长江宜宾-江津段漂流性鱼卵纵向分布特征及与环境因子关系
2024-01-01陈小娟杨志朱其广曹俊董纯朱俊潘增





摘要:通过2023年4-7月对长江上游宜宾、合江和江津断面鱼类早期资源的逐日监测,了解漂流性鱼卵种类组成、鱼卵径流量、产卵高峰期及产卵场的纵向分布特征,分析逐日产卵径流量变动与水文、水温条件的关系,明晰不同断面鱼卵径流量峰值出现的流量区间,辨识影响鱼卵径流量变动的关键环境要素。结果表明:共采集到漂流性鱼卵24种,包括长江上游特有鱼类鱼卵8种,其中红唇薄鳅(Leptobotia rubrilabris)、长薄鳅(L. elongata)、长鳍吻鮈(Rhinogobio ventralis)为国家二级保护动物;流经宜宾、合江和江津断面的漂流性鱼卵总径流量分别为729.45×104、17 301.92×104和76 027.44×104粒,鱼卵总径流量从上游到下游呈明显增加的特征;3个断面鱼卵径流量的高峰期出现在5月下旬以后,其中宜宾断面最早,江津断面次之,合江断面最晚;金沙江一期工程蓄水后,保护区干流江段产卵场的纵向分布特征发生了明显改变,主要产卵场位置下移;流量上涨量、水位上涨量和流量日上涨率是影响3个断面鱼卵径流量变动的最重要因素。研究结果可为长江上游保护区干流鱼类早期资源保护、生态调度等提供科学依据。
关键词:产漂流性卵鱼类;鱼类早期资源;鱼卵径流量;产卵场;纵向分布;长江上游
中图分类号:S931.1" " " " 文献标志码:A" " " " 文章编号:1674-3075(2024)05-0193-11
长江上游珍稀特有鱼类国家级自然保护区(以下简称“保护区”)包括长江上游干流和赤水河、沱江、岷江、南广河等支流的部分江段,其中长江干流江段上起金沙江向家坝坝轴线下1.8 km 处,下至重庆地维大桥,其河道曲折,滩沱相间,复杂的地形地貌和发达的河网水系为鱼类提供了产卵、索饵和越冬等重要栖息地(高少波等,2013;孙志禹等,2014)。资料显示,历史上该保护区分布有25个产漂流性卵鱼类的集中产卵场(危起伟,2012),拥有复杂的鱼类区系组成和丰富的鱼类生态类群,是我国重要的淡水鱼类资源宝库(王梦等,2022),是连接金沙江下游和三峡库区生态系统的重要区域,对维系金沙江下游和长江上游淡水鱼类种质资源、鱼类生物多样性及生态系统稳定性具有重要意义(田辉伍等,2016)。
鱼类早期资源是反映鱼类种群数量变动的重要指标(易伯鲁等,1988)。产漂流性卵鱼类作为长江上游主要的鱼类生态类群之一,是长江上游鱼类生物多样性的重要组成部分,其早期资源变动能有效反映水生态系统健康状况(危起伟,2012)。有关该保护区鱼类早期资源的调查研究主要包括:吴国犀等(1988)和刘乐和等(1986;1990)重点对保护区江段的草鱼(Ctenopharyngodon idellus)、铜鱼(Coreius heterodon)和圆口铜鱼(Coreius guichenoti)等的产卵场分布特征及其与水文条件的关系进行了探讨;李祥艳等(2022)和胡兴坤等(2023)调查发现金沙江下游干流梯级水库蓄水后,宜宾江段仍分布有产漂流性卵鱼类的产卵场,但产卵量已明显下降;王龙飞等(2022)探究了泸州江段环境因子和水文过程对鱼类早期资源的影响,发现上游来水引起的流量增涨是影响该江段鱼类产卵的关键水文指标;众多学者(唐锡良,2010;段辛斌等,2015;高天珩等,2015;王涵等,2017;张先炳等,2021)开展了江津江段鱼类早期资源分布规律、铜鱼等产漂流性卵鱼类的繁殖规律以及产卵场分布的相关研究。这些鱼类早期资源的研究均集中在保护区干流的一个断面上,缺乏对纵向多断面分布特征的调查研究,也缺乏基于多断面数据的鱼卵径流量变动与环境条件综合分析。
本文于2023年4-7月,分别在保护区干流宜宾、合江和江津断面开展早期资源调查,获取流经各个断面的漂流性鱼卵种类组成、数量,以及鱼卵径流量、产卵出现时间、产卵高峰期及重要鱼类产卵场分布的信息,在此基础上,分析其纵向分布特征,比较不同断面间的差异,并分析所有产漂流性卵鱼类的逐日鱼卵径流量大小与环境条件的关系,辨识影响鱼类逐日鱼卵径流量变动的关键因素,以期为长江上游保护区江段鱼类早期资源保护提供基础数据支撑。
