铜绿假单胞菌噬菌体的分离鉴定和全基因组序列分析
2024-01-02谢丹莹谢景昊黄若超李和平王月影
谢丹莹,谢景昊,黄若超,陆 鑫,杨 晨,郭 爽,李和平,王月影
(1.河南农业大学动物医学院农业农村部动物生化与营养重点实验室,河南 郑州 450046 ; 2. 河南省动物生长发育调控重点实验室,河南 郑州 450046)
铜绿假单胞菌(Pseudomonasaeruginosa,Pa)又称绿脓杆菌,广泛存在于土壤、水、空气、动物和人类呼吸道和肠道内。Pa是一种机会性人兽共患病的病原体,可引起家禽、家畜和宠物的各种急慢性感染,如烧伤感染、中耳炎、乳腺炎、子宫内膜炎、肺炎、尿路感染、肠炎、脑膜炎和囊性纤维化等,最终发展成致死性腹泻、菌血症和败血症等疾病[1-5],给养殖业造成巨大经济损失。Pa适宜在潮湿环境中生长,也是引起环境污染和严重医院感染的常见病原体[6]。Pa还是一种重要的食源性病原菌,主要毒力因子是外毒素A(Pseudomonas exotoxin A,PEA)、脂多糖(Lipopolysaccharides,LPS)、内毒素蛋白(Endotoxin protein,OEP)和蛋白分解酶等,可引起患者产生胃肠炎和神经症状[7-9]。全球范围内耐药铜绿假单胞菌的检出率介于10%~28%[10-12]。由耐药铜绿假单胞菌引起的感染性疾病的病死率高,治疗困难,给临床治疗带来了巨大挑战。铜绿假单胞菌感染的治疗方法主要为抗生素联合支持治疗,而抗生素长期大量使用可导致耐药菌株日益增多,增加了临床治疗铜绿假单胞菌感染的难度[13]。因此,寻找新型方式代替抗生素治疗已成为研究热点。
噬菌体是感染细菌的一种病毒,具有宿主特异性高、安全性高和成本低等优点,是替代抗生素和细菌耐药研究中的热点之一[13]。其中,铜绿假单胞菌噬菌体制剂研发已成为控制耐药铜绿假单胞菌感染性疾病的重要手段[14]。多项研究证明了噬菌体治疗耐药铜绿假单胞菌的可行性[15-17]。最新的研究表明,噬菌体能有效破坏铜绿假单胞菌的生物膜[18]。但是,噬菌体治疗耐药菌感染还面临一些局限,如宿主菌血清型特异性、噬菌体抗性等问题[19]。本试验利用铜绿假单胞菌临床分离株PA31为宿主菌,从水中分离鉴定了1株铜绿假单胞菌噬菌体,根据噬菌体分类和命名指南[20],将新分离的噬菌体命名为vB_PaeM_Ty,对其形态学、生化特性和基因组学方面进行分析,为进一步研究噬菌体治疗铜绿假单胞菌感染提供理论依据和资源储备。
1 材料与方法
1.1 试验菌株 铜绿假单胞菌临床分离株PA31由河南农业大学动物分子药理毒理学创新团队刘建华教授馈赠,分离自家兔的肝脏组织。本实验室保存的铜绿假单胞菌(PA31)、大肠杆菌(Escherichiacoli,No. HM68-1、39、34、33、HM79-2)、沙门菌(Salmonella,No. SJ249、JS1A、SJ16L、SJ43、S、SJ15、SQ)和粪肠球菌(Enterococcusfaecalis,No. 8a、n22、5b、22a、SJ-9)的临床分离株,用于噬菌体裂解谱分析。
