巴西橡胶树ACA/ECA基因家族鉴定及分析
2024-01-08杨晶晶方永军张鸿韬龙翔宇秦云霞阳江华肖小虎
杨晶晶 方永军 张鸿韬 龙翔宇 秦云霞 阳江华 肖小虎

关键词:巴西橡胶树;Ca2+-ATPase(ACA/ECA);基因家族;生物学功能;表达分析
钙离子(Ca2+)作为植物细胞的第二信使几乎参与了植物生长发育的各个方面,包括对生物和非生物胁迫的反应[1]。植物细胞内Ca2+信号的传递主要是通过改变细胞内外钙离子浓度实现的,而Ca2+浓度的变化则主要由膜上的跨膜转运蛋白进行控制[2]。植物细胞的Ca2+转运主要由钙离子通道、钙离子反向转运蛋白和钙离子泵[3],Ca2+-ATPase属于P-型ATP酶离子超家族的P2分支,根据其蛋白质氨基酸序列和生化特征方面的差异分为2个亚家族(P2A和P2B),也分别称为内质网型Ca2+-ATPase(ER-typecalciumATPase,ECA,P2A)和自抑制的质膜(PM)型Ca2+-ATPase(autoinhibitedcalciumATPase,ACA,P2B)[4]。在植物细胞中,定位于质膜和内质网中的P型Ca2+-ATP酶被认为在调节细胞质Ca2+浓度方面起重要作用[5],同时P型Ca2+-ATP酶在植物生长发育的各个方面都是不可或缺的,包括花粉管生长、细胞程序性死亡、根部尖端生长,以及生物和非生物胁迫应答(如低温、高温、盐胁迫、干旱和渗透胁迫等)、共生等方面[3]。目前已经在拟南芥[6]、水稻[7]、白菜[8]、茄科植物[9]、苜蓿[10]等植物中鉴定得到ACA/ECA基因家族多个成员,并对其在植物中的功能进行了研究。研究表明,在拟南芥中,ACA8基因功能的缺失能显著提高拟南芥对零下低温的抗冻性[11];在低钙环境下,ECA1表达受到干扰的转基因株系耐受低Ca2+(0.2mmol/L)或高Mn2+(0.5mmol/L)胁迫的能力降低[12]。水稻ACA/ECA家族由15個成员组成,其中有3个属于内质网膜型钙离子泵,12个属于自抑制型钙离子泵,盐、干旱和ABA胁迫可以显著激活OsACA6的表达,但热胁迫激活作用相对较弱[13]。ACA/ECA家族在大白菜不同组织中表达模式不同,5个成员编码蛋白定位于细胞膜上,其中Bra002762、Bra035649、Bra031701与低温和盐胁迫响应有关,而Bra003276和Bra024117与自交不亲和性相关[5],大白菜缺少ECA基因会导致新叶生长缓慢,叶片枯萎黄化[14]。在茄科植物中,低温和盐胁迫处理后,SLyECA1基因表达丰度较高,且处理前后表达趋势变化明显,说明SLyACA1基因可能是提高植株抗性的关键基因[9]。野生大豆Ca2+ATPase基因GsACA1在苜蓿中超量表达,能够显著提高转基因苜蓿的耐盐碱性[10]。以上研究表明,ACA/ECA基因家族成员在多种植物中均发挥了重要作用。
巴西橡胶树是大戟科橡胶树属乔木,其产生的胶乳是天然橡胶生成的主要原料。天然橡胶(naturalrubber)是一种以聚异戊二烯为主要成分的天然高分子化合物,与合成橡胶相比天然橡胶性能更加优良,在很多方面具有不可替代性,可用于生产飞机轮胎、手套等数万种橡胶制品,在军事、医疗、运输等行业起着非常重要的作用[15],中国是目前为止世界上最大的天然橡胶消费国,年消费量占全世界总消费量的40%以上,但自给率不足为15%[16]。理论预测,以及生产上出现的高产橡胶树单株都显示出巴西橡胶树还具有巨大的产胶潜能[17-18]。