1" "材料与方法
1.1" "早期资源调查及数据获取
分别于2023年4月7日-7月15日在宜宾断面(104°37′23′′E,28°45′48′′N)、4月4日-7月19日在泸州江段合江码头断面(105°50′08′′E,28°48′45′′N)、4月6日-7月15日在江津通泰门断面(106°15′05′′ E,29°18′41′′ N)使用弶网(网长2.0 m,网口面积0.393 m2,网目40目;网后接30 cm×20 cm×25 cm的长方形集苗框)进行逐日卵苗采样,各采样断面分布如图1。
各断面位于河段顺直、水面宽阔的江段内,采样点设置在靠近河道深泓线一侧。每日在07:00-09:00、16:30-18:30两个时间段采样,根据鱼卵、仔稚鱼的数量以及水中悬移物的多少确定每次采集的持续时间,一般为15~60 min(曹文宣等,2007;雷欢等,2022;杨志等,2023)。每次采样时,记录采样起止时间、采样网具网口的入水水深、倾角,测定网口前流速、水温、透明度等,网口流速采用流速仪记录,断面逐时水温使用水下温度计自动记录,水体透明度使用萨氏盘测量。各次采样时的流速、水温及透明度取平均值。从全国水雨情信息网(http://xxfb.mwr.cn/sq_djdh. html)收集鱼类早期资源调查期间各个断面附近水文站或水位站(干流有向家坝坝下水文站、朱沱水文站及合江水位站;支流有赤水河赤水水文站)的流量和水位数据;合江断面流量数据采用朱沱站流量减去赤水河赤水站流量获得。
1.2" "样品处理及鱼卵种类鉴定
采样时将网内的鱼卵、鱼苗和悬浮物等进行过滤,分拣鱼卵和仔稚鱼并计数。鱼卵根据卵膜性质(黏沉性、漂流性、浮性)进行分类(曹文宣等,2007),观察及记录其发育期后,保存于95%的乙醇中,并采用分子生物学方法进行种类鉴定。鉴定步骤为:(1)提取卵或者鱼苗的mtDNA;(2)利用cytb或COI通用引物对所提取DNA进行扩增;(3)使用DNA STAR软件包中的Seq-Man模块对碱基序列进行检查;(4)在NCBI网(https://blast. ncbi. nlm.nih.gov/ BlastAlign. cgi)中进行序列比对,以序列相似度最高为鉴定标准,从而确定样品的种类(杨志等,2023)。
1.3" "数据处理
1.3.1" "鱼卵径流量及产卵场位置估算" "参照易伯鲁等(1988)的方法对各断面各调查日的鱼卵径流量进行估算。同时,根据所采鱼卵的发育时期,结合调查断面以上江段江水的平均流速,推算鱼卵的漂流距离,计算公式如下:
S=V×T" " " " ①
式中:S为鱼卵的漂流距离(m),V为江水平均流速(m/s),T为当时水温条件下胚胎发育经历的时间(s)。根据鱼卵的漂流距离,估算鱼类产卵时所处的江段位置。
1.3.2" "产卵时间及产卵高峰期" "根据不同江段鱼卵的逐日径流量,确定鱼卵出现的最早时间以及产卵高峰期。同时,对长薄鳅、长鳍吻鮈、铜鱼等重要鱼类鱼卵出现的最早时间进行统计分析,了解这些鱼类鱼卵最早出现时间上的差异。
1.3.3" "逐日鱼卵径流量大小与环境条件的关系" "为尽可能地消除产卵亲鱼数量差异以及产卵场分布不均衡对产卵丰度的影响,本文使用逐日鱼卵径流量大小的相对比例代替其绝对值(King et al,2016)。考虑到不同采样断面间流量固有的差异,使用散点图分析逐日平均流量与鱼卵日径流量比例的关系,从而获得鱼卵径流量比例出现峰值时的流量范围;同时,使用随机森林模型(random forest,RF)(杨志等,2023),分析监测期间流经3个监测断面的所有产漂流性卵鱼类的逐日产卵径流量大小与环境条件(主要为变动值参数,以及受河道地形影响较小的参数)的关系(表1)。在构建RF模型时,以各断面漂流性卵逐日径流量的比例为因变量,以14个环境变量(表1)为自变量。 这14个变量 采用leave-one-out方法作为RF的验证程序;采用指标%IncMSE(% increase in mean squared error,表示增加的均方误差百分比)和IncNodePurity(increase in node purity,表示增加的节点纯度)判别各自变量对因变量影响的相对重要性,并绘制重要影响变量以及常受关注的环境变量(持续涨水的天数)对因变量的偏效应(partial effect)图。
