萨能奶山羊初乳和常乳的代谢组学分析
2024-01-03康佳欣廖敏和商佳琦任皓威卢玺丞
康佳欣,李 萌,廖敏和,3,商佳琦,3,任皓威,3,卢玺丞,刘 宁,3,
(1.齐齐哈尔大学食品与生物工程学院,黑龙江 齐齐哈尔 161006;2.东北农业大学食品学院,乳品科学教育部重点实验室,黑龙江 哈尔滨 150030;3.哈尔滨腾凝科技有限公司,黑龙江 哈尔滨 150028)
萨能奶山羊是中国重要的奶山羊品种之一,具有产奶量高、奶质好、繁殖快和抗病能力强等特点[1]。萨能奶山羊乳是萨能奶山羊从乳腺分泌的一种白色或微黄色液体,含有丰富的蛋白质、脂肪、碳水化合物、维生素、矿物质等营养素及其他代谢物。相比牛乳,山羊乳以低致敏性[2]和易消化吸收[3]的特点,深受人们的重视。
山羊乳成分易受到泌乳期、地理区域和饲料等因素的影响。目前对于泌乳期的分析较为普遍,已有研究表明随着泌乳时间的延长,山羊乳中总干物质、蛋白质和脂肪含量均呈下降趋势[4-5],这些研究集中于分析不同泌乳期间常规乳成分的差异,但对于山羊乳中小分子代谢物的研究鲜有报道,小分子代谢物的变化通常采用代谢组学方法进行分析。
代谢组学是“组学”科学的一个新分支,研究特定有机体中代谢物组成和相关代谢通路分析的一门学科,主要技术手段包括核磁共振(nuclear magnetic resonance,NMR)、气相色谱-质谱(gas chromatography-mass spectrometry,GC-MS)联用及液相色谱-质谱联用技术[6-7]。代谢组学技术已经广泛用于分析不同泌乳期乳中的代谢物差异,Li Mohan等[8]采用气相色谱-飞行时间质谱技术比较不同泌乳期母乳的代谢物变化,共筛选出72 个差异性代谢物,与成熟乳相比,初乳中高丰度代谢物有17 个,丰度下调有55 个。Wu Junfang等[9]采用NMR技术对比不同泌乳期人乳的差异性代谢物,与泌乳9~24 d相比,泌乳31~87 d的乳糖和胆碱等物质相对含量升高,柠檬酸和磷酸胆碱等物质相对含量降低;Klein等[10]采用NMR和GC-MS技术对比泌乳早期和晚期牛乳中代谢物差异,共检测出44 个差异性代谢物,丙酮和β-羟基丁酸可作为泌乳早期潜在的标志性代谢物;Li Mohan等[11]采用超高效液相色谱-四极杆飞行时间质谱法分析驴初乳和驴常乳的代谢物差异,共识别出52 个差异性代谢物,与驴常乳相比,驴初乳中有44 个代谢物丰度上调,8 个代谢物丰度下调,这些研究证实了代谢组学可以用于区分不同泌乳期乳中的差异性代谢物。