目前已知的2500余种产胶植物中橡胶树表现出优异的产胶能力,推测与橡胶树中高丰度表达的橡胶延伸因子/小橡胶粒子蛋白(REF/SRPP)基因家族发生扩增有关[19]。已有研究表明,REF/SRPP基因[20]、HbNN2基因[21]、蔗糖转运蛋白HbSUT3[22]对橡胶树产胶具有重要作用。生产上,割胶和乙烯刺激都能显著提高胶乳产量,表明激素和伤害信号的传递与橡胶生物合成关系密切。钙离子是植物细胞内重要的信号传递因子,在植物应对生物和非生物胁迫方面具有重要功能,而目前关于钙离子泵ACA/ECA家族在橡胶树中的研究却未见报道。本研究以已发表植物ACA/ECA蛋白序列为探针从橡胶树转录组和基因组中鉴定出橡胶树ACA/ECA家族的全体成员,并从理化性质、系统进化和表达模式等方面进行了全面系统的分析。研究结果将为进一步研究ACA/ECA基因在橡胶树乳管钙离子信号的传递,以及橡胶树生长发育和逆境胁迫应答等方面的功能奠定基础。
1材料与方法
1.1材料
供试材料均来自于海南省儋州市中国热带农业科学院试验基地。用于表达分析的不同组织材料(叶片、树皮、胶乳、种子、雄花、雌花等6种组织)均来自于正常开割2年的热研7-33-97橡胶树。由于生产上开割树多为嫁接苗,根部并非热研7-33-97品种,因而本研究选用热研7-33-97组培苗的根作为根材料。为了分析乙烯刺激对基因表达的影响,对正常开割2年的热研7-33-97橡胶树(未涂抹过乙烯利,割胶频率为3d1刀)进行乙烯利刺激试验,提前0、3、12、24h将1.5%的乙烯利涂抹在橡胶树割面,然后在同一时间点采集胶乳样品。为了进一步研究割胶对ACA/ECA基因表达的影响,选取相邻2个林段,PR107品系和热研8-79的未开割橡胶树,同时进行3d1刀割胶,分别在第1、3、5、7、9刀采集胶乳样品。利用试剂盒提取以上样品的RNA,送往北京百迈克公司进行二代illumina转录组测序。
1.2方法
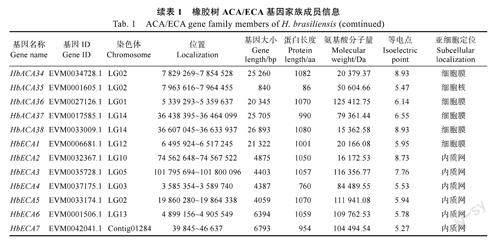
1.2.1橡胶树ACA/ECA家族成员鉴定及理化性质分析根据文献[6]的方法,从NCBI(https://www.ncbi.nlm.nih.gov)数据库中下载模式植物拟南芥的ACA/ECA蛋白序列,以拟南芥蛋白序列作为探针,通过本地服务器进行blastp比对(参数e值为1e-4),得到候选橡胶树ACA/ECA家族成员。利用在线软件CD-Search(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)对候选ACA/ECA家族成员所包含的结构域进行进一步鉴定分析,得到橡胶树ACA/ECA家族成员。利用在线软件Expasy(https://web.expasy.org/protparam/)和Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)对橡胶树ACA/ECA基因家族成员的氨基酸分子量、蛋白长度、理论等电点以及亚细胞定位进行分析。
1.2.2构建系统进化树以及基因结构图在NCBI网站通过blastp比对检索并下载水稻、杨树和木薯的ACA/ECA蛋白序列,使用多序列比對软件在线mafft(https://mafft.cbrc.jp/alignment/software/)对拟南芥、水稻、杨树、橡胶树和木薯5种植物的蛋白序列进行比对,使用MEGA6.0[23]软件采用邻接法构建系统进化树,参数bootstrap设置为1000,利用在线软件Figtree(http://tree.bio.ed.ac.uk/software/figtree/)对进化树进行调整;利用基因组序列和基因组注释GFF3文件对橡胶树ACA/ECA基因家族进行基因结构分析,通过GSDS(http://gsds.gao-lab.org/)[24]在线软件结合系统进化树绘制出橡胶树ACA/ECA基因结构图。