2" "结果与分析
2.1" "鱼卵种类组成、数量及纵向分布特征
2023年4-7月,在宜宾、合江和江津断面共采集到漂流性卵6 447粒,其中宜宾断面采集到203粒、合江714粒、江津5 530粒。经鉴定,3个监测断面共采集到漂流性卵25种,上游特有鱼类的鱼卵有红唇薄鰍、宽体沙鳅(Sinibotia reevesae)、小眼薄鰍(L. microphthalrna)、异鳔鳅鮀(Xenophysogobio boulengeri)、圆筒吻鮈(Rhinogobio cylindricu)、长薄鳅、长鳍吻鮈、中华金沙鳅(Jinshaia sinensis)等8种,其中红唇薄鳅、长薄鳅、长鳍吻鮈为国家二级保护动物。
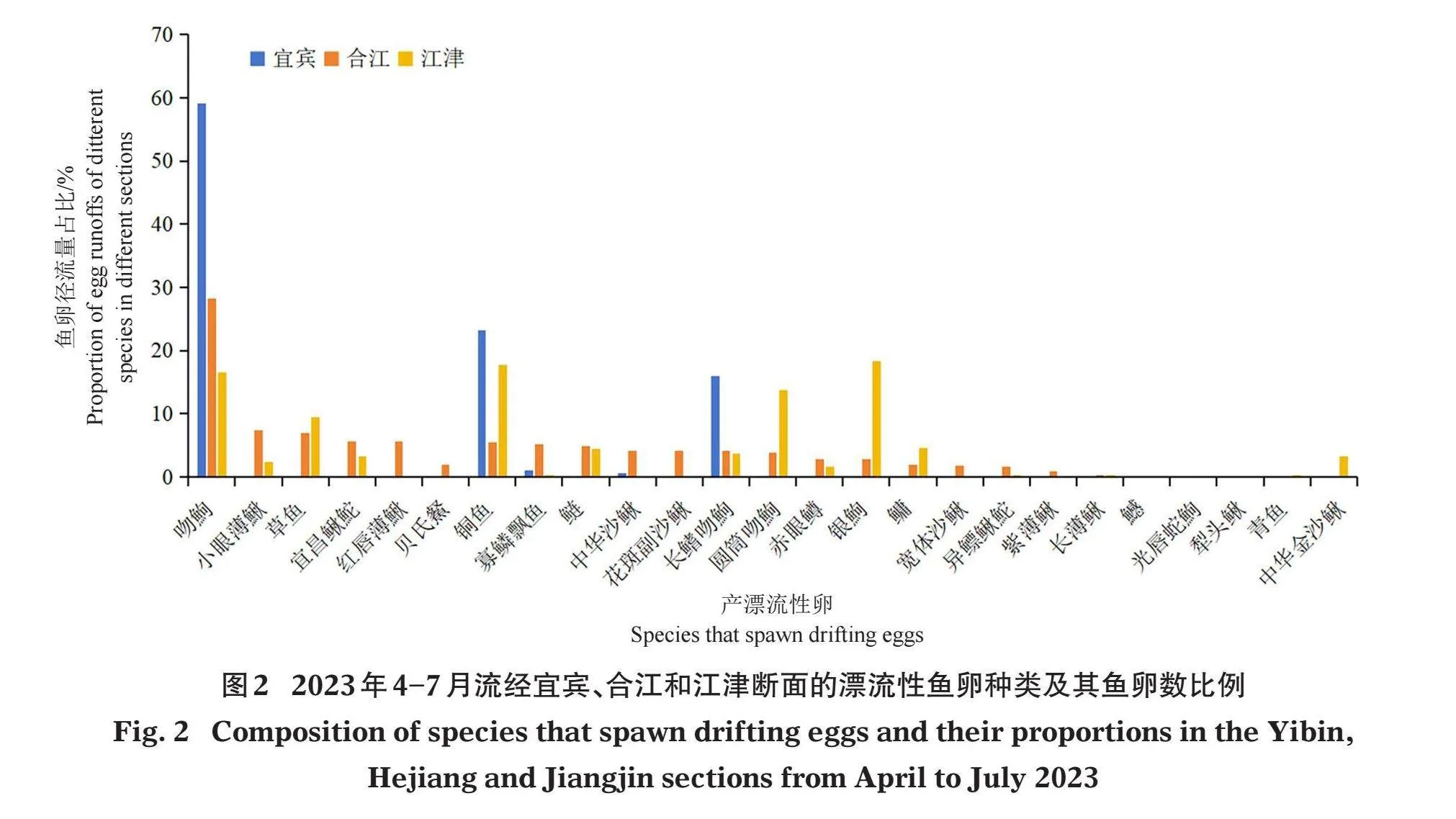
不同种类在各个断面被采集到的漂流性卵数比例见图2。结果显示:在宜宾断面,共采集到吻鮈(R.typus)、铜鱼、长鳍吻鮈、寡鳞飘鱼(Pseudolaubuca engraulis)和中华沙鳅(S. superciliaris) 5种鱼的鱼卵,其中吻鮈、铜鱼和长鳍吻鮈卵数量位于前3位,分别占该断面鱼卵总采集数量的59.09%、23.23%和16.04%;在合江断面,共采集到吻鮈、铜鱼、长鳍吻鮈、宜昌鳅鮀(Gobiobotia filifer)、小眼薄鰍等鱼卵20 种,其中吻鮈、小眼薄鳅和草鱼卵数量比例占前3位,分别为28.18%、7.44%和6.94%;在江津断面,共采集到银鮈(Squalidus argentatus)、吻鮈、铜鱼、长鳍吻鮈、宜昌鳅鮀等鱼卵19种,其中银鮈、铜鱼、吻鮈数量比例占前3位 ,分别为18.42%、17.70%和16.57%。对比合江和江津断面,仅在合江断面能够采集到的鱼卵种类有红唇薄鰍、中华沙鳅、宽体沙鳅、花斑副沙鳅(Parabotia fasciata)和紫薄鳅(L. taeniaps);仅在江津断面能够采集到的鱼卵种类有中华金沙鳅、鱤(Elopichthys bambusa)、犁头鳅(Lepturichthys fimbriat)、青鱼(Mylopharyngodon piceus)和光唇蛇鮈(Saurogobilo gymnocheilus)。
在鱼卵的数量比例上,沿河流纵向,从上游到下游,采集到的吻鮈卵数量比例逐渐下降,而其他4种鱼卵(铜鱼、长鳍吻鮈、寡鳞飘鱼和中华沙鳅)的数量比例均呈现先在合江断面增加,然后江津断面减少的特征;小眼薄鳅和异鳔鳅鮀卵的数量比例先在合江江段明显增加,然后在江津江段减少;宜昌鳅鮀和鲢(Hypophthalmichthys molitrix)鱼卵的数量比例呈先明显上升,再略有减少的特征;银鮈、圆筒吻鮈、草鱼(Ctenopharyngodon idellus)和鳙(Aristichthys nobilis)鱼卵的数量比例则呈一直上升的特征(图2)。
2.2" "鱼卵径流量
监测期间,流经宜宾、合江和江津断面的漂流性鱼卵总径流量为729.45×104、17 301.92×104和76 027.44×104粒,鱼卵总径流量从上游到下游呈明显增加的特征;产漂流性卵的不同种类在各断面上的鱼卵径流量显示:在宜宾断面,吻鮈卵的径流量最大,为431.04×104粒,其次为铜鱼,为169.47×104粒;在合江断面,吻鮈卵的径流量最大,为4 876.51×104粒,其次为小眼薄鰍和草鱼,分别为1 287.85×104粒和1 201.38×104粒;在江津断面,银鮈卵的径流量最大,为14 001.17×104粒;其次为铜鱼、吻鮈和圆筒吻鮈,分别为13 453.60×104、12 596.03×104和10 418.59×104粒(图3)。
