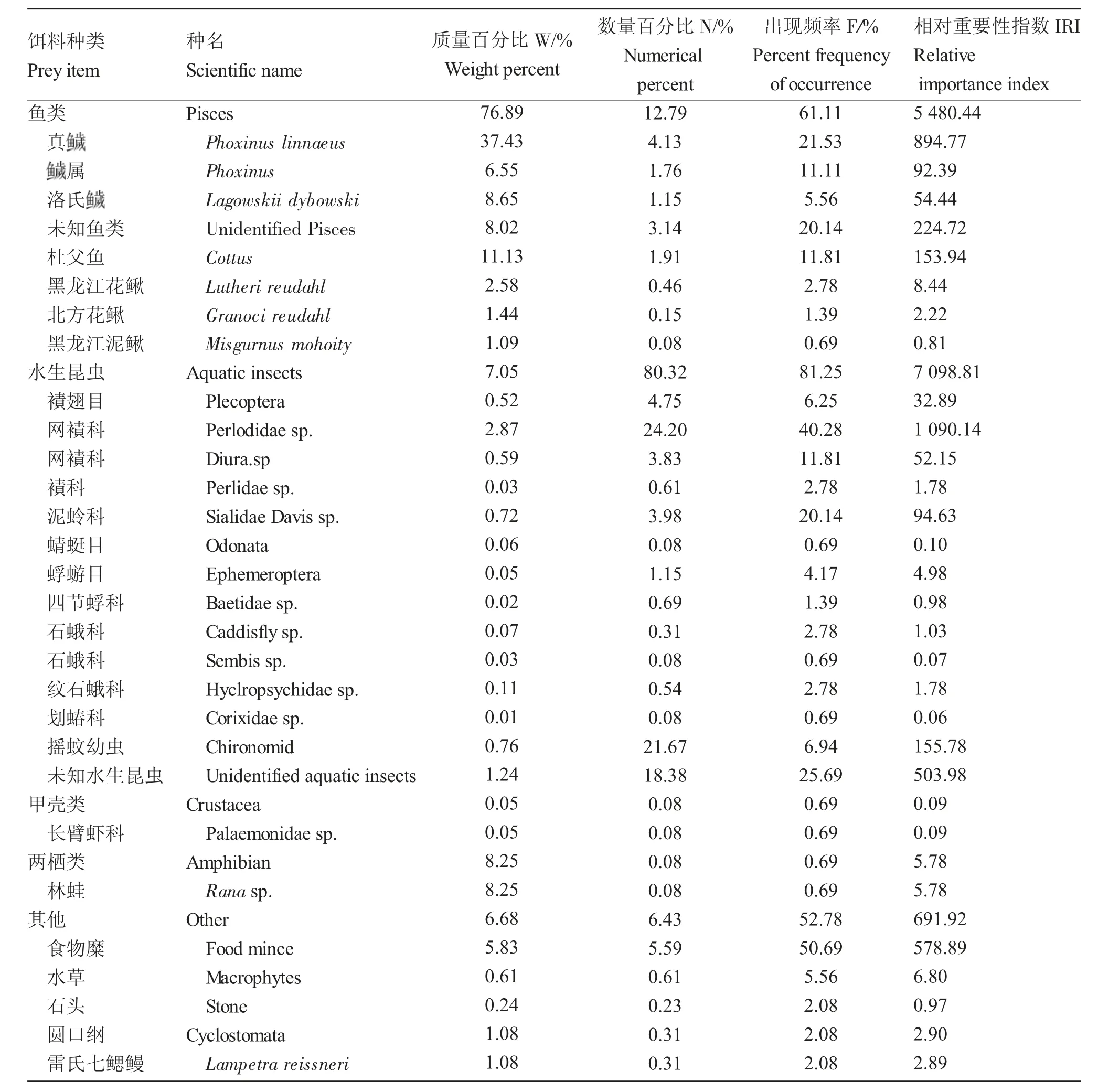
表1 额木尔河流域江鳕的食物组成Tab.1 The food composition of Lota lota in the Emuer River
2.2 食性随空间的变化
2.2.1 食物组成随空间的变化
将江鳕样本按照4 个采样区域进行分类,对比不同空间的食物组成差异,由于小于100.0 mm 和大于200.0 mm 的样本数量较少,以及支流大林河段样本数量太少,在统计分析前将此部分除去。
本研究中,100.0~200.0 mm 体长范围内江鳕的食物组成主要分为5 个类群,鱼类、水生昆虫、甲壳类、圆口纲和其他。由表2 可知,从相对重要性指数来看,漠河市西林吉镇段和兴安镇段均为水生昆虫最高,支流龙河段鱼类最高;从饵料的质量百分比来看,三个地点均为鱼类占比最高,水生昆虫次之;从数量百分比来看,均为水生昆虫最高,鱼类次之;从出现频率来看,均为水生昆虫最高,其次为鱼类。依据聚类分析的结果,组间相似度均大于85%(图2),表明额木尔河段江鳕的食物组成在空间分布上未出现明显的食性转换。尽管聚类分析结果显示江鳕的食物组恒未出现明显的食性转换,然而江鳕的食物组成随空间变化仍存在一定的差异,其中漠河市西林吉镇段和兴安镇段食物组成更相似。

表2 额木尔河流域江鳕食物组成的空间变化Tab.2 The Spatial variation diet composition of Lota lota in the Emuer River
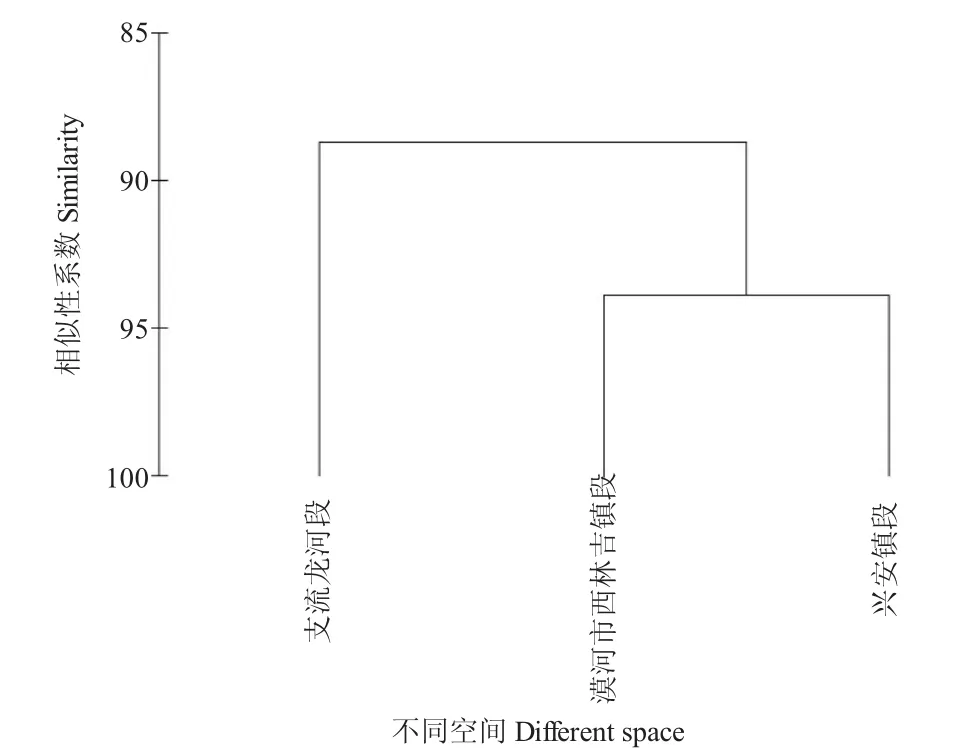
图2 额木尔河流域江鳕食物组成的聚类分析Fig.2 Cluster analysis of the dietary composition of Lota lota in the Emuer River
2.2.2 摄食强度随空间的变化