1.2.3橡胶树ACA/ECA家族成员染色体定位分析利用Tbtools[25]软件,根据橡胶树GFF3文件相关信息绘制出橡胶树ACA/ECA基因家族的染色体定位图;利用MCscanX软件对橡胶树基因组进行共线性分析,绘制染色体之间的共线性连线[26]。
1.2.4橡胶树ACA/ECA家族成员表达模式分析利用本实验室转录组数据,对ACA/ECA家族成员在不同组织中、乙烯利处理不同时间点以及割胶处理条件下的表达进行分析[27]。将测序得到的转录组数据上传到本地服务器,利用RSEM[28]软件包进行表达分析得到每个基因的FPKM值,利用Tbtools软件中的HeatMap小程序绘制热图。
2结果与分析
2.1橡胶树ACA/ECA家族成员鉴定及理化性质分析
利用已发表模式植物拟南芥的ACA/ECA蛋白序列[6],通过BLAST比对筛选橡胶树ACA/ECA基因家族候选成员,并利用在线软件CD-Search对候选成员的蛋白结构域进行进一步鉴定,最终鉴定得到45个ACA/ECA基因家族成员,其中包含38个ACA和7个ECA亚家族成员,分别命名为HbACA1~HbACA38和HbECA1~HbECA7。橡胶树ACA/ECA基因编码的蛋白分子量在10048.04Da(HbACA26)~125412.75Da(HbACA36)之间,理论等电点在5.18(HbACA18)~9.21(HbACA26)之间,45个成员中30个成员等电点均小于7,说明大多属于酸性蛋白。亚细胞定位结果显示,橡胶树ACA/ECA基因家族成员大多定位于细胞膜(表1)。
2.2系统进化树的构建及基因结构分析
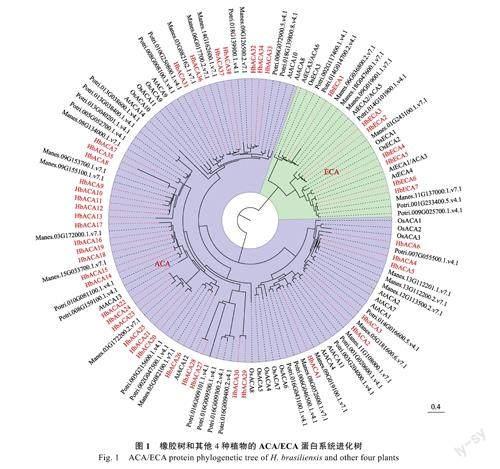
为了比较拟南芥(Arabidopsisthaliana,At)、木薯(Manihotesculenta,Me)、杨树(Populustrichocarpa,Pt)、水稻(Oryzasativa,Os)4种植物和橡胶树(Heveabrasiliensis,Hb)ACA/ECA蛋白在进化上的相互关系,利用mafft软件对5种植物蛋白序列进行多序列比对,并用MEGA6.0软件构建系统进化树,然后用Figtree软件对进化树进行修饰。从进化树可以看出(图1),ACA/ECA基因家族成员分别聚于ACA和ECA两个不同分支,橡胶树与木薯的ACA/ECA成员聚在一起,这与系统进化上木薯与橡胶树的亲缘关系较近是一致的;其中HbACA13、HbACA12、HbACA11、HbACA10、HbACA9、HbACA8和HbACA35聚集在同一个小分支,并且这7个成员均位于LGO2号染色体上,推测这7个成员是通过串联复制进化而来。
利用基因组GFF3注释文件对橡胶树ACA/ECA基因家族成员进行基因结构分析,并通过GSDS软件结合进化树绘制内含子-外显子结构图(图2)。在橡胶树45个ACA/ECA基因家族成员中,35个成员含有内含子,包括26个ACA和7个ECA成员,橡胶树ACA/ECA家族成员内含子数量在0~33之间,存在明显差异。同一分支上的成员在内含子数目和位置方面相对一致,除HbACA12、HbACA24、HbACA20、HbACA22、ACA25、HbACA15和HbACA16等12个成员无内含子外,其他成员均存在1~33个内含子;部分成员出现了较大数目的内含子,如HbACA31、HbACA34、HbACA38和HbECA1的内含子数目在30~33之间。