沿河流纵向,从上游到下游,银鮈、铜鱼、吻鮈、长鳍吻鮈、圆筒吻鮈、赤眼鳟(Squaliobarbus curriculus)、草鱼、鲢、鳙、宜昌鳅鮀、小眼薄鳅、长薄鳅和中华金沙鳅等12种鱼类的鱼卵径流量呈明显增加趋势,越靠近下游,适应三峡库区生境的鱼类如“四大家鱼”、铜鱼、银鮈、赤眼鳟等的鱼卵径流量越大(图3)。特有鱼类中,红唇薄鳅、宽体沙鳅仅出现在合江断面,异鳔鳅鮀的鱼卵径流量以合江断面最大,而长鳍吻鮈、圆筒吻鮈的鱼卵径流量以江津断面最大,中华金沙鳅仅出现在江津断面;在合江和江津断面鱼卵径流量均较大的特有鱼类有小眼薄鳅(图3)。
2.3" "鱼卵出现时间及产卵高峰期
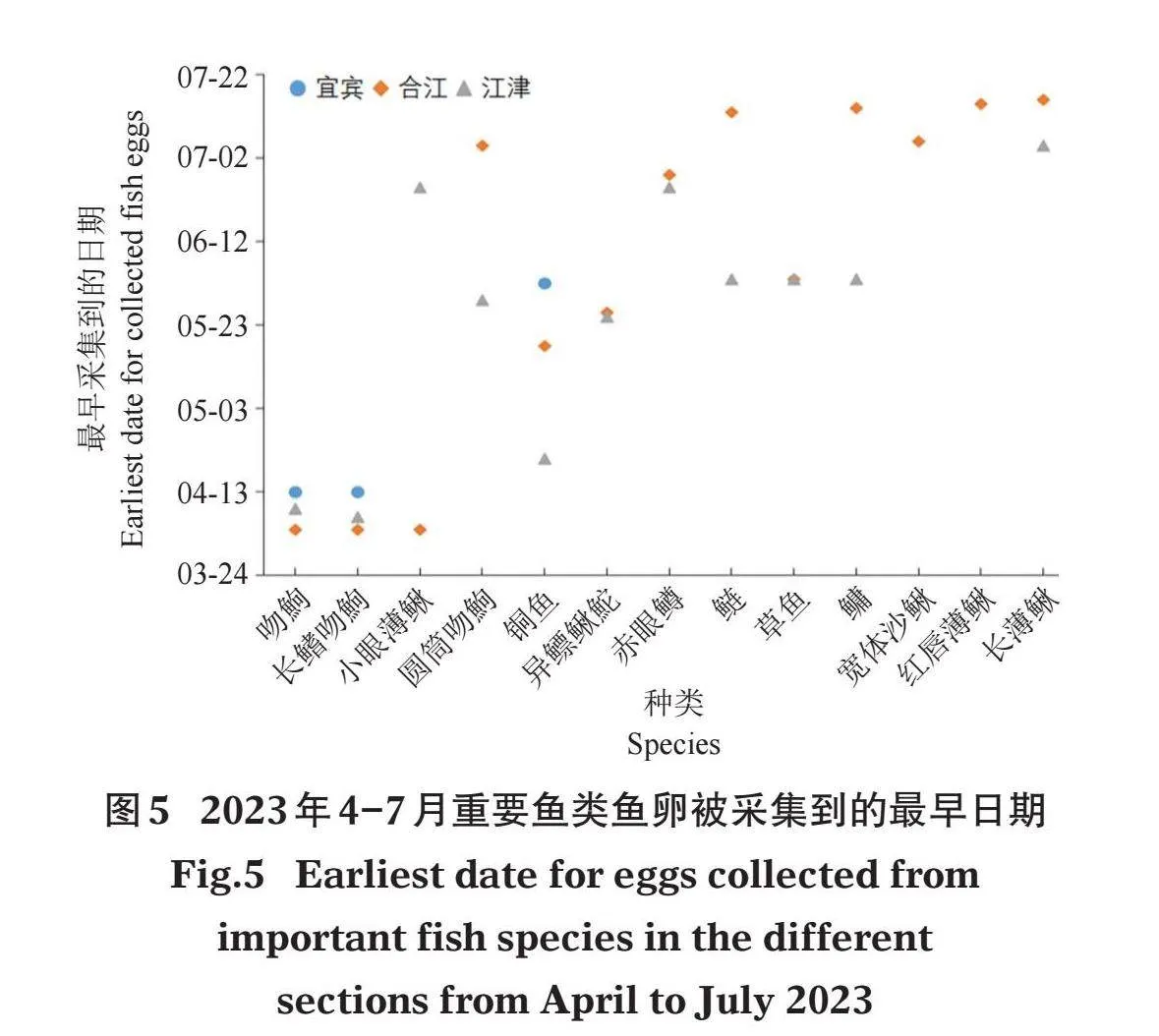
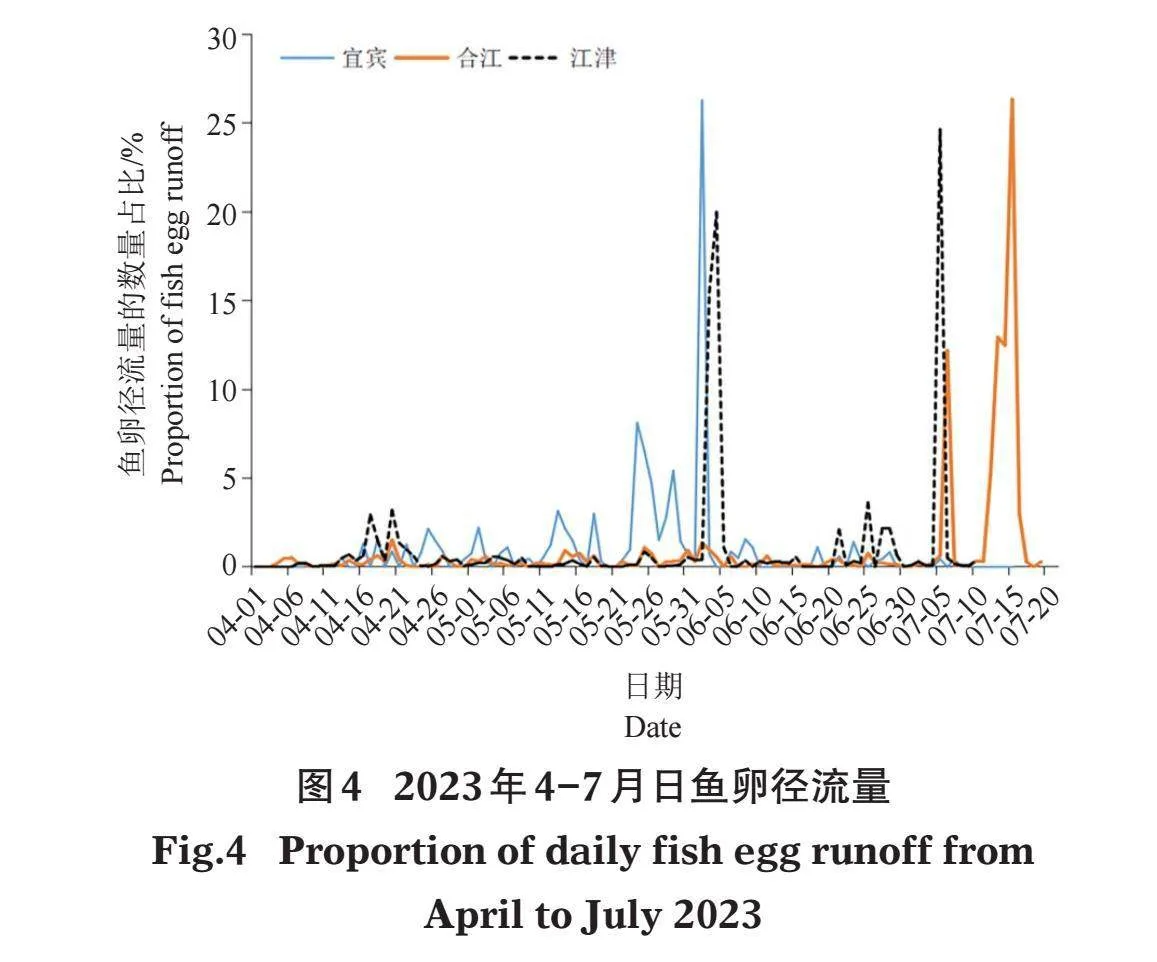
监测期间,鱼卵在宜宾、合江和江津断面最早被采集到的时间分别为4月13日、4月4日和4月7日,合江和江津断面在监测结束时(7月19日和7月15日)仍能采集到鱼卵,而宜宾断面在7月中旬产卵已基本结束。宜宾断面在4月中旬会出现小的产卵高峰,但大的产卵高峰则在5月下旬及以后出现,产卵高峰期出现在5月24-29日和6月2日,此期间鱼卵径流量分别占该断面总鱼卵径流量的29.05%和26.30%;合江断面产卵高峰期出现在7月6日和7月13-15日,此期间鱼卵径流量分别占12.21%和51.79%;江津断面产卵高峰期出现在6月3-4日和7月5日,此期间鱼卵径流量分别占35.46%和24.65%(图4)。
吻鮈、长鳍吻鮈和小眼薄鳅的鱼卵最早,在4月上旬即可被采集到;随后是铜鱼卵,其在4月中旬后即可被采集到;其他鱼类包括圆筒吻鮈、草鱼、鲢、鳙等的鱼卵在5月下旬及以后才能被采集到,长薄鳅的鱼卵在7月上旬及以后才能被采集到(图5)。
2.4" "重要鱼类产卵场分布
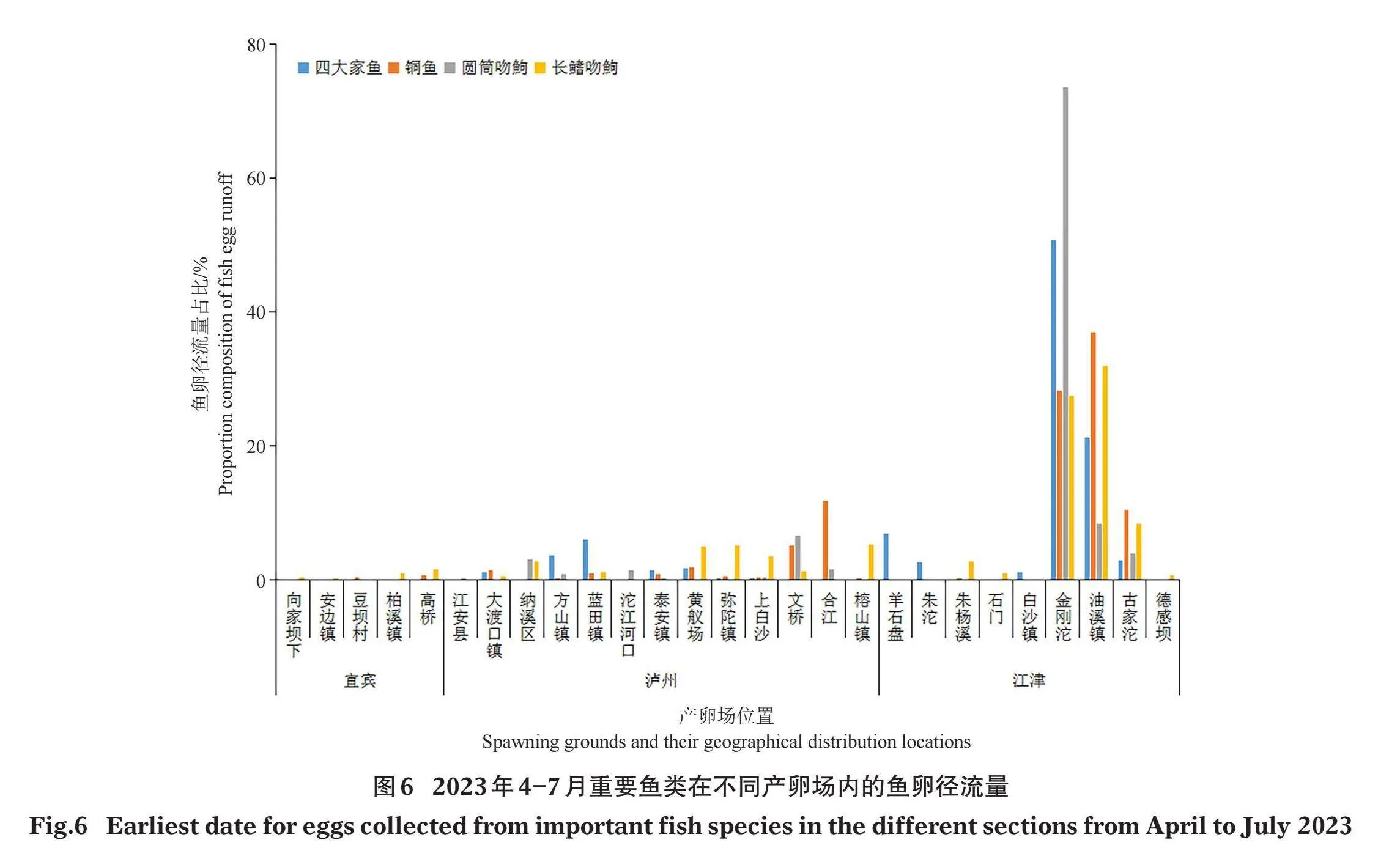
宜宾至江津断面广泛分布有铜鱼、长鳍吻鮈、圆筒吻鮈等产漂流性卵鱼类的产卵场,其中产卵量较大的产卵场主要分布在合江至江津约110 km的江段(鱼卵径流量比例gt;5%)(图6)。具体而言,铜鱼在保护区干流宜宾至江津段内的集中产卵场有5处,分别位于泸州文桥、合江县、重庆金刚沱、油溪镇和古家沱江段,这5个产卵场来源的鱼卵径流量比例分别为5.08%、11.81%、28.21%、37.01%和10.43%;“四大家鱼”的集中产卵场有4处,分别位于泸州蓝田镇、重庆羊石盘、金刚沱和油溪镇江段,鱼卵径流量分别占5.96%、6.89%、50.69%和21.29%;圆筒吻鮈的集中产卵场有3处,分别位于泸州文桥、重庆金刚沱和油溪镇江段,占6.54 %、73.56%和8.32%;长鳍吻鮈的集中产卵场有5 处,位于泸州弥陀镇、榕山镇、重庆金刚沱、油溪镇和古家沱江段,分别占5.15 %、5.20%、27.53 %、31.86%和8.38%。
2.5" "逐日鱼卵径流量大小与环境条件的关系
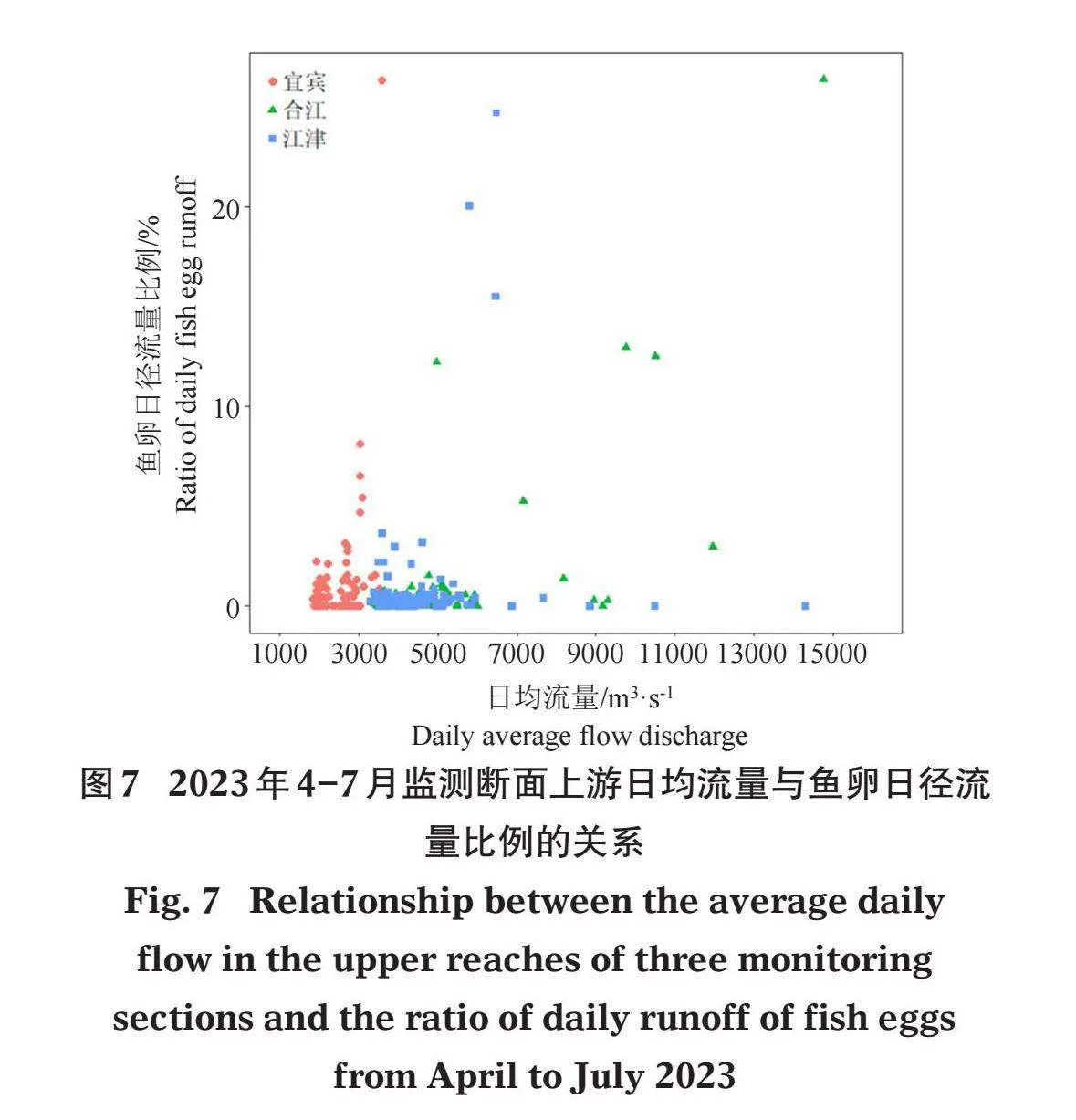
当宜宾断面上游流量为3 000~3 800 m3/s时,流经宜宾断面的鱼卵的日径流量占监测期间总鱼卵径流量的比例相对较高(4.67%~26.30%);当合江断面上游流量在5 000 m3/s以上时,该断面的鱼卵日径流量占比相对较高(2.98%~26.36%);当江津断面流量为4 500 ~6 500 m3/s时,该断面的鱼卵日径流量占比相对较高(3.20%~24.65%)(图7)。
随机森林模型结果显示:14个环境因子中,流量上涨量(FIQ)、水位上涨量(WRQ)和流量日上涨率(DIRF)的%InMSE值和IncNodePurity值,与其他因子的值明显分离,且大于这些因子的值,表明这3个因子为影响3个监测断面鱼卵日径流量比例的最重要因子(图8)。14个环境因子可解释各断面鱼卵日径流量比例80.17%的变异。