经SPSS 检验,本研究中不同空间的江鳕平均胃饱满指数(F=1.179,P>0.05)和空胃率(X2=0.754,P>0.05)均未呈现显著差异,即摄食强度在不同空间未出现显著差异。支流龙河段平均胃饱满指数最高(1.02%),兴安镇段次之(0.65%),漠河市西林吉镇段最低(0.51%);相反,漠河市西林吉镇段的空胃率最高(4.65%),兴安镇段次之(3.57%),支流龙河段的空胃率最低(0%)(图3)。

图3 江鳕的空胃率和平均胃饱满指数Fig.3 Percent of empty stomachs and mean stomach fullness index of Lota lota
2.3 兴安镇段江鳕的食性随体长变化
2.3.1 兴安镇段江鳕的食物组成
通过对比不同空间江鳕的摄食强度,未出现显著性差异(P>0.05),由于兴安镇段江鳕样品量最多,因此选择兴安镇段的江鳕研究其摄食随不同体长的变化特征。结果显示,兴安镇段江鳕摄食食物类群包括鱼类、水生昆虫、甲壳类、圆口纲、两栖类和其他。从相对重要性指数来看,水生昆虫是最重要的食物类群(IRI=6 873.67),其次是鱼类(IRI=4 746.57)。在水生昆虫中,网襀科最高(IRI=643.56);鱼类中,最高的是真(IRI=706.67)。按照质量百分比,鱼类是最大的类群(68.15%),其次是两栖类(17.58%)。在鱼类中,真是最重要的,其质量百分比最高(36.31%);在两栖类中,仅为林蛙(17.58%)。按照数量百分比,水生昆虫是最重要的食物类群(79.50%),其次为鱼类(12.67%)。水生昆虫中,摇蚊幼虫数量百分比最高(Chironomid)(33.00%);鱼类中,真的数量百分比最高(4.17%)。按照出现频率,水生昆虫的最高(80.95%),其次是其他(60.32%)。水生昆虫中,网襀科的出现频率最高(34.92%);其他中,食物糜出现频率最高(57.14%)(表3)。
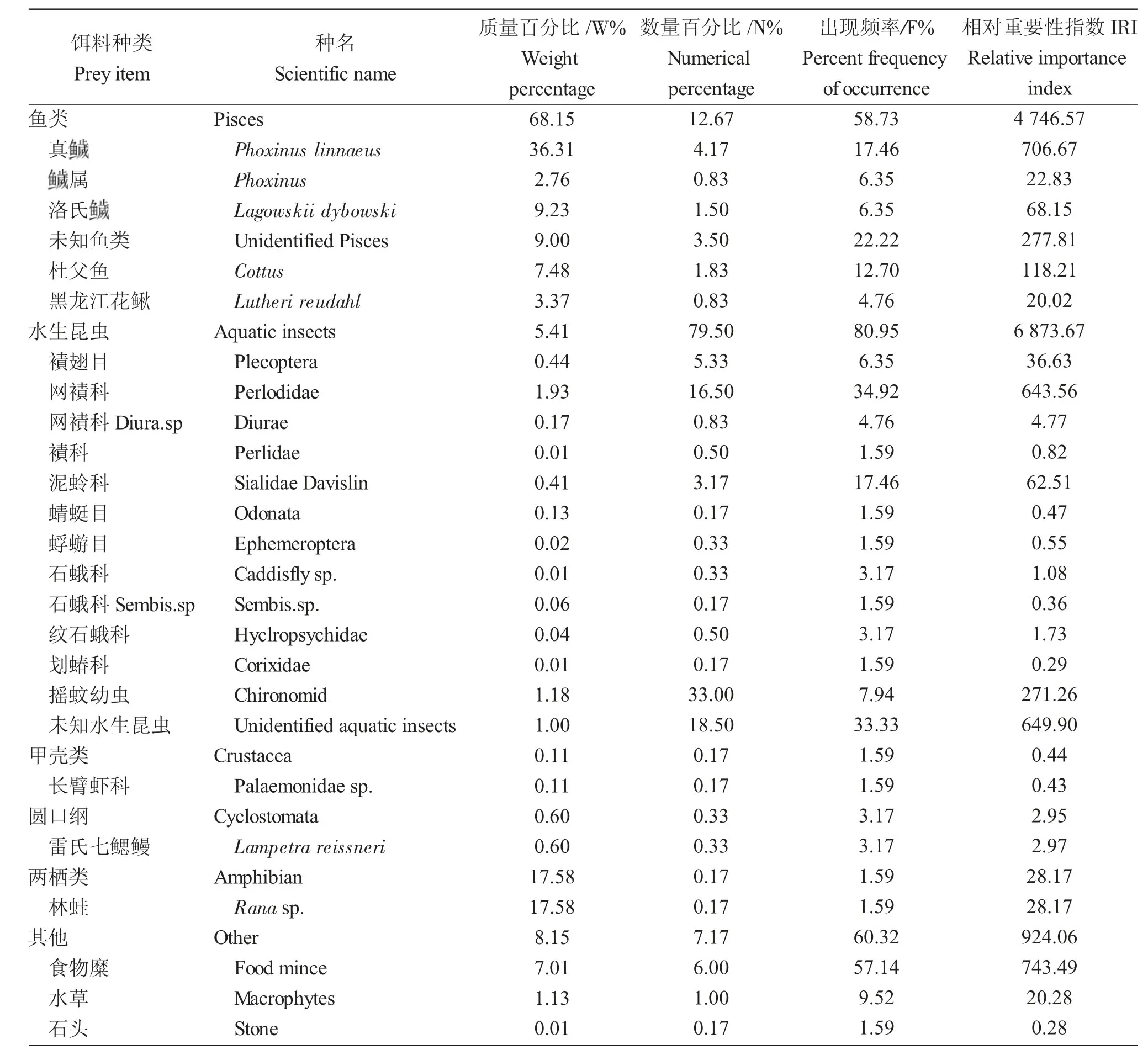
表3 额木尔河兴安镇段江鳕食物组成Tab.3 The diet composition of Lota lota in Xingan Town of the Emuer River
