西北太平洋海域太平洋褶柔鱼秋生群产卵场指数动态评估❋
2022-02-24夏欣梅田永军刘世刚李建超马舒扬
夏欣梅, 田永军, 刘 阳❋❋, 刘世刚, 李建超, 马舒扬
(1.中国海洋大学水产学院渔业海洋学实验室, 山东 青岛 266003; 2. 自然资源部第三海洋研究所, 福建 厦门 361000)
太平洋褶柔鱼(Todarodespacificus)主要分布于西北太平洋,特别是日本海、中国的东海及黄海,是中国、日本和韩国的重要商业捕捞鱼种[1-2]。太平洋褶柔鱼寿命1年,根据产卵时间的不同分为夏生群、秋生群和冬生群[3],渔船主要捕捞秋生群和冬生群,其中秋生群于每年9—12月在东海北部和日本海西南部产卵[4-5],孵化后向北洄游至日本海的索饵场索饵[4,6]。日本自1979年起记录了秋生群的渔获量和单位捕捞努力量渔获量(Catch per unit effort,CPUE)数据(见图1),近年来,太平洋褶柔鱼秋生群的渔获量和CPUE不断下降,2018年渔获量降至24 247 t,这是自1986年以来的最低渔获量。
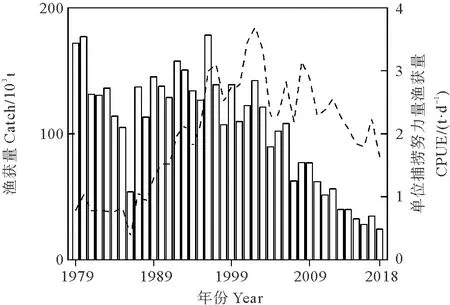
(空白条带为渔获量,黑色虚线为CPUE。Blank bar represents catch and black dashed line represents CPUE.)
目前对太平洋褶柔鱼的研究认为其资源丰度主要受海洋环境条件的影响。Sakurai等[3]认为太平洋褶柔鱼的资源丰度很大程度取决于产卵和补充过程,而海表温度(Sea surface temperature,SST)是影响太平洋褶柔鱼产卵的重要因子[3,7-12]。唐峰华等[13]认为太平洋褶柔鱼渔场的最适SST为16~18 ℃、最适叶绿素a(Chlorophylla,Chla)浓度为0.37~0.45 mg/m3。吴梦瑶等[14]利用剩余产量模型评估太平洋褶柔鱼秋生群资源,发现当SST在18~23 ℃时,渔获量会随温度升高而降低。张硕等[15]利用SST建立误差反向传播(Error backpropagation,BP)人工神经网络(Artificial neural networks,ANN)模型预测太平洋褶柔鱼冬生群资源丰度。产卵场指数可用于评估产卵场环境适宜性并分析海洋鱼类资源丰度的变化[16-17],如SST、适宜产卵场面积(Suitable spawning area,SSA)、Chla浓度等。但目前关于长时间尺度下太平洋褶柔鱼资源丰度变化的研究还很少,因此,本文基于1979—2018年潜在产卵场开发了产卵场指数,分析产卵场动态与太平洋褶柔鱼秋生群资源丰度的关系,为将来管理和评估太平洋褶柔鱼资源提供科学依据。
1 数据和方法
1.1 研究数据
以往的研究通常使用渔获量或CPUE作为资源丰度指标,故本研究使用的资源丰度指标为渔获量与CPUE[2-3,16],数据来自日本水产综合研究中心(Fishe-ries research agency,FRA)《平成30(2018)年太平洋褶柔鱼秋生群资源评估报告》[18],时间范围为1979—2018年,渔获量的单位为t,CPUE的单位为t/d。