因此,本研究采用超高效液相色谱-四极杆静电场轨道阱质谱(ultra-high performance liquid chromatographyquadrupole electrostatic field orbitrap mass spectrometry,UPLC-QE-orbitrap-MS)非靶向代谢组学方法,对萨能奶山羊初乳和常乳中的代谢物进行综合表征,进一步分析相关的代谢通路,筛选出它们的标志性代谢物。本研究全面了解萨能奶山羊初乳和常乳的小分子代谢物成分变化,结果将有助于羊乳营养的进一步研究,并为分析不同泌乳期的其他物种原料乳提供基础数据支持。
1 材料与方法
1.1 材料与试剂
萨能奶山羊初乳(产后3 d)和常乳(产后120 d)由黑龙江绿能生态牧业有限公司提供。
甲醇、乙腈、乙酸铵、氨水(均为色谱级)德国CNW公司;超纯水 香港屈臣氏集团有限公司。
1.2 仪器与设备
BSA124S-CW分析天平 德国Sartorius公司;Vanquish UPLC仪、Q Exactive HFX高分辨质谱系统、Heraeus Fresco17离心机 美国Thermo Fisher Scientific公司;PS-60AL超声仪 深圳市雷德邦电子有限公司。
1.3 方法
1.3.1 样品采集
随机选取16 只萨能奶山羊,初乳和常乳各采集8 个样品,每个样本人工采集40~45 mL初乳或常乳存放于50 mL无菌离心管中并作好标记。萨能奶山羊要求健康且体质量相近,胎次为1 胎,平均泌乳期250~270 d,喂养的饲料以苜蓿和青贮为主。在挤奶前用45~50 ℃的温水清洗乳房部位,消毒乳头,避免乳头表面的菌污染乳汁。所有采集的样品在6 h内经过冷链运输至实验室,放入超低温冰箱贮存备用。
1.3.2 样品预处理
将样品在4 ℃解冻,取50 μL样品至离心管中,加入400 μL提取液(甲醇和乙腈以体积比为1∶1混合,含同位素标记内标混合物),涡旋混匀30 s。随后在冰水浴中超声10 min,-40 ℃冰箱中孵育1 h,沉淀蛋白。然后在4 ℃、12 000 r/min离心15 min,取上清液过膜后转移至进样瓶中上机检测。所有样品另取等量上清液混合成质控样品上机检测。