1.3 主要仪器 透射电子显微镜(JEM-1400),日本电子株式会社产品;BIO RAD凝胶成像仪,上海伯乐生命医学产品有限公司产品;核酸电泳仪,上海天能科技有限公司产品。
1.4 样本处理 以铜绿假单胞菌临床分离株PA31为宿主菌。多位点采集河南农业大学校园湖水20份。将采集的湖水混合,取混合液和LB肉汤混合,再加入宿主菌液,置于摇床振荡培养过夜。冷冻离心后,取上清用0.22 μm滤膜过滤,得到初始噬菌体液。
1.5 噬菌体的分离纯化 采用点滴法鉴定噬菌体。取600 μL 1×1010CFU/mL宿主菌液(OD600 nm=0.4)与10 mL半固体LB琼脂培养基混匀,倾入LB琼脂培养基上,滴加10 μL初始噬菌体液,37 ℃过夜培养。若形成透亮、清晰的噬菌斑,证明成功分离出噬菌体;反之,则无。根据参考文献[21]报道的方法略作修改,采用双层琼脂平板法纯化噬菌体。将初始噬菌体液倍比稀释,分别取各梯度稀释液100 μL与600 μL宿主菌液混合于离心管中,倒入10 mL半固体LB琼脂培养基,混匀后倾入LB琼脂培养基上,过夜培养,直到形成透亮的噬菌斑。挑取单个噬菌斑于1 mL SM悬浮培养液中,5 000×g离心10 min,倍比稀释后同上采用双层琼脂平板法纯化5~7次,直到形成大小、形态一致的噬菌斑。根据最后一次纯化结果,按公式(1)计算噬菌体的效价。
效价(PFU/mL)=噬斑数×稀释倍数×10
(1)
1.6 噬菌体的裂解谱分析 以 PA31和实验室保存的Escherichiacoli、Salmonella、Enterococcusfaecalis临床分离株作为宿主菌,通过点滴法分析噬菌体的裂解谱。分别吸取400 μL菌液,加入10 mL半固体LB琼脂培养基,上下颠倒2~3次后迅速倒入到LB琼脂培养基,滴加8 μL噬菌体液,静置15 min,然后37 ℃过夜培养,观察是否形成噬菌斑。
1.7 噬菌体的透射电镜观察 根据参考文献[22]报道的方法浓缩噬菌体,取0.5 mL 1×1010CFU/mL宿主菌液(OD600 nm=0.4)和1 mL 1×1012PFU/mL的噬菌体液,放入摇床180 r/min振荡培养至液体清亮。10 000×g离心10 min,取上清用0.22 μm滤膜过滤。加入DNase Ⅰ、RNase A和NaCl混匀冰浴1 h。8 000×g离心10 min,取上清,加入体质比为10%的PEG 8000,4 ℃过夜培养。8 000×g 离心10 min,弃上清,加入SM悬浮培养液和等体积氯仿,5 000×g离心15 min,弃上清,得到的沉淀物即为浓缩后的噬菌体颗粒,送至河南中医药大学实验中心,用2%磷钨酸负染10 min,在透射电子显微镜下观察并拍照。
1.8 噬菌体生物学特性分析
问题3 一般地,一元二次方程ax2+bx+c=0(a≠0)的根与二次函数f(x)=ax2+bx+c(a≠0)的图象有何联系呢?