基于随机森林模型,得到影响监测断面鱼卵日径流量比例变动的重要变量的偏效应图(图9)。总体上,鱼卵日径流量比例随着水位上涨量和流量上涨量的增加呈现缓慢上升-迅速上升-迅速下降-保持相对稳定-迅速上升-保持相对稳定的趋势。当水位上涨1.4 m以上或流量上涨大于1 500 m3/s时,鱼卵日径流量比例较高。随着流量日上涨率增加,鱼卵日径流量比例呈现先缓慢上升,再迅速上升的趋势。当流量日上涨率为1 580 m3/s时,鱼卵日径流量比例较高;当持续涨水天数大于5 d时,各断面的鱼卵日径流量比例较高。
3" "讨论
3.1" "鱼卵纵向分布特征
本研究在宜宾断面采集到的漂流性卵的种类数明显少于合江和江津断面。合江和江津断面间漂流性卵种类数差异很小(仅相差1种),表明距离向家坝较近时,因大坝至采样断面间流水江段短且受大坝阻隔影响(胡兴坤等,2023),流经宜宾断面漂流性卵鱼类种类数少;当距离向家坝较远时,由于流水江段长且有支流鱼卵汇入的可能性,漂流性卵的种类数会明显增加。同时,从鱼卵种类组成看,合江和江津断面间差异较小,但有少数种类鱼卵仅在合江或江津断面采集到,如红唇薄鳅和宽体沙鳅仅见于合江断面,中华金沙鳅、青鱼、鱤、光唇蛇鮈和犁头鳅仅见于江津断面。造成不同断面鱼卵种类组成差异的原因可能与鱼类在不同江段的地理分布差异有关,如唐成等(2023)调查发现,合江和江津江段间鱼类群落存在明显差异;高天珩等(2013)对保护区干流江段的渔获物调查显示,在宜宾、合江和江津3个采样江段中,宽体沙鳅仅在合江江段出现。此外,不同断面间鱼卵种类组成差异也可能与离三峡水库的远近有关,如仅在江津断面采集到的部分鱼卵种类(如青鱼、鱤等)均为适应库区生境的鱼类(图3)。
漂流性卵的径流量呈现明显的纵向分布特征,沿河流纵向,越往下游,流经断面的鱼卵数量及径流量越大(图3)。刘乐和等(1990)对长江上游各断面铜鱼产卵规模纵向分布特征的研究也观测到该现象。该现象的出现与河流纵向鱼卵来源及亲鱼资源丰度增加有关。许多针对鱼类群落结构的纵向梯度调查研究表明,越往流域下游,鱼类资源丰度及物种多样性越高(Vašek et al,2016; Guedes et al,2020; López-Rodríguez et al,2024),而亲鱼数量及物种多样性的增加,会使鱼卵数量及种类来源的多样性增加。在本文中,由于金沙江下游干流梯级对鱼卵下行的阻隔影响(胡兴坤等,2023),宜宾断面的鱼卵主要来源于向家坝坝址至宜宾断面间的30 km江段,产卵场范围及可容纳的亲鱼数量有限,而宜宾以下至江津江段,多为宽谷江段,岸坡缓坦、水道弯曲、江心岛及州滩密布,一直以来是鱼类资源丰富及产卵场密布的江段(危起伟,2012;周岐兵等,2022;朱俊等,2024)。同时,此江段有岷江、沱江、赤水河等大型支流汇入,客观增加了下游断面鱼卵的来源(刘飞等,2019; 吕浩等,2019)。此外,从鱼卵径流量的纵向分布特征来看,适应库区生境的鲢、鳙、草鱼、赤眼鳟等鱼类在江津断面的鱼卵径流量明显大于合江断面(图3),表明三峡库区鱼类的上溯产卵可能会对江津断面鱼卵径流量变动造成影响,这仍需要进一步调查研究确定。
2023年鱼卵径流量高峰期在不同断面间并未呈现明显的纵向分布特征,其中宜宾断面出现时间最早,江津断面次之,最晚为合江断面(图5),这可能与不同断面上游产卵亲鱼种类组成及数量差异有关。已有调查发现,不同鱼类种类产卵繁殖时间存在差异(曹文宣等,2007; 朱俊等,2024),加上产卵亲鱼数量的差异,产卵高峰期时间会出现差异。本文中,流经宜宾断面的主要鱼卵种类为吻鮈和铜鱼,其产卵时间较早,江津断面除吻鮈和铜鱼以外,还包括圆筒吻鮈、鲢、鳙等产卵时间较晚的种类,而合江江段则包括红唇薄鳅、小眼薄鳅等产卵时间很晚(通常在6月下旬后才开始产卵)的种类(图3和图5),产卵时间差异加上不同断面亲鱼数量的差异导致不同江段间产卵高峰期的差异。此外,鱼卵径流量高峰期的纵向分布特征,也可能与近年来保护区各断面早期资源高峰期的时间动态改变有关。已有研究发现,宜宾断面2018-2020年产漂流性卵鱼类自然繁殖的高峰期与2016-2017年相比提前(胡兴坤等,2023),而江津断面2019-2022年产漂流性卵鱼类自然繁殖的高峰期呈推后的特征(朱俊等,2024),鱼类早期资源高峰期在不同断面上的变化仍需进一步开展相关监测分析研究。