2.3.2 食物组成随体长的变化
由图4 可知,江鳕食物的质量百分比随体长的变化存在差异。体长小于200.0 mm 的江鳕主要以鱼类和水生昆虫为食,前者是最重要的食物类群;在100.0~149.9 mm 和150.0~199.9 mm 体长组,鱼类的质量百分比分别为69.35%和83.75%,水生昆虫的质量百分比分别为14.08%和7.96%;体长为100.0~199.9 mm 时,随着体长的增加,鱼类的质量百分比逐渐增加,水生昆虫的质量百分比逐渐降低。当体长大于或等于200.0 mm 时,江鳕食物类群中开始出现两栖类;江鳕的体长大于或等于200.0 mm 时,鱼类的质量百分比降低,为49.02%,两栖类的质量百分比增加,为43.31%,水生昆虫的质量百分比减少,仅为0.12%。对不同体长组江鳕食物组成的聚类分析显示,3 个体长组分为2 组,其中一组为100.0~199.9 mm(A 组),其平均相似性系数为85%;另一组为≥200.0 mm(B 组)。A 和B 组之间的平均差异性系数为57%。A 组江鳕主要摄食鱼类和水生昆虫,B 组主要摄食鱼类和两栖类,水生昆虫鲜有摄食(图5)。

图4 兴安镇段江鳕饵料种类质量百分比变化Fig.4 Variation of percentage of food item weight of Lota lota in Xingan Town

图5 额木尔河流域江鳕食物组成的聚类分析Fig.5 Cluster analysis of the food item composition of Lota lota in the Emuer River
2.3.3 摄食强度随体长的变化
单因素方差分析和X2检验发现,江鳕的平均饱满指数(P>0.05)和空胃率(P>0.05)均存在不显著的体长变化,各体长组空胃率随体长的增加,空胃率降低,不同体长组间江鳕空胃率的范围在0%~5%之间,体长大于或等于200.0 mm 时,空胃率最低(0%),体长为100.0~149.9 mm 时,空胃率最高(5%);相反,不同体长组间江鳕的平均饱满指数随体长的增加逐渐增加,100.0~149.9 mm 体长组最低(0.37±0.34)%,200.0 mm 以上体长组平均饱满指数最高(1.45±2.19)%(图6)。