1.3.3 检测条件[12]
UPLC 条件:色谱柱为UPLCBEH Amide(2.1 mm×100 mm,1.7 μm);流速为500 μL/min;柱温为30 ℃;样品盘温度为4 ℃;流动相A为水相(含25 mmol/L乙酸铵和25 mmol/L氨水),流动相B为乙腈;进样体积为3 μL。梯度洗脱程序见表1。

表1 梯度洗脱程序Table 1 Gradient elution program
MS条件:Q Exactive HFX质谱仪在采集软件(Xcalibur,Thermo)的控制下,进行质谱数据采集,以信息依赖采集模式获取全扫描MS谱。电喷雾电离源条件为鞘气体流量50 arb;辅助气体流量10 arb;毛细管温度320 ℃;全MS分辨率60 000,MS/MS分辨率7 500;碰撞能量10/30/60 eV;在正和负模式下离子喷雾电压分别为3 500 V和-3 200 V。
1.4 数据处理
原始数据经ProteoWizard软件转成mzXML格式后,使用自主编写的R程序包(内核为XCMS)进行峰识别、峰提取、峰对齐和积分等处理,然后与BiotreeDB(V2.1)二级质谱数据库匹配进行物质注释,算法打分的Cutoff值设为0.3。
2 结果与分析
2.1 UPLC-QE-orbitrap-MS图谱分析
采用UPLC-QE-orbitrap-MS法分别在正、负离子模式下对初乳和常乳进行信息采集,由图1可知,在正、负离子模式下初乳和常乳的总离子流图基线平稳,说明仪器的稳定性良好。出峰时间、响应强度以及出峰数量存在差异,但是不能直观获取到样品的小分子代谢物信息,所以需要对原始数据进一步处理。
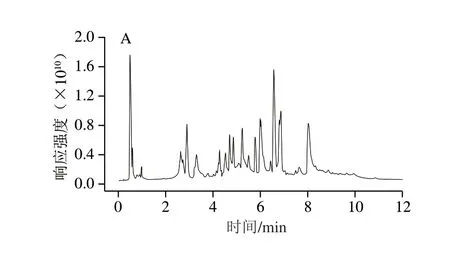
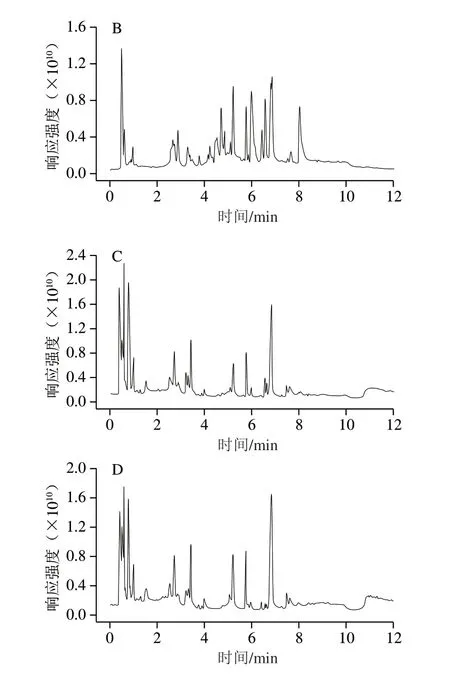
图1 初乳和常乳随机单个样品的总离子流图Fig.1 Total ion current chromatograms of random individual samples of colostrum and mature milk
2.2 主成分分析(principal component analysis,PCA)