2.3橡胶树ACA/ECA基因家族成员染色体定位分析
从45个ACA/ECA基因家族染色体定位图可看出(图3),45个成员分布在14条染色体上,LG02和LG09号染色体上分布有最多的成员,均为11个,LG16染色体上有4个成员,LG08染色体分布有3个成员,LG07、LG03、LG12、LG05、LG14染色体上均有2个成员,LG06、LG01、LG10、LG13、LG11染色体上分布最少,均为1个成员,另外,contig上也有1个成员。进一步分析发现,LG02和LG09号染色体上的成员大部分成簇分布,LG08、LG12、LG14和LG16四条染色体上有成员成对分布,这些成员可能是通过串联重复复制而来,表明这些ACA/ECA家族成员蛋白序列具有较高的相似性,可能具有类似的生物学功能。蓝色线连接表明成员之间具有共线性,在进化过程中通过染色体加倍或大片段复制形成。
2.4橡胶树ACA/ECA基因家族表达分析
利用本实验室转录组数据对45个橡胶树ACA/ECA基因家族成员在不同组织(胶乳、树皮、叶片、种子、根、雌花和雄花)、乙烯利和割胶处理下的胶乳的表达情况进行分析并绘制热图。从橡胶树ACA/ECA基因家族成员在不同组织中的表达可以看出(图4),HbACA1、HbACA3、HbACA36、HbECA1、HbECA6和HbECA7功能较为保守,在各组织中均有表达;HbACA2在胶乳中特异高表达,而在其他组织中低表达或不表达,表现出组织特异性。另外,除HbACA2外,HbACA36、HbACA38和HbECA7四个成员在胶乳中的表达丰度也比较高,其中HbACA36在胶乳表达丰度最高;在叶片中,HbECA6、HbECA7、HbACA1、HbACA31和HbACA36表达丰度相对较高,其中HbACA31表达丰度最高;在树皮中,HbECA6、HbECA7、HbACA1、HbACA31、HbACA36、HbACA26和HbACA3表达丰度较高。HbECA7在叶片、树皮和根等多种组织中都有相对较高的表达,说明该成员可能在橡胶树钙离子运输方面具有重要功能。
乙烯利刺激是橡胶生产中常用的一种增产手段,为了进一步研究橡胶树ACA/ECA基因在乙烯利刺激后胶乳中的表达,本课题组在乙烯利处理后的4个不同时间点(0、3、12、24h)对ACA/ECA基因家族成员在胶乳中的表达进行了分析(图5)。经乙烯利处理后HbACA2、HbACA36、HbACA38、HbECA7四个成员表达呈上升趋势,其中HbACA36在乙烯利处理24h后呈现高丰度表达,并且在处理前后相对于其他成员都处于较高表达水平,这和之前不同组织表达分析的结果一致。HbACA3在乙烯利处理后上调表达,HbECA5在胶乳中低表达或不表达但经乙烯利刺激24h后上调表达。结合染色体定位图来看,HbACA36和HbACA38有较高同源性,但在表达上却存在差异,HbACA36表达丰度更高,并且乙烯利刺激后表达变化也更明显,推测HbACA36在乳管钙离子运输方面起主要作用。研究结果表明,部分成员乙烯利刺激后在胶乳中的表达呈明显的上升趋势,推测这些成员可能在乳管胶乳再生方面发挥重要作用。