1.8.1 噬菌体最佳感染复数(Multiplicity of infection,MOI)测定 根据参考文献[23]报道的方法略作修改。取100 μL 1×1010CFU/mL宿主菌液(OD600 nm=0.4),将噬菌体效价稀释为108、109、1010、1011和1012PFU/mL,以MOI分别为0.01、0.1、1、10和100的比例将宿主菌液与噬菌体液混合,加入等体积的LB肉汤,恒温摇床振荡培养6 h,10 000×g 离心10 min,取上清用0.22 μm滤膜过滤,双层琼脂平板法测定噬菌体效价,以培养末期噬菌体效价最高的MOI为最佳MOI。
1.8.2 噬菌体的一步生长曲线绘制 将1×1010CFU/mL宿主菌液与1×1012PFU/mL噬菌体液按照最佳感染复数进行配比,室温孵育100 min,每隔10 min取样,采用双层琼脂平板法测定噬菌体效价,以效价为纵坐标,培养时间为横坐标绘制一步生长曲线,判断潜伏期和裂解期,并按公式(2)计算裂解量。
裂解量(PFU/mL)=裂解末期噬菌体效价÷初始宿主菌液浓度
(2)
1.8.3 温度对噬菌体稳定性的影响 将600 μL 1×1012PFU/mL噬菌体液置于不同温度(45、60、70和90 ℃)中水浴1 h,在60 min内每隔10 min取样,采用双层琼脂平板法测定噬菌体效价,绘制温度对噬菌体影响的曲线。
1.8.4 pH对噬菌体稳定性的影响 取100 μL 1×1012PFU/mL噬菌体液与不同pH(pH=2~12)的SM悬浮培养液混合,37 ℃恒温孵育1 h,采用双层琼脂平板法测定噬菌体效价,以pH为横坐标,噬菌体效价为纵坐标,绘制pH对噬菌体影响的曲线。
1.9 噬菌体基因组提取和核酸类型鉴定 按照病毒DNA提取试剂盒说明书进行操作,将0.5 mL噬菌体液经12 000×g离心5 min,弃掉沉淀,在上清中加入蛋白酶K、溶液V和无水乙醇,12 000×g离心2 min,弃掉上清液,沉淀加入漂洗液,12 000×g离心2 min,重复2次。最后沉淀加入洗脱液,静置5 min,离心得到噬菌体的基因组。取60 μL噬菌体的基因组,平均分成3份,分别加入DNase Ⅰ、RNase A和Mung Bean Nuclease各2 μL,37 ℃处理1 h,用0.1%琼脂糖凝胶电泳鉴定其核酸类型。