3.2" "产卵场纵向分布特征
段辛斌等(2015)2010-2012年在江津断面的鱼类早期资源调查发现,在金沙江下游一期工程蓄水前,产漂流性卵鱼类在保护区干流江段内的产卵场主要分布在泸州市的榕山镇、合江县和弥沱镇江段。朱俊等(2024)2019-2022年在江津断面的调查发现,在江津以上170 km江段广泛分布着产漂流性卵鱼类的产卵场,其产卵场主要分布在泸州市的弥陀镇以及江津区的朱杨溪、石门镇、金刚沱和油溪镇江段,其中金刚沱和油溪镇为产卵规模排在前2位的产卵场。本文结果表明,主要产卵场分布在江津区内的金刚沱和油溪镇,与以上研究结果均有所差异。这表明:(1)金沙江一期工程蓄水后,保护区干流江段产卵场的纵向分布特征发生了明显改变,鱼类主要产卵场在保护区江段内的位置明显下移,逐渐分布于保护区干流江段的下游区域;(2)相邻年份间主要产卵场的分布位置也会存在一定的差异。导致产卵场下移的原因,除与金沙江下游梯级水库引起的低温水下泄以及水沙情势改变有关以外(王兴敏,2019; Zhou et al,2024),也很可能与三峡水库的运行有关,如图5所示,流经江津断面的鱼卵中很多是鲢、鳙、草鱼、赤眼鳟等的鱼卵,而这些鱼类通常在三峡库区江段内栖息觅食。王震等(2019)对寡鳞飘鱼仔鱼的发生动态和丰度的分析发现,三峡水库与保护区干流江段组合而成的河-库复合生态系统,在产漂流性卵鱼类的早期补充中起着重要作用。
3.3" "影响漂流性鱼卵径流量变动的生境条件
已有调查研究发现,产漂流性卵鱼类产卵繁殖与适宜的水温及流量过程条件密切相关(易伯鲁等,1988; 王龙飞等,2022; 朱俊等,2024),产卵多发生在江河的涨水过程中,水位升高、流量增大、流速加快、流态紊乱、透明度减小等多种水文因素在洪峰过程中的变化,综合影响着产漂流性卵鱼类的产卵繁殖。产卵前持续涨水的天数、流量、水位及透明度的变幅以及产卵时的流量大小等通常被认为是影响产漂流性卵鱼类鱼卵径流量变动的关键生境条件(易伯鲁等,1988,王龙飞等,2022; 杨志等,2023;朱俊等,2024)。在保护区干流区域,王龙飞等(2022)根据2019-2020年在泸州断面的调查结果,认为流经该断面的鱼卵密度与向家坝出库量、水体透明度以及断面水位显著相关,繁殖季节上游来水引起的流量增涨是影响产漂流性卵鱼类产卵的关键因素。王兴敏(2019)基于产卵场实测数据及一维水动力-水温模拟结果,认为保护区干流漂流性鱼类的产卵繁殖主要受流速、水温、日均水位上涨率和持续涨水天数的影响,其中适宜的日均水位上涨率为0.2~0.3 m/d,适宜的涨水持续时间为 5~7 d。朱俊等(2024)对2019-2022年江津断面早期资源调查分析显示,洪峰过程中的水位涨幅、初始流量和透明度是影响保护区江津断面产漂流性卵鱼类繁殖规模大小的重要因素。本文结果显示,流量上涨量、水位上涨量和流量日上涨率是影响保护区干流3个断面鱼卵径流量变动的最重要因素,也表明流量的增涨过程在保护区干流产漂流性卵鱼类产卵繁殖中扮演着重要的角色。尽管本研究与其他在保护区干流江段进行的类似研究(王龙飞等,2022;朱俊等,2024)使用了相同的统计参数如持续涨水天数、水位上涨量等,但是不同研究得出的影响鱼卵径流量变化的关键因素存在差异。这种差异出现的原因可能与统计数据的来源有关,王龙飞等(2022)和朱俊等(2024)的研究结果均基于单一断面的监测数据,而本文的统计数据则基于多断面的监测数据。已有研究发现,不同鱼类种类对水文条件的需求存在较大差异,银鮈、吻鮈等鱼类产卵水文要求低,铜鱼、圆筒吻鮈、鲢、鳙等鱼类产卵对水文的要求较高(Jiang et al,2010; 黎明政等,2019; 朱俊等,2024),而这些鱼类在不同断面间的鱼卵径流量比例存在明显差别。这种差别会对不同断面鱼卵径流量变动影响因素的最终分析结果产生影响(段辛斌等,2015;朱俊等,2024),而基于多断面监测数据的统计分析有助于获得更为全面的结果。