环境数据包括海表温度和水深,数据范围覆盖太平洋褶柔鱼秋生群的产卵场21°N—40°N,121°E—142°E(见图2)[5]。其中SST数据来源于海洋水色数据库(https:∥oceancolor.gsfc.nasa.gov/)和美国国家海洋和大气管理局(https:∥psl.noaa.gov/research/),时间分辨率为月,水深数据来源于ETOPO1全球地形模型(https:∥www.ngdc.noaa.gov/)。为了保证数据在空间分辨率上的一致性,将所有环境数据重采样至1°×1°。

图2 西北太平洋海域太平洋褶柔鱼秋生群分布区域
1.2 产卵场指数
利用适宜太平洋褶柔鱼产卵的SST范围和水深计算了1979—2018年产卵场的年变化。以往的研究表明,最适宜太平洋褶柔鱼产卵的SST为18~24 ℃,在此范围内太平洋褶柔鱼幼体的存活率和游泳能力较高[9],繁殖假说认为太平洋褶柔鱼通常在水深-500~-100 m的大陆架和大陆坡上产卵[3,5,7,19-20]。因此,本论文将适宜产卵的SST范围设定为18~24 ℃,水深范围设定为-500~-100 m,40年潜在产卵场绘图均采用ArcMap 10.5实现。
为了分析产卵场环境条件对太平洋褶柔鱼秋生群资源丰度的影响,本研究使用了3种产卵场指数:适宜海表温度均值(Mean value of suitability sea surface temperature,MVSS)、适宜性SST加权面积(Suitability SST weighted area,SSWA)和等温线经向位置(Meridional position of SST isoline,MP)。
太平洋褶柔鱼秋生群的产卵期为9—12月[3,5],故本研究中的MVSS使用潜在产卵场9—12月的平均SST计算。SSWA用来描述潜在产卵场的时空变化,考虑了SST对太平洋褶柔鱼产卵和存活率的不同影响,利用如下表达式计算第i年的SSWA[16]:
(1)
式中:Ni表示第i年产卵场中18~24 ℃的网格数;TSIj表示网格j的温度适宜性指数(Temperature suitability index,TSI);Sj表示将网格j固定为1°×1°(经度×纬度)时的大小。
(2)
式中:μ表示平均SST为21 ℃;σ表示方差为1.5。
适宜产卵的SST范围为18~24 ℃,因此我们采用适宜产卵的临界等温线的平均纬度反映潜在产卵场总体位置的变化,即18 ℃等温线经向位置(Meridional position of 18 ℃ SST isoline,MP18)和24 ℃等温线经向位置(Meridional position of 24 ℃ SST isoline,MP24),18和24 ℃等温线平均纬度的提取和3种产卵场指数构建均采用MATLAB R2018a实现。
1.3 统计分析
贯序跃变检测(Sequential regime shift detection,SRSD)被用来分析长时间尺度下资源丰度和产卵场指数的变化情况[21-22]。
首先,根据t检验确定具有显著差异的两组连续数据平均值的差值(Difference,diff)[22]:
(3)
。
(4)

(5)
式中:Xi为第i年的变量;m为出现跃变后至第m年的时间长度[22]。RSI=0时表示未发生跃变,RSI>0时表示发生正向跃变,RSI<0时表示发生负向跃变。