为判别初乳和常乳样品组间差异,采用PCA方法进行多元统计分析。图2A、B分别为正负离子模式下初乳和常乳的PCA得分图,正离子模型参数为=0.581>0.5(为模型对X变量的解释性),负离子模型参数为=0.633>0.5,说明拟合性较好,模型较稳定。由图2可知,所有样品均在95%置信区间内,初乳大多数集中在置信区间上侧;常乳大部分集中在置信区间下侧,这说明初乳和常乳能够区分开,存在组间差异,但是两组组内差异较大,这可能是因为PCA是一种无监督的分析方法,不能忽略组内差异[13],因此需要采用一种有监督的方法进一步分析。
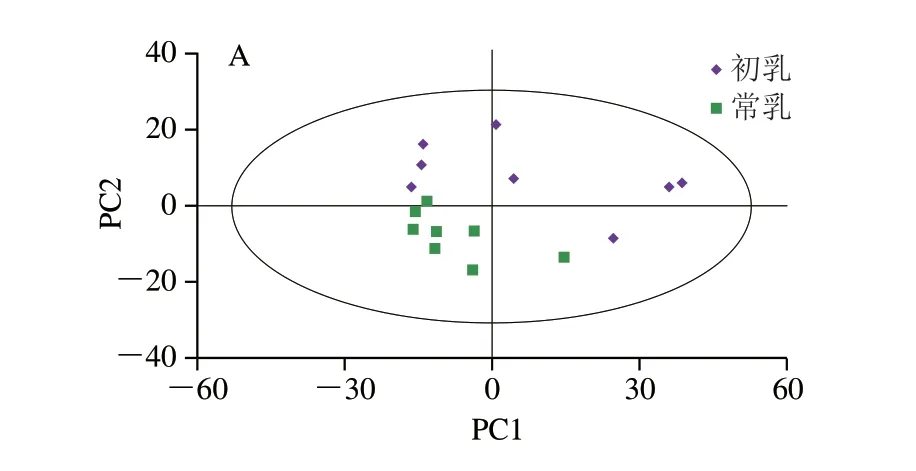
图2 初乳和常乳的PCA得分图Fig.2 PCA score plots of colostrum and mature milk
2.3 正交偏最小二乘判别分析(orthogonal partial least squares-discriminant analysis,OPLS-DA)
OPLS-DA是一种有监督的分析方法,这种模型可以减少组内差异、放大组间差异,排除无关因素对实验数据造成的影响[14-15],因此本实验又采用OPLS-DA模型对初乳和常乳样品进行分析。图3A、B分别为正负离子模式下初乳和常乳的OPLS-DA得分图,在正离子模式下=0.398,=0.94,Q2=0.648(为模型对Y变量的解释性,Q2为模型的可预测性);在负离子模式下=0.38,=0.959,Q2=0.739。这两个模型的和Q2都大于0.5,说明模型的稳定性较好,所有样品都处于95%置信区间内,初乳全部分布在置信区间左侧,常乳全部分布在置信区间右侧,初乳和常乳组间区分显著,OPLS-DA结果优于PCA。从分布形态上看,初乳样品较分散,常乳样品较集中,这可能是由于初乳中萨能奶山羊个体差异较大,导致组内差异较大。虽然萨能奶山羊个体差异会对初乳内代谢物产生一定影响,但是两组样品组间区分显著,这说明不同泌乳期的影响大于个体差异影响。图3C、D分别为正负离子模式下初乳和常乳的OPLS-DA置换检验图,通过置换检验判断OPLS-DA是否出现过度拟合。和Q2的回归线与横坐标相交小于0,说明模型可靠。置换检验随机模型的Q2值均小于原模型的Q2值,说明原模型有较好的稳定性,不存在过度拟合现象。
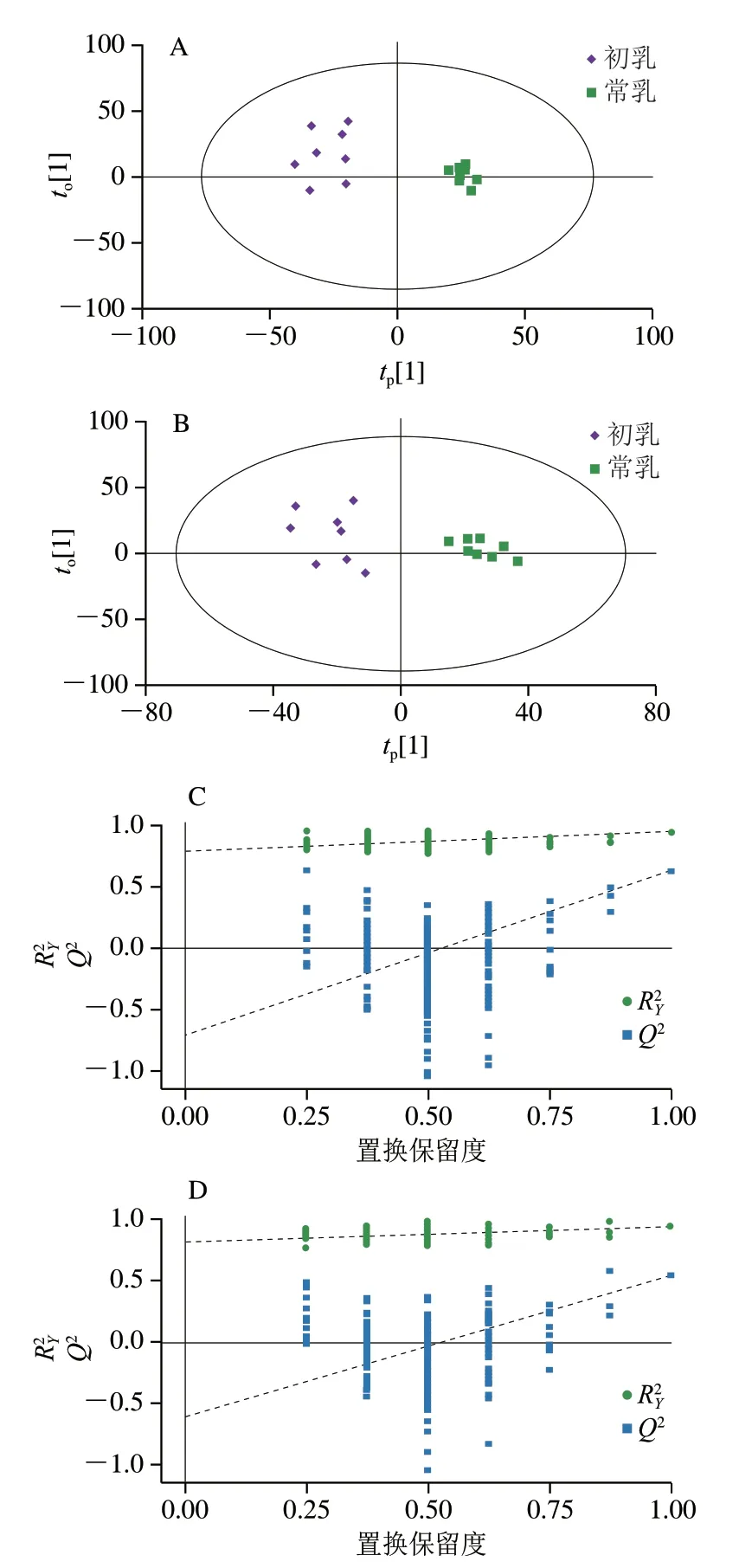