3.4" "保护区干流产漂流性卵鱼类产卵繁殖保护建议
调查发现,保护区干流断面不仅可采集到红唇薄鰍、长鳍吻鮈、长薄鳅等8种上游特有鱼类或国家保护鱼类的鱼卵,而且也可采集到铜鱼、鲢、鳙等重要种质资源鱼卵或经济鱼类的鱼卵,表明在长江上游梯级水电开发背景下,保护区干流江段是长江流域重要鱼类早期资源保护的关键江段,因此保护该江段的生境完整性对于长江上游鱼类资源的保护具有重要的意义。在此背景下,保护区中开展的经济生产活动应尽量避免对河道生境的影响。
相较之金沙江下游一期工程蓄水前,目前产漂流性卵鱼类的主要产卵场已明显下移,造成该现象的可能原因,除了金沙江下游梯级水库的蓄水运行对保护区干流江段产卵场分布的空间特征的影响以外,还与三峡水库形成的河-库复合生态系统有关(王震等,2019)。建议持续实施生态调度恢复保护区干流江段的水文情势,促进鱼类自然繁殖,同时开展三峡水库鱼类资源持续监测,关注三峡库区鱼类群落结构的演变特征。
鱼卵径流量峰值出现的流量区间在不同断面间存在明显差异。这就需要通过生态调度使3个断面上游江段具有各自的适宜流量范围。考虑到不同断面鱼卵种类组成及产卵时间的差异,建议在4月中下旬,保持宜宾断面上游流量在3 000 m3/s以上,促进产卵早的长鳍吻鮈、吻鮈等鱼类的产卵繁殖;在5月下旬,保持江津断面流量在4 500 m3/s以上,促进铜鱼、圆筒吻鮈等鱼类的产卵繁殖;在6月下旬及以后,保持合江断面上游流量在5 000 m3/s以上,促进红唇薄鳅、长薄鳅、“四大家鱼”等鱼类的产卵繁殖。同时,为使保护区干流不同断面的鱼卵径流量具有较大规模,建议不同断面上游产卵场的水位上涨量大于1.4 m、流量上涨量大于1 500 m3/s、流量日上涨率大于1 580 m3/s,并在满足上述条件满足的前提下保持持续涨水天数在5 d以上。
参考文献
曹文宣, 常剑波, 乔晔, 等, 2007. 长江鱼类早期资源[M].北京: 中国水利水电出版社.
段辛斌, 田辉伍, 高天珩, 等, 2015. 金沙江一期工程蓄水前长江上游产漂流性卵鱼类产卵场现状[J].长江流域资源与环境, 24(8): 1358-1365.
高少波, 唐会元, 乔晔, 等,2013.金沙江下游鱼类资源现状研究[J].水生态学杂志, 34(1): 44-49.
高天珩, 田辉伍, 叶超, 等, 2013. 长江上游珍稀特有鱼类国家级自然保护区干流段鱼类组成及其多样性[J].淡水渔业, 43(2): 36-42.
高天珩, 田辉伍, 王涵, 等, 2015. 长江上游江津断面铜鱼鱼卵时空分布特征及影响因子分析[J].水产学报, 39(8): 1099-1106.
胡兴坤, 邵科, 阙延福, 等, 2023. 金沙江下游宜宾段产漂流性卵鱼类早期资源动态变化[J].水生态学杂志, 44(2):73-80.
刘乐和, 吴国犀, 曹维孝, 等, 1986. 葛洲坝水利枢纽兴建后对青、草、鲢、鳙繁殖生态效应的研究[J].水生生物学报, (4): 353-364.
刘乐和, 吴国犀, 王志玲, 1990. 葛洲坝水利枢纽兴建后长江干流铜鱼和圆口铜鱼的繁殖生态[J].水生生物学报, (3): 205-215.
刘飞, 张富斌, 王雪, 等, 2019. 赤水河产漂流性卵鱼类的繁殖活动及其与环境因子之间的关系[J].水生生物学报, 43(增刊): 77-83.
黎明政, 马琴, 陈林, 等, 2019. 三峡水库产漂流性卵鱼类繁殖现状及水文需求研究[J].水生生物学报, 43(增刊): 84-96.
李祥艳,田辉伍,蒲艳,等,2022.长江上游宜宾江段鱼类早期资源现状研究[J].渔业科学进展,43(4):93-104.
吕浩, 田辉伍, 申绍祎, 等, 2019. 岷江下游产漂流性卵鱼类早期资源现状[J].长江流域资源与环境, 28(3): 586-593.