最后,未发生跃变发生时连续数据的平均值使用Mean表示:
(6)
本研究中设定参数显著性检验置信水平p=0.1、转换截至长度l=10和异常值处置Huber权重参数h=1,时间尺度均为1979—2018年。
为了进一步量化产卵场指数与太平洋褶柔鱼秋生群资源丰度之间的关系,本研究采用了相关分析和梯度森林法。梯度森林是一种机器学习方法,建立在随机森林上[23],旨在确定预测变量和响应变量之间的复杂关系,并提供响应变量的平均R2加权重要性和累计重要性结果。在本研究中,梯度森林被用来分析各种产卵场指数的相对重要性,梯度森林模型构建和绘图使用R软件中的gradientForest包实现。
2 结果
2.1 资源丰度的变化
用SRSD方法检测太平洋褶柔鱼秋生群资源丰度1979—2018年的变化如图3所示。结果显示渔获量在2003—2004年和2012—2013年发生2次跃变,对应RSI值分别为-1.4和-0.8,即2次跃变都呈下降态势。在2004年发生跃变前,渔获量平均值为132.5×103t,2004年发生跃变后渔获量平均值下降至75.0×103t,2013年后渔获量均值锐减为33.1×103t。
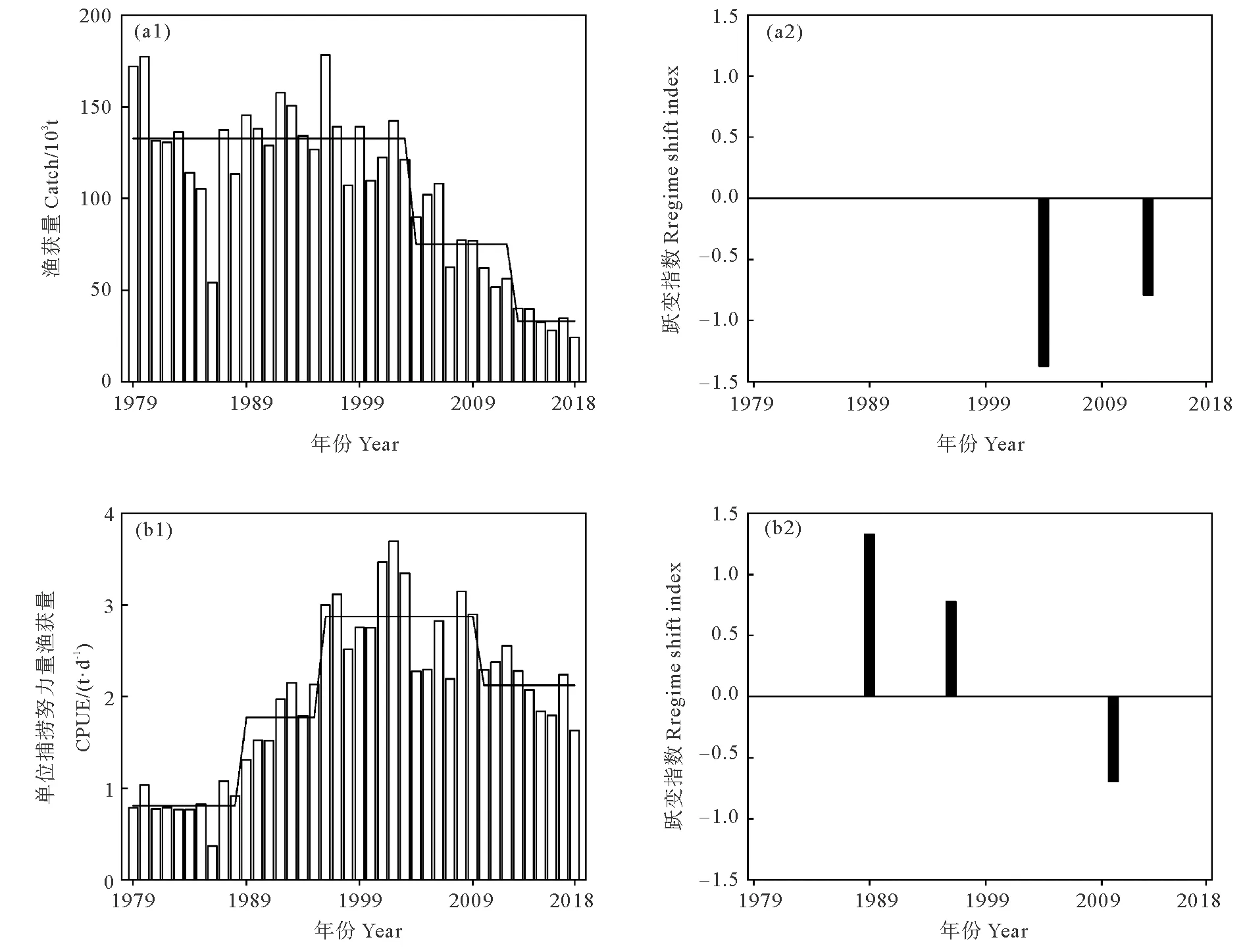
((a1)渔获量;(a2)渔获量跃变指数;(b1)单位捕捞努力量渔获量;(b2)单位捕捞努力量渔获量跃变指数。空白条带为资源丰度,黑色实线为平均值,黑色条带为跃变指数。(a1) Catch;(a2) Catch RSI;(b1) CPUE;(b2) CPUE RSI. The blank bar is abundance, the black solid line is mean, and black bar is regime shifts index.)
CPUE在1988—1989年、1995—1996年、2009—2010年发生3次跃变,对应RSI值分别为1.3、0.8和-0.7,即1988—1989年和1995—1996年跃变呈上升态势,而2009—2010年跃变呈下降态势。在1989年发生跃变前,CPUE保持在平均0.8 t/d,1989年后上升至1.8 t/d,1996年后进一步增加,平均为2.9 t/d,2010年后开始减少,平均为2.1 t/d。
综上可见,1989年发生跃变后,太平洋褶柔鱼秋生群资源丰度呈上升趋势,甚至在1996年渔获量高达178.3×103t,2002年CPUE高达3.7 t/d。2004年跃变后,太平褶柔鱼资源丰度呈下降趋势,特别是2018年渔获量锐减至24.2×103t,仅为1996年的13.6%。
2.2 潜在产卵场的变化特性
利用适宜太平洋褶柔鱼产卵的SST范围和水深计算了1979—2018年产卵场的年变化,适宜的SST范围设定为18~24 ℃,水深范围设定为-500~-100 m,图4显示40年潜在产卵场主要从台湾北部,经对马海峡一直向东北延伸到日本本州岛的近岸海域。

(彩色区域为潜在产卵场,灰色区域为陆地。The colored area is potential spawning ground and gray area is the land.)