1.10 噬菌体的全基因组分析 将提取的噬菌体的全基因组送至派森诺生物科技有限公司进行全基因组测序。利用Illumina NovaSeq平台进行测序,测序数据通过A5-MiSeq(https://arxiv.org/abs/1401.5130)和SPAdes(http://cab.spbu.ru/files/release3.12.0/manual.html)进行拼接,通过GeneMarkS(http://topaz.gatech.edu/GeneMark/)预测基因组编码的蛋白质。Rfam数据库(http://rfam.xfam.org/)预测非编码RNA。采用Diamond软件(http://github.com/bbuchfink/diamond)完成蛋白质编码基因的序列比对,在NCBI数据库中用BLASTp对开放阅读框(Open reading frames,ORFs)进行功能注释,利用SnapGene 4.24软件进行全基因组分析。将噬菌体全基因序列和主要衣壳蛋白基因序列在NCBI进行对比,下载同源性较高的噬菌体基因组序列,用MEGA 11.0软件构建系统发育树。
2 结果
2.1 噬菌体的分离纯化 以 PA31为宿主菌,分离得到1株铜绿假单胞菌噬菌体,在双层琼脂平板上形成圆形、清晰的噬菌斑,直径为1~2 mm(图1),效价为1×1012PFU/mL。
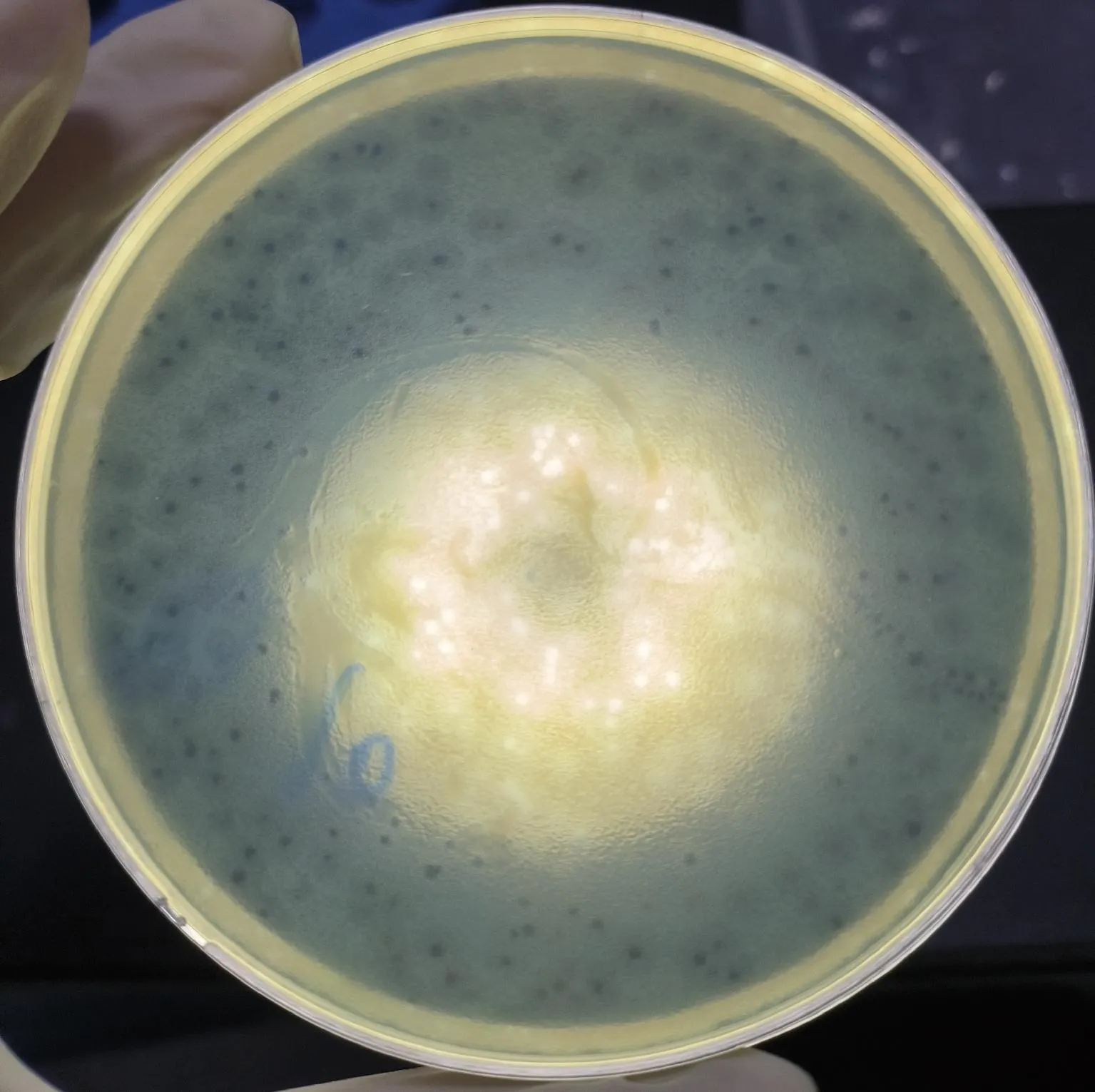
图1 噬菌体的噬菌斑观察Fig.1 Plaque observation of phages
2.2 噬菌体的裂解谱分析 噬菌体分离株对Pseudomonasaeruginosa、Escherichiacoli、Salmonella和Enterococcusfaecalis临床分离株的裂解结果显示,除宿主菌PA31和Escherichiacoli-39、Enterococcusfaecalis-8a菌株外,其他受试细菌平板上均未出现明显清晰的噬菌斑(表1),表明分离的噬菌体具有较强的宿主特异性。

表1 噬菌体分离株的裂解谱Table 1 Lysis spectrum of isolated phage
2.3 噬菌体的透射电镜观察 透射电子显微镜观察噬菌体的形态,结果显示,该噬菌体头部呈类矩形状,大小为50 nm×20 nm;尾部呈“尾鞘”样结构,长度在100 nm左右(图2),根据形态判断该噬菌体为有尾病毒目(Caudovirales)、肌尾病毒科(Myoviridae),根据噬菌体分类和命名指南[20],将该噬菌体命名为vB_PaeM_Ty。

图2 透射电镜观察噬菌体形态(150 000 ×)Fig.2 Observation of phage morphology by transmission electron microscope(150 000 ×)