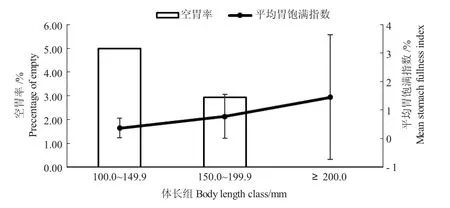
图6 兴安镇段江鳕的空胃率和平均胃饱满指数Fig.6 Percent of empty stomach and mean stomach fullness index of Lota lota in Xingan Town
3 讨论
3.1 食物组成
本文利用传统方法分析了额木尔河江鳕的食物组成,发现江鳕食物组成包括鱼类、水生昆虫、甲壳类、两栖类、圆口纲和其他等6 大类群。杨树勋等[21]研究发现,牡丹江上游江鳕以鱼类、水生昆虫和底栖动物为食;孟令博等[11]在乌苏里江虎头江段江鳕的食物中仅发现鱼类和甲壳类饵料生物;阿达可白克·可尔江等[1]研究发现,可可托海水库的江鳕以鱼类和水生昆虫及软体动物为食;任慕莲等研究表明,额尔齐斯河的江鳕食物主要为鱼类,其次为水生昆虫[12]。由于江鳕喜栖居于水质清澈的沙底或有水草生长的水域[6],本文江鳕摄食的极少量水草和石头,很可能为江鳕摄食其他鱼类或底栖生物时误食。综上所述,江鳕属于肉食性鱼类,不同水域中的江鳕均以鱼类和水生昆虫等为主要食物,是其固有属性,对于在不同水域中食物产生的差异,可能是不同水域中饵料生物的种类或数量发生变化,导致江鳕的食物类群随之改变[22-24]。对于在不同水域中食物差异,则是由食物的可得性引起,体现了江鳕食物的可塑性[1]。
3.2 食物组成和摄食强度随空间的变化
鱼类的食性和自然水域中饵料生物丰度不同有关,鱼类食性往往因时间、地点及饵料生物丰富度不同而变化[25]。在整个采样水域,江鳕主要摄食鱼类和水生昆虫,但不同空间之间存在差异,其原因可能是采样的地点差异[26]。如需揭示饵料生物分布的差异,后续将进一步对采样水域的饵料分布调查研究;同时,采样水域的环境存在不同程度的种间竞争,这也可能是差异产生的原因[27]。从摄食强度来看,三个不同空间的平均胃饱满指数和空胃率未出现明显的差异,很可能是因为研究样本的体长均为100.0~200.0 mm,江鳕的摄食强度差异不显著[28]。