雷欢, 陈锋, 谢文星, 等, 2022. 汉江中下游梯级枢纽联合生态调度对产漂流性卵鱼类自然繁殖的效应研究[J].湖泊科学, 34(4): 1219-1233.
孙志禹, 张敏, 陈永柏, 2014. 水电开发背景下长江上游保护区珍稀特有鱼类保护实践[J].淡水渔业, 44(6): 3-8.
唐锡良, 2010. 长江上游江津江段鱼类早期资源研究[D].重庆: 西南大学.
唐成, 邓华堂, 田辉伍, 等, 2023. 长江上游珍稀特有鱼类国家级自然保护区干流段鱼类群落结构特征分析[J].水产学报, 47(2): 79-98.
田辉伍, 何春, 刘明典, 等, 2016. 长江上游干流三层流刺网渔获物结构研究[J].淡水渔业, 46(5): 37-42.
危起伟, 2012. 长江上游珍稀特有鱼类国家级自然保护区科学考察报告[M].北京: 科学出版社.
吴国犀, 刘乐和, 王志玲, 等, 1988. 长江上游金沙江江段草鱼自然繁殖的研究[J].淡水渔业, (1): 3-6.
王涵, 田辉伍, 陈大庆, 等, 2017. 长江上游江津段寡鳞飘鱼早期资源研究[J].水生态学杂志, 38(2): 82-87.
王震, Arunjith T S, 谢松光, 等, 2019. 金沙江梯级大坝运行和三峡水库运行水位增高对长江上游干流寡鳞飘鱼仔鱼丰度和分布的影响[J].水生生物学报,43(3): 606-611.
王兴敏, 2019.电站运行对向家坝下游产漂流性卵鱼类生境的影响及生态调度效果评价[D].成都: 四川大学.
王龙飞, 田辉伍, 严忠銮, 等, 2022. 长江上游泸州江段鱼类早期资源现状及其与水文条件响应关系[J].长江流域资源与环境, 31(4): 814-822.
王梦, 杨鑫, 王维, 等, 2022. 基于eDNA技术的长江上游珍稀特有鱼类国家级自然保护区重庆段鱼类多样性研究[J].水生生物学报, 46(1): 2-16.
易伯鲁, 余志堂, 梁秩燊, 等, 1988. 葛洲坝水利枢纽与长江四大家鱼[M].武汉: 湖北科学技术出版社.
杨志, 龚云, 胡挺, 等, 2023. 三峡库区涪陵江段产漂流性卵鱼类的早期资源及其对水文和水温特征的响应[J].湖泊科学, 35(2): 586-598.
张先炳, 杨胜发, 杨威, 等, 2021. 长江上游宜宾-江津与涪陵-丰都江段鱼类早期资源分布研究[J].淡水渔业, 51(5): 51-59.
周岐兵, 程飞, 王震, 等,2022. 长江上游合江江段鱼类早期资源与向家坝水库生态调度效果初步研究[J].中国环境监测, 38(1): 95-103.
朱俊, 曹俊, 周涛, 等, 2024. 长江上游江津段产漂流性卵鱼类早期资源现状及繁殖需求[J].水生态学杂志, 45(1):75-86.
Guedes G H S, Mattos T M, da Silva Camilo G, et al, 2020. Artificial flow regime promotes abiotic and biotic gradients: Testing the concept of longitudinal zonation in an off-river reservoir[J]. Ecohydrology amp; Hydrobiology, 20(2): 256-264.
Jiang W, Liu H Z, Duan Z H, et al, 2010. Seasonal variation in drifting eggs and larvae in the Upper Yangtze, China[J]. Zoological Science, 27(5): 402-409.
King A J, Gwinn D C, Tonkin Z, et al, 2016. Using abiotic drivers of fish spawning to inform environmental flow management[J]. Journal of Applied Ecology, 53: 34-43.
López-Rodríguez A, Meerhoff M, D’Anatro A, et al, 2024. Longitudinal changes on ecological diversity of Neotropical fish along a 1700 km river gradient show declines induced by dams[J]. Perspectives in Ecology and Conservation, 22(2): 186-195.
Vašek M, Prchalová M, Říha M, et al, 2016. Fish community response to the longitudinal environmental gradient in Czech deep-valley reservoirs: implications for ecological monitoring and management[J]. Ecological Indicators, 63: 219-230.
Zhou Y J, Wang G Q, Wu H L, et al, 2024. Changes in flow and sediment transport caused by cascade hydropower in the upper reaches of Yangtze River and their influence on spawning of Coreius heterodon [J]. CATENA, 235: 107622.
(责任编辑" "郑金秀)
Longitudinal Distribution of Drifting Fish Eggs from Yibin to Jiangjin in the Upper Yangtze River and the Relationship with Environmental Factors
CHEN Xiao‐juan1, YANG Zhi1, ZHU Qi‐guang1, CAO Jun1, DONG Chun1, ZHU Jun2, PAN Zeng2
(1. Institute of Hydroecology, Ministry of Water Resources and Chinese Academy of Sciences,
Key Laboratory of Ecological Impacts of Hydraulic-Projects and Restoration of
Aquatic Ecosystem of Ministry of Water Resources, Wuhan 430079, P.R. China.;
2. China Three Gorges Group Co., Ltd., Wuhan 430010, P.R. China)
Abstract:Early-stage fish resources are an important indicator that reflect variations in fish community dynamics. Fish species that spawn drifting eggs are one of the main ecological fish groups in the upper Yangtze River and an important component of fish species diversity. Therefore, variation of early-stage resources of these species effectively reflect the health of the aquatic ecosystem. In this study, we carried out daily monitoring of early-stage fish resources in the Yibin, Hejiang and Jiangjin sections of the upper Yangtze River from April to July 2023. Based on the monitoring results, we analyzed species composition of eggs, egg runoff, peak spawning period and the longitudinal distribution of spawning grounds for species that release drifting eggs. The relationship between daily variation of fish egg runoff and environmental variables (hydrologic conditions and water temperature) was then analyzed. Finally, the flow interval where peak egg runoff appeared in the different sections, and the key environmental factors affecting fish egg runoff were identified. A total of 24 fish species that spawn drifting eggs were collected, including 8 species endemic to the upper Yangtze River, among which are three species (Leptobotia rubrilabris, L. elongata, and Rhinogobio ventralis) that are national second-class protected animals. The total runoff of drifting eggs flowing through the Yibin, Hejiang and Jiangjin sections were, respectively, 729.45×104, 17 301.92×104 and 76 027.44×104, and the total runoff of fish eggs clearly increased from upstream to downstream. The peak of fish egg runoff in all three sections appeared after the end of May, with the Yibin section being the earliest (May 24-29, June 2), followed by the Jiangjin section (June 3-4, July 5) and the Hejiang section (July 6, July 13-15). After the first phase of the project, when Jinsha River was impounded, the longitudinal distribution of spawning grounds in the mainstem of the upper Yangtze River changed markedly, and the main spawning grounds moved downstream. The increases in discharge, water level and daily discharge rate were the most important factors affecting the variation of fish egg runoff in the three sections. These results provide a scientific basis for implementing early fish resource protection measures, especially the regulation of ecological flow in the upper reaches of the upper Yangtze River.
Key words: fish species with drifting eggs; early-stage fish resources; fish egg runoff; spawning ground; longitudinal distribution; the upper Yangtze River