2.4 vB_PaeM_Ty的最佳MOI 在MOI为1.0时,vB_PaeM_Ty效价最高,达到1.32×1011PFU/mL;MOI为0.01时,vB_PaeM_Ty效价最低,为1.00×109PFU/mL(表2)。因此,vB_PaeM_Ty的最佳MOI为1.0。
2.5 vB_PaeM_Ty的一步生长曲线 vB_PaeM_Ty的一步生长曲线见图3,vB_PaeM_Ty与宿主菌孵育20 min内,噬菌体效价变化不明显,之后随时间延长效价开始逐渐升高,60 min时效价达到高峰,此时噬菌体效价为1.42×1012PFU/mL,随后效价趋于稳定。结果表明,噬菌体的潜伏期为20 min,裂解期为40 min,根据公式计算裂解量约为142 PFU/mL。
2.6 温度对vB_PaeM_Ty稳定性的影响 由图4可知,不同温度条件下,随着时间的延长,vB_PaeM_Ty的效价均呈下降趋势,而且温度越高时vB_PaeM_Ty效价下降越快;在45、60和70 ℃静置60 min,vB_PaeM_Ty活性较稳定,90 ℃条件下随培养时间延长噬菌体效价在逐渐下降,60 min时效价下降至3.0×102PFU/mL。结果表明,vB_PaeM_Ty对温度的耐受能力较好。

图4 温度对vB_PaeM_Ty稳定性的影响Fig.4 Influence of temperature on vB_PaeM_Ty stability
2.7 pH对vB_Pae_Ty稳定性的影响 由图5可知,vB_PaeM_Ty在pH 3~12时效价较稳定;pH为7时,噬菌体效价最高,约为3.7×108PFU/mL,裂解能力最高;pH为2时,噬菌体vB_PaeM_Ty效价最低,约为1.0×105PFU/mL。
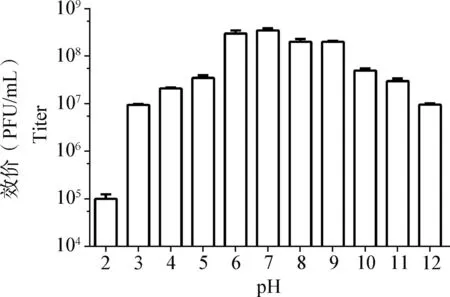
图5 pH对vB_PaeM_Ty稳定性的影响Fig.5 Influence of pH on vB_PaeM_Ty stability
2.8 vB_Pae_Ty的核酸类型和全基因组功能分析 琼脂糖凝胶电泳结果见图6,噬菌体vB_PaeM_Ty的基因组核酸为线性双链DNA(Double strand DNA,dsDNA)。vB_PaeM_Ty的基因组长50 051 bp,GC含量为47.23%,编码74个ORFs。噬菌体vB_PaeM_Ty全基因组核酸序列提交至GenBank,登录号为OQ555403。