图3 初乳和常乳的OPLS-DA得分图和OPLS-DA置换检验图Fig.3 OPLS-DA score plot and OPLS-DA permutation test of colostrum and mature milk
2.4 差异性代谢物的筛选
筛选OPLS-DA模型中t检验的P<0.05,同时满足VIP>1的数据,与BiotreeDB(V2.1)二级质谱数据库进行匹配得到118 个差异性代谢物(表2、3),其中初乳中有62 个代谢物相对含量高于常乳,有56 个代谢物相对含量低于常乳。在这些差异性代谢物中脂质代谢物的种类较为丰富,包括磷脂酰胆碱(phosphatidylcholine,P C)、甘油二酯(diacylglycerol,D G)、鞘磷脂(sphing omyelin,S M)、磷脂酰乙酰胺(phosphatidylethanolamine,P E)、神经酰胺(ceramide,Cer)、磷脂酰肌醇(phosphatidylinositol,PI)。已有研究表明初乳和常乳中脂质的含量和种类不同[16-17],本研究证实了这一点。初乳中有9 个脂质代谢物分别为3 个PC、2 个DG、2 个SM、1 个PE和1 个Cer;常乳中有10 个脂质代谢物分别为5 个Cer、3 个PC、1 个DG、1 个PE和1 个PI,可以看出SM在初乳中相对含量高于常乳,PI在初乳中相对含量低于常乳,Liu Hongyan等[18]通过对比山羊初乳和常乳的脂质组成发现与常乳相比,初乳中SM的相对含量升高,PI的相对含量降低,与本研究结果一致。在初乳的脂质差异性代谢物中,PC(22:0/P-16:0)的差异倍数最大,在初乳中相对含量是常乳中的6.14 倍,它又称卵磷脂,研究表明在大脑中PC在乙酰化酶的作用下与乙酰辅酶A反应生成乙酰胆碱,乙酰胆碱具有强化大脑神经突触发育的功效[19],因此PC在初乳中相对含量升高可能与新生羊羔的大脑神经发育有关。
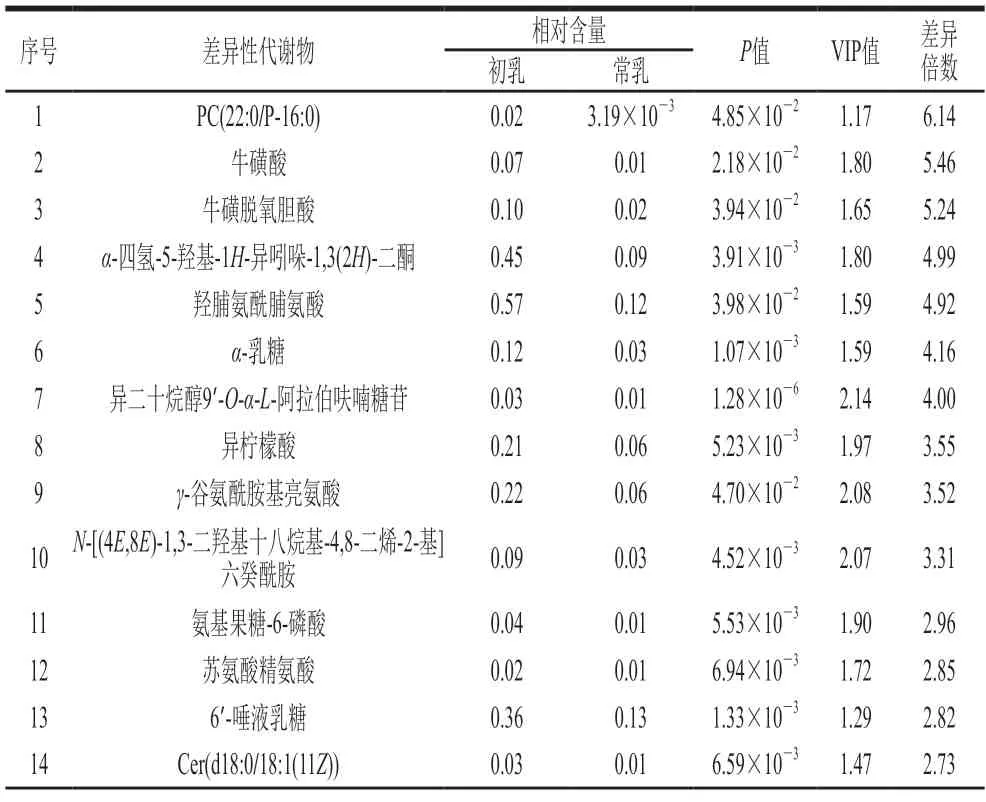
表2 正离子模式下初乳和常乳对比的差异性代谢物Table 2 Differential metabolites between colostrum and mature milk in positive ion mode
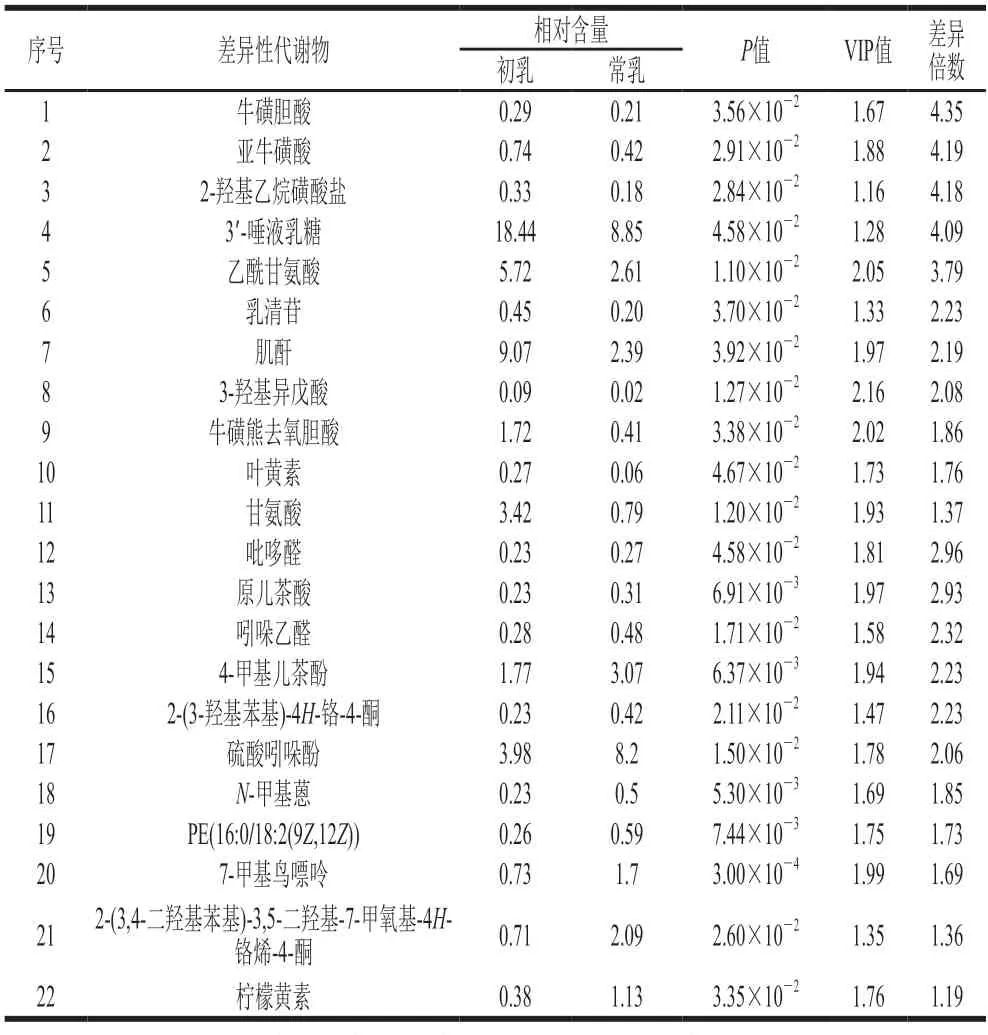