在橡胶生产中发现,割胶能够促进橡胶树产胶。为了进一步分析割胶对橡胶树ACA/ECA基因家族成员表达的影响,选用2种不同产量水平的未开割橡胶树品种进行割胶试验,包括相对低产的PR107和高产品种热研8-79;同时对2个品种的已开割橡胶树进行割胶试验,通过转录组测序对基因的表达进行分析。由图6可以看出,在割胶过程中ACA/ECA基因家族成员在PR107和热研8-79中的表达趋势整体一致,如HbECA6、HbECA7、HbACA1、HbACA2、HbACA3、HbACA36、HbACA37和HbACA38在2个品种的割胶过程中均有表达。其中HbACA36在PR107和热研8-79割胶过程中均处于较高的表达水平,这与前面不同组织以及乙烯利处理的试验结果一致。但是HbACA36在热研8-79割胶过程的表达表现出明显的上升趋势(第1、3、5、7、9刀的FPKM值分别为206.19、221.96、281.45、353.36、355.59),而HbACA36在PR107中的表达则表现出先下降后上升再下降的表达趋势(第1、3、5、7、9刀的FPKM值分别为212.89、160.28、209.92、205.15、159.48),HbACA36在热研8-79中的整体表达水平高于PR107,这可能与热研8-79的高产具有一定的相关性。此外,HbACA18和HbACA19经割胶处理后仅在热研8-79中表达,再次说明橡胶树ACA/ECA基因家族成员在2个品种中的表达存在一定差异。
3讨论
Ca2+-ATPase(ACA/ECA)在植物逆境胁迫应答和生长发育等方面起重要作用[28],目前,在拟南芥[6]、大白菜[8]、茄科植物[9]等多种植物中鉴定得到ACA/ECA家族成员,并对部分家族成员的功能进行了系统分析,而关于ACA/ECA基因在橡胶树中的功能的研究还未见报道。天然橡胶是重要的战略物质,在工业和国防等方面发挥着重要作用,橡胶树作为生产天然橡胶的唯一来源,其种植和生产一直备受关注[29],随着经济的发展,天然橡胶的消耗也不断增长,出现供不应求[30],因而提高橡胶产量显得尤为重要,钙离子信号在乙烯刺激和割胶过程中的传递对增加胶乳产量促使乳管胶乳再生至关重要,因此探明ACA/ECA基因家族在橡胶树中的功能有重要研究意义。本研究鉴定得到橡胶树中45个ACA/ECA基因家族成员,包括38个ACA成员,7个ECA成员,其中ACA成员明显多于ECA成员,这与其他植物的研究结果一致。从系统进化树来看有2个明显分支,在2个分支中橡胶树与木薯ACA/ECA聚在一起,这与橡胶树和木薯在系统进化上有较近的亲缘关系是一致的。从染色体定位图看,有成员成对或者成簇存在,其中2号和9号染色体上的部分成员聚在一起,结合进化树可以看出,相对于杨树和木薯等其他植物,橡胶树ACA/ECA成员在部分分支上出现了扩张,并且扩张主要由串联复制产生的。在表达方面,部分ACA/ECA成员在不同组织中的表达表现出组织特异性,如:HbACA2在胶乳中表达丰度较高,HbACA12在种子中表达丰度较高,而在其他组织中表达丰度很低或不表达;在胶乳中,HbACA2、HbACA36、HbACA38和HbECA7四个成员表达丰度相对较高,其中HbACA36在胶乳中表达丰度最高,并且明显高于其他3个成员,推测HbACA36在乳管钙离子转运方面具有重要作用。乙烯利刺激和割胶是橡胶树生产中最主要的2种增产手段,乙烯利处理后,HbACA36在胶乳中表达呈现明显的上升趋势,在高产品种热研8-79割胶过程中HbACA36的表达也表现出明显的上升趋势,表明HbACA36在橡膠树乳管中可能具有重要功能并且和橡胶树的产量具有一定的相关性。综上表明,HbACA36在橡胶树乳管的信号传递方面起重要作用,进而影响橡胶树乳管中的橡胶生物合成。本研究相关结果为进一步研究ACA/ECA基因的生物学功能,及其在乙烯刺激橡胶树增产过程中的重要作用奠定了基础。