对vB_PaeM_Ty的全基因组进行非冗余蛋白数据库(Non-Redundant Protein Sequence Database,NR)注释后,得到71个相应的编码蛋白,基因的进化谱系:非同源组(Evolutionary genealogy of genes:non-supervised orthologous groups,eggNOG)注释后得到8个相似蛋白,京都基因和基因组数据库(Kyoto Encyclopedia of Genes and Genomes,KEGG)注释后得到6个相似蛋白,蛋白注释数据库(SwissProt)注释后得到6个相似蛋白,基因本体(Gene Ontology,GO)注释后得到11个相似蛋白。SnapGene 4.24软件绘制的全基因组分析结果见图7。通过BLASTp进行功能预测,有25个体现功能和结构的蛋白(表3),其他均为假定蛋白;其中结构蛋白有尾刺蛋白(Tail protein)和衣壳蛋白(Capsid protein)等,DNA相关蛋白有DNA结合蛋白(DNA binding protein)和DNA聚合酶(DNA polymerase)等,裂解细菌的蛋白有内溶素(Endolysin)和果胶裂解酶(Pectate lyase)等。
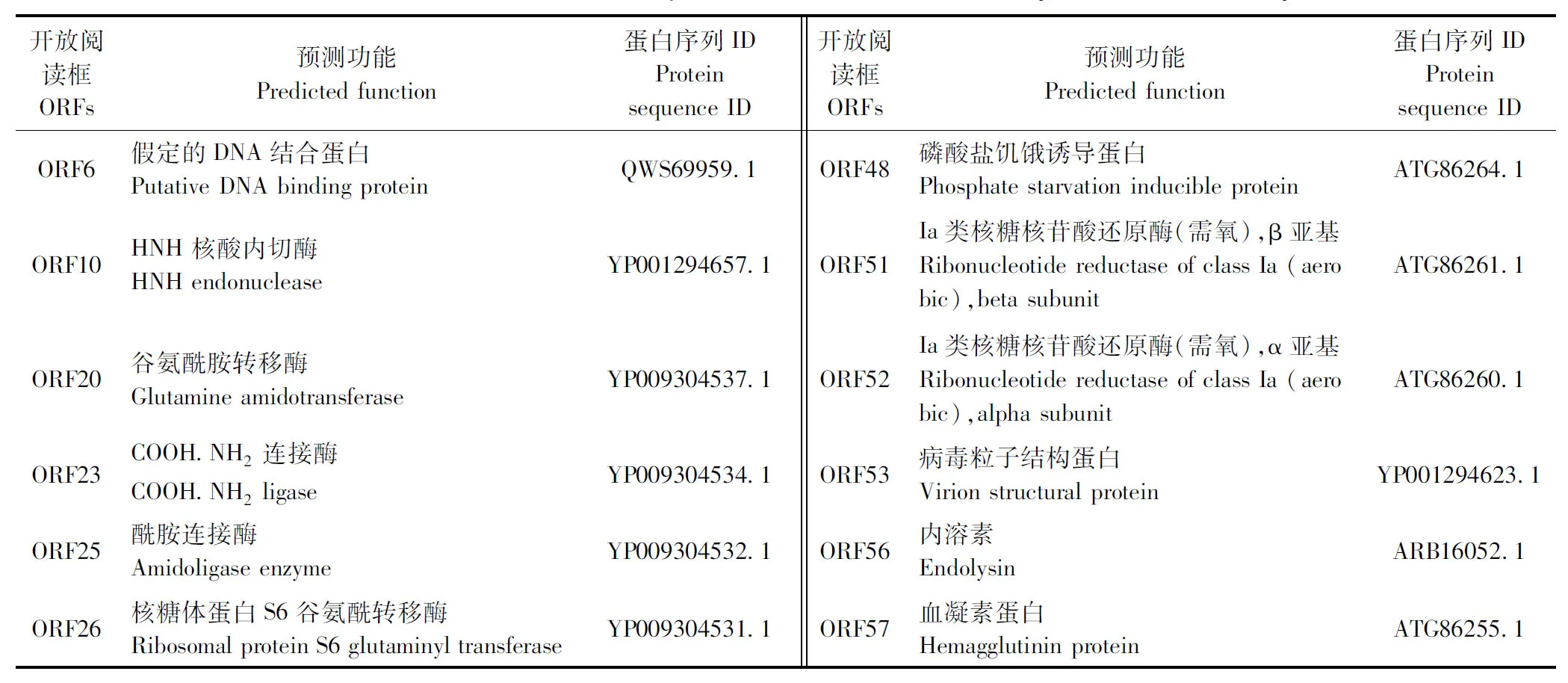
表3 vB_PaeM_Ty的全基因组ORFs预测和功能分析Table 3 Genome-wide ORFs prediction and functional analysis of vB_PaeM_Ty
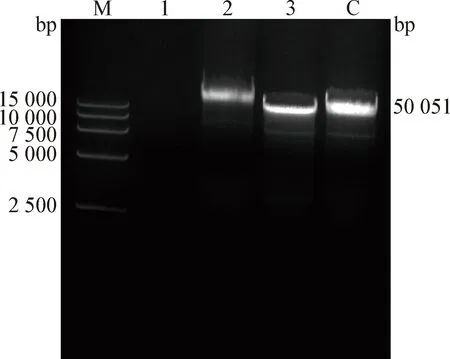
图6 vB_PaeM_Ty基因组的凝胶电泳分析Fig.6 Gel electrophoresis analysis of vB_PaeM_Ty genomeM:DL15 000 DNA Marker; 1:DNase I酶切; 2:RNase A酶切;3:Mung Bean Nuclease酶切; C:阴性对照(全基因组)M:DL15 000 DNA Marker; 1:Digestion with DNase I; 2:Digestion with RNase A; 3:Digestion with Mung Bean Nuclease; C:Negative control (whole genome)

图7 vB_PaeM_Ty全基因组结构示意图Fig.7 Schematic representation of whole genome structure of vB_PaeM_Ty