表3 负离子模式下初乳和常乳对比的差异性代谢物Table 3 Differential metabolites between colostrum and mature milk in negative ion mode
常乳中2′-岩藻糖基乳糖(2′-fucosyllactose,2′-FL)的相对含量高于初乳,是常乳中差异倍数最大的代谢物,它在常乳中相对含量是初乳的12.97 倍。2′-FL是人乳中最重要的低聚糖之一[20],具有增强婴儿免疫[21]、促进肠道菌群生长[22]等作用,一些商家已经把2′-FL应用于婴儿配方食品中使得成分和营养价值更接近人乳,这可能是羊乳更适合婴幼儿食用的原因之一。
初乳中存在一些相对含量高于常乳的二肽,如γ-谷氨酰胺基亮氨酸、苏氨酸精氨酸、天冬氨酸-脯氨酸和N-鸟氨酸-L-牛磺酸,它们在初乳中的相对含量是常乳的3.52、2.85、2.00、1.37 倍,在体内二肽可以参与蛋白质的合成,进一步被组织利用合成蛋白质[23]。初乳中的牛磺酸除了二肽形式外还有多种存在形式,如牛磺酸、牛磺脱氧胆酸、牛磺胆酸、亚牛磺酸、牛磺熊去氧胆酸,它们在初乳中相对含量是常乳的5.46、5.24、4.35、4.19、1.86 倍,其中牛磺酸是初乳中差异倍数排名第2的代谢物,它是一种含硫的半必需氨基酸,可以提高神经元的存活率、促进海马神经元的生长、具有调节下丘脑-垂体-肾上腺轴以及脑组织中miRNA表达的作用[24-26]。牛磺酸还可以与脱氧胆酸或胆酸结合生成牛磺脱氧胆酸或牛磺胆酸,后两者在羊羔体内起到促进脂质类物质吸收的作用[27-28]。由此可见,初乳可以为新生羊羔的生长发育提供充足的营养。
初乳中一些核苷类代谢物的相对含量高于常乳,如鸟苷、肌苷、N6-甲基腺苷,它们在初乳中相对含量是常乳的2.50、2.40、1.71 倍,前两者在与磷酸结合后合成鸟苷酸和肌苷酸,它们是核苷酸类化合物参与了生物体内几乎所有的生物化学反应,如多种单体核苷酸经过脱水缩合后合成DNA和RNA。此外,核苷酸也可以提高婴儿的免疫调节功能和记忆力[29]。
2.5 差异性代谢物的代谢通路分析