3.3 兴安镇段江鳕食性随体长变化
兴安镇段江鳕在体长为200.0 mm 时食性转换(相似性系数>60%)。随着体长的增加,鱼类和水生昆虫的质量百分比从83.42%增加到91.7%,在体长组大于200.0 mm 时,水生昆虫减少,两栖类增加,鱼类和两栖类的质量百分比合计为92.32%。可能是随着鱼类体长的增长,鱼类摄食器官逐渐完善,捕食能力有所提高,食性类型也会发生一定的变化[29,30]。同时,随着食性的转变,在体长增长后摄食更多个体较大的饵料生物,保证在获取更多食物能量的同时消耗更少的捕食成本。该现象符合Gerking[31]提出的“最佳摄食理论”;随摄食器官和捕食能力的变化,一方面加强了捕食较大个体饵料生物的能力,另一方面也缓解了种内对食物资源的激烈竞争,以满足不同体长的个体对饵料的需求[32]。
鱼类摄食强度随体长增加而变化是普遍的现象[33,34]。本研究表明,在体长较小时,胃饱满指数较低,随着体长的增长,兴安镇段江鳕的胃饱满指数增加,空胃率降低。空胃率在200.0 mm 以上最低(0%),平均饱满指数最高。综合空胃率与胃饱满指数,江鳕的摄食强度随着个体发育逐渐增加,这可能与生长发育的能量储备有关,随着个体增长,江鳕的摄食能力不断提高,以满足其快速的生长和性腺发育[35]。
本文研究表明,江鳕主要摄食鱼类、水生昆虫和甲壳类,随着体长的增加,捕食的饵料生物种类增多。为了更好地保护额木尔河江鳕的资源,合理开发利用,建议做到以下几点:(1)加强自然水域中野生江鳕种质资源及栖息环境的保护;(2)加强江鳕的饵料生物(如小型鱼类)的保护,禁止破坏性的非法渔具渔法(如电捕鱼等),为江鳕提供充足的饵料生物;(3)深入开展科学研究,进一步研究额木尔河江鳕的饵料生物随季节的变化,开展江鳕繁殖生物学、关键栖息地分布及其生境特征的研究。