2.9 vB_PaeM_Ty的系统发育树分析 通过BLASTn比对全基因组序列,vB_PaeM_Ty与假单胞菌噬菌体K4的同源性最高,有92%的覆盖率和97.36%的同源性;与假单胞菌噬菌体O4全基因序列有91%的覆盖率和97.58%的同源性。根据全基因组和主要衣壳蛋白(ORF67)序列构建系统发育树,发现vB_PaeM_Ty与噬菌体K4、O4和IME180有较为密切的同源关系(图8)。除假单胞菌属噬菌体外,vB_PaeM_Ty与克雷伯菌噬菌体(Klebsiellaphage)、弧菌噬菌体(Vibriophage)、埃文氏噬菌体(Erwiniaphage)和柠檬酸杆菌噬菌体(Citrobacterphage)在同一分支(图9)。

图8 噬菌体vB_PaeM_Ty全基因组系统发育树Fig.8 Complete genome phylogenetic tree of phage vB_PaeM_Ty●:噬菌体vB_PaeM_Ty的全基因组序列●:Complete genome sequence of phage vB_PaeM_Ty

图9 以噬菌体vB_PaeM_Ty衣壳蛋白构建的系统发育树Fig.9 Phylogenetic tree construction based on the capsid protein of phage vB_PaeM_Ty▲:噬菌体vB_PaeM_Ty推定的主要衣壳蛋白▲:Putative main capsid protein of phage vB_PaeM_Ty
3 讨论
铜绿假单胞菌感染给养殖业造成巨大的经济损失,其也是引起环境污染和严重医院感染的常见菌株。随着抗生素的滥用,多重耐药菌频繁出现,如何防治多重耐药菌,开发高效、广谱的噬菌体制剂替代抗生素,已成为目前研究的热点。目前,国内外关于铜绿假单胞菌噬菌体的研究数量相对较少。噬菌体有较强的宿主特异性,不同国家和地区的流行株也有差异。因此分离新的噬菌体,建立广泛的噬菌体库,可为噬菌体的临床应用提供重要的储备资源。
噬菌体作为细菌的特异性病毒,只特异性裂解特定病原菌,被视为安全的生物抗菌剂[24],对耐药菌株治疗有良好效果[25]。噬菌体在治疗铜绿假单胞菌感染中也展现出良好的效果,且具有可靠的安全性[26]。本试验从水中分离得到1株铜绿假单胞菌噬菌体vB_PaeM_Ty,根据形态判断该噬菌体为有尾病毒目(Caudovirales)、肌尾病毒科(Myoviridae)。通过裂解谱分析,发现噬菌体vB_PaeM_Ty仅能裂解宿主菌PA31和大肠杆菌-39、粪肠球菌-8a,表明vB_PaeM_Ty具有较强的宿主特异性。