萨能奶山羊的泌乳过程是多个代谢通路共同调控的复杂过程,不能通过某一代谢物相对含量的高低进行整体判断,因此为更全面、更系统地了解萨能奶山羊初乳到常乳泌乳期间的生物学变化,需要进一步对差异性代谢物的代谢通路进行分析。通过与京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)中的代谢通路数据库进行匹配后进行富集分析和拓扑分析,筛选出与代谢物差异相关性最高的关键通路。初乳和常乳对比的关键代谢通路分析结果(表4)表明,共筛选出9 条代谢通路,1号为正负离子模式下共同检测出的通路,2~7号为正离子模式,8~9号为负离子模式,分别为牛磺酸和亚牛磺酸代谢、苯丙氨酸酪氨酸和色氨酸生物合成、柠檬酸循环、烟酸和烟酰胺代谢、淀粉和蔗糖代谢、半乳糖代谢、酪氨酸代谢、甘氨酸丝氨酸和苏氨酸代谢、一次性胆汁酸生物合成。为了更直观地看出这些代谢通路在富集分析中的富集显著情况以及拓扑分析中的影响程度,将上述9 条代谢通路以气泡图展示(图4A)。气泡图中每一个气泡代表一条代谢通路,气泡颜色表示该通路在富集分析中的P值,颜色越深P值越小,富集程度越显著;气泡大小表示拓扑分析的影响因子大小,气泡越大影响因子越大[30]。从图4A可以看出,牛磺酸和亚牛磺酸代谢的颜色最深且气泡最大,说明这条代谢通路富集程度最显著且影响因子最大,且命中代谢物数量最多,分别为牛磺酸、亚牛磺酸和牛磺胆酸,它们在初乳中的差异倍数较大,显著高于常乳中的相对含量(P<0.05),这可能是该通路富集程度最显著且影响因子最大的原因之一。已有研究表明牛磺酸的含量会随着泌乳时间的延长而逐渐降低[8,25],这与本研究的结果一致。实验还观察到半乳糖代谢气泡大小仅次于牛磺酸和亚牛磺酸代谢,说明这条代谢通路的影响因子较大,这可能是与该通路的命中代谢物差异倍数有关,初乳中α-乳糖的相对含量是常乳的4.16 倍,显著高于常乳中的相对含量(P<0.05),可能导致该通路影响因子较大[11,31-32]。
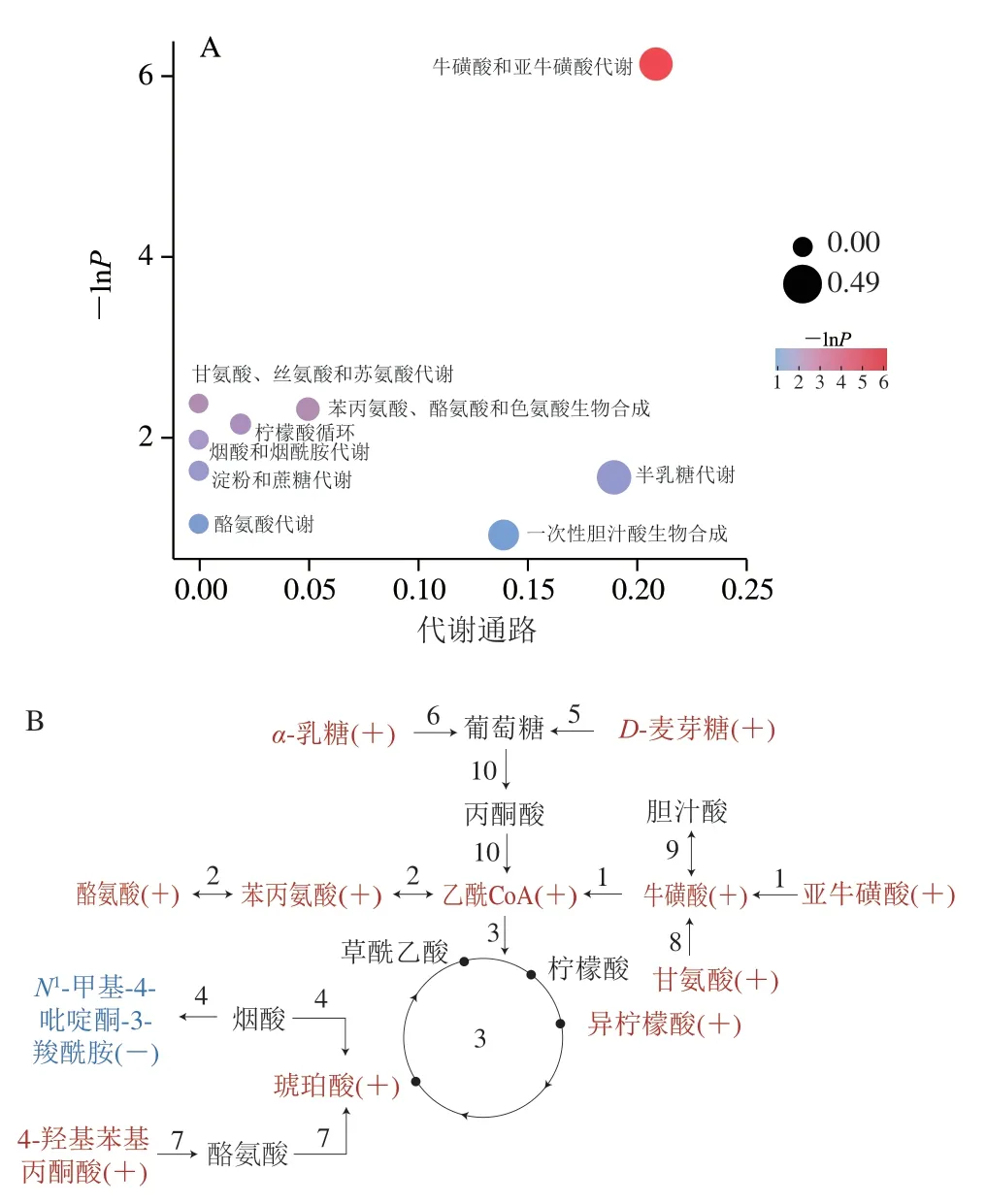
图4 初乳和常乳的代谢通路气泡图(A)和路径图(B)Fig.4 Bubble chart (A) and roadmap (B) of metabolic pathways in colostrum and mature milk
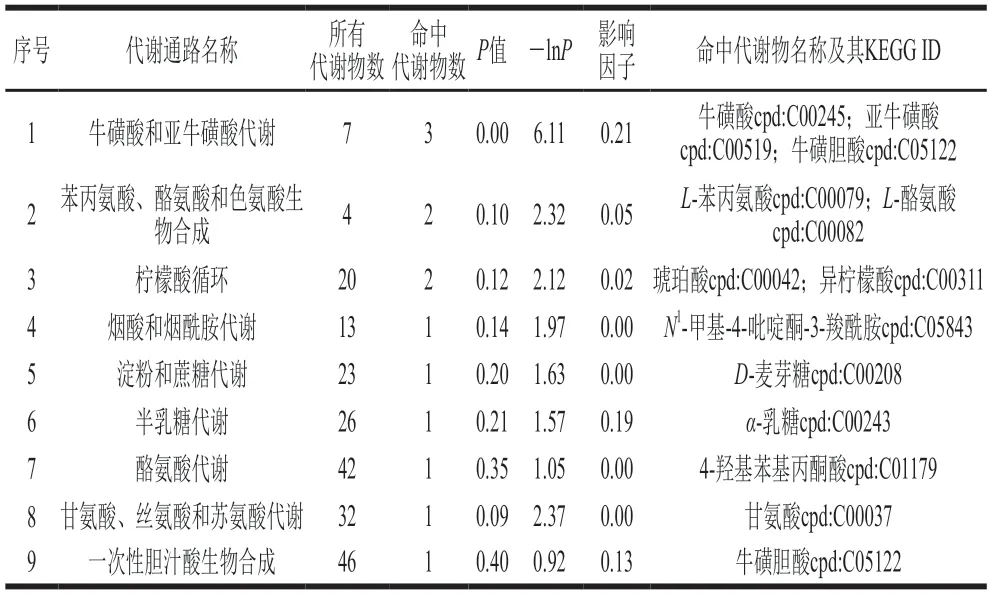
表4 初乳和常乳对比的关键代谢通路分析Table 4 Analysis of key metabolic pathways in colostrum and mature milk