噬菌体vB_PaeM_Ty的潜伏期较短,只有20 min,可见其能较快发挥杀灭宿主菌的作用。vB_PaeM_Ty裂解量约为142 PFU/mL,说明其裂解能力较强,具有开发为抗菌制剂的潜力。vB_PaeM_Ty在45、60和70 ℃放置60 min效价保持较高,在90 ℃ 放置60 min时仍具有活性。铜绿假单胞菌噬菌体PL39在80 ℃ 放置10 min完全失活[24];温度耐受性较好的铜绿假单胞菌噬菌体vB_PaeM_TG2在85 ℃放置1 h完全失活[27]。vB_PaeM_Ty在pH 3~12范围内活性稳定,与铜绿假单胞菌噬菌体的PsCh、PsIn、Ps25、Ps12on-D和PSJ-1的研究结果类似[28,29]。表明vB_PaeM_Ty具有良好的耐热性能和酸碱耐受性,这些为其临床应用提供了便利。
噬菌体vB_PaeM_Ty的DNA测序结果显示,其与铜绿假单胞菌噬菌体K4有较为亲近的同源关系。噬菌体K4 基因组长度为 50 358 bp,编码 77 个蛋白和1个 tRNA-Arg,Paundecimvirus属成员,能够显著抑制牛奶和午餐肉等样品中宿主菌的生长,有效控制食品中铜绿假单胞菌的污染[30,31]。提示本试验分离的vB_PaeM_Ty可能具有治疗铜绿假单胞菌感染的潜力。本试验功能预测分析发现,74个ORFs中25个为功能蛋白,其余为假定蛋白。其中,ORF56和ORF73分别编码内溶素和果胶裂解酶。内溶素是噬菌体感染细菌后期编码的肽聚糖水解酶,其在裂解宿主细菌细胞壁的肽聚糖,释放子代病毒粒子方面发挥重要作用,尤其是对dsDNA噬菌体[32,33]。研究表明,内溶素对肠球菌、金黄色葡萄球菌和猪链球菌等革兰阳性病原菌具有良好的杀菌作用[34-36]。果胶裂解酶是解聚酶的一种,可通过消除机制裂解 β-1,4糖苷键,降解细菌多糖成分,清除和抑制生物被膜的形成。Rice等[37]研究发现,尾刺蛋白中存在果胶裂解酶结构域,成功分离并表征了果胶裂解酶,验证了该酶在治疗细菌感染方面的潜力。因此,内溶素和果胶裂解酶在噬菌体杀灭细菌中具有重要作用,也有助于探究噬菌体-细菌的感染机制,这些特征为其开发为噬菌体制剂并应用于临床提供了理论依据。
目前,噬菌体治疗细菌感染仍有许多重要问题需要解决,如噬菌体宿主谱窄、细菌抗性产生快、保存条件严格等问题。因此,分离筛选更多的宽谱噬菌体,构建噬菌体库,通过鸡尾酒疗法提高噬菌谱,或对噬菌体裂解细菌机制、裂解酶、“穿孔素-内溶素”双组份裂解系统、尾端吸附机制、噬菌体-宿主互作等进行研究具有重要意义[24,38-40]。综上所述,本试验分离筛选到的噬菌体vB_PaeM_Ty,是1株烈性噬菌体,耐高温环境,且耐酸碱,包含多种功能相关蛋白,为铜绿假单胞菌噬菌体的文库构建和临床应用等后续研究提供了重要的理论依据。