为了更系统地了解泌乳期间的代谢通路之间的相关性,参考KEGG代谢物数据库中代谢通路图谱将这9 条关键代谢通路连接在一起(图4B)。图4B中红色表示该代谢物在初乳中相对含量高于常乳,蓝色表示该代谢物在初乳中相对含量低于常乳,黑色表示未匹配到的代谢物,由此可知初乳中相对含量高的代谢物为牛磺酸、亚牛磺酸、牛磺胆酸、L-苯丙氨酸、L-酪氨酸、琥珀酸、异柠檬酸、D-麦芽糖、α-乳糖、4-羟基苯基丙酮酸、甘氨酸,常乳中相对含量高的代谢物为N1-甲基-4-吡啶酮-3-羧酰胺,它们可作为这种萨能奶山羊初乳和常乳中潜在的标志性代谢物。从图4B可以看出,除N1-甲基-4-吡啶酮-3-羧酰胺外,其他代谢物都经过一系列的代谢通路后最终进入柠檬酸循环,如亚牛磺酸、牛磺胆酸、L-苯丙氨酸、L-酪氨酸、D-麦芽糖、α-乳糖转换为乙酰辅酶A进入柠檬酸循环,4-羟基苯基丙酮酸转换为琥珀酸进入柠檬酸循环,因此可以推测在泌乳初期柠檬酸循环起到桥梁作用,为新生羊羔健康生长提供充足的能量。在12 个命中代谢物中,仅有N1-甲基-4-吡啶酮-3-羧酰胺在初乳中的相对含量低于常乳,这可能是因为初乳中大部分烟酸生成琥珀酸,使得生成N1-甲基-4-吡啶酮-3-羧酰胺的相对含量降低。
3 结论
以萨能奶山羊初乳和常乳为研究对象,采用UPLCQE-orbitrap-MS的非靶向代谢组学方法,研究泌乳期对萨能奶山羊乳中代谢物的影响。结果表明:初乳中脂质代谢物、二肽类代谢物、核苷类代谢物、牛磺酸及其衍生物等62 个差异性代谢物的相对含量显著高于常乳(P<0.05),2′-FL等56 个差异性代谢物的相对含量显著低于常乳(P<0.05)。筛选出与差异性代谢物匹配最高的9 条关键通路,它们共同表征萨能奶山羊的泌乳过程的代谢物差异,这些代谢通路的命中差异性代谢物数量为12 个,初乳中相对含量高的代谢物为牛磺酸、亚牛磺酸、牛磺胆酸、L-苯丙氨酸、L-酪氨酸、琥珀酸、异柠檬酸、D-麦芽糖、α-乳糖、4-羟基苯基丙酮酸、甘氨酸,常乳中相对含量高的代谢物为N1-甲基-4-吡啶酮-3-羧酰胺,它们可作为萨能奶山羊初乳和常乳中潜在的标志性代谢物,在更多样本测定中加以验证。本研究结果有利于了解在不同泌乳期间萨能奶山羊乳中小分子营养成分的变化,为鉴别不同泌乳期的其他物种乳提供相应的参考。