1979—1998年潜在产卵场时空分布如图5所示,总体上产卵场位于东海北部到日本海西南部。图5中1979、1980、1982、1986、1988、1990—1992年向西南延伸至122°E,27°N附近,1989年以后日本海西南部产卵场区域略有增加,除1989、1994、1996、1979—1998年太平洋日本沿岸侧均有一小块产卵场分布。
1999—2018年潜在产卵场时空分布如图6所示,总体上产卵场位于东海北部到日本海西南部。图6中2005—2009年、2011、2015年明显向西南延伸至台湾北部,与图5相比总体上2002年以后太平洋日本沿岸的产卵场范围略有扩大,但对马海峡附近的产卵场区域明显缩小。

(黑色区域为潜在产卵场,灰色区域为陆地,白色区域为海洋。The black area is potential spawning ground, gray area is the land, and the white area is ocean.)

(黑色区域为潜在产卵场,灰色区域为陆地,白色区域为海洋。The black area is potential spawning ground, gray area is the land, and the white area is ocean.)
2.3 产卵场指数的变化
用SRSD方法检测太平洋褶柔鱼秋生群产卵场指数1979—2018年的变化如图7所示。
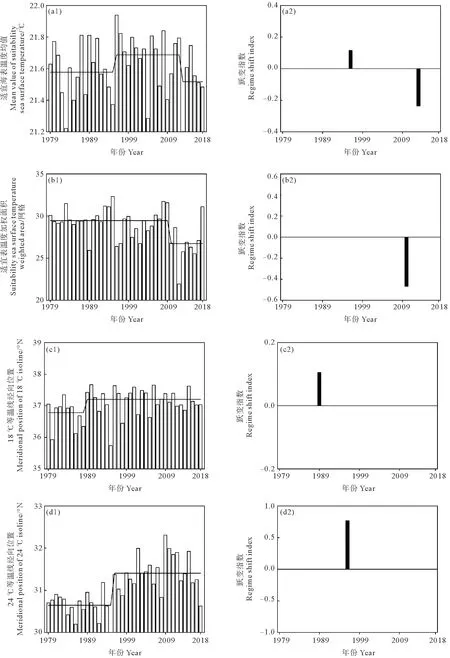
(空白条带为产卵场指数,黑色实线为平均值,黑色条带为跃变指数。The blank bar is spawning ground index, the black solid line is mean, and black bar is regime shifts index.)
结果显示MVSS在1995—1996年、2012—2013年发生2次跃变,对应的RSI值分别为0.1和-0.2,表明MVSS在1995—1996年呈上升态势,2012—2013年呈下降态势。在1996年发生跃变前,MVSS均值为21.58 ℃,1996年发生跃变后MVSS小幅增加至平均21.69 ℃,直至2013年降至平均21.52 ℃。总体上,MVSS在1996年达到最大值21.9 ℃,2013年达到最小值21.2 ℃。
SSWA在2009—2010年发生了1次跃变,RSI值为-0.5,表明SSWA跃变呈下降态势。在2010年发生跃变前,SSWA均值为29.5网格,2010年以后SSWA骤减至26.7网格。总体上,SSWA在1995年达到最大值32.3网格,2012年达到最小值21.9网格。
MP18与MP24分别在1988—1989年与1995—1996年发生2次跃变,对应的RSI值分别为0.1和0.8,表明2次跃变均呈上升态势。在1989年发生跃变前,MP18均值为36.8°N,1989年后北移至37.2°N。在1996年发生跃变前,MP24均值为30.6°N,1996年后均值北移至31.4°N。总体上,MP18最南为1995年达到35.7°N,最北为1990年37.6°N,MP24最南为1995年达到29.8°N,最北为2009年达到32.3°N。
1979—2018年产卵场指数发生上升态势跃变的时间为1988—1989年和1995—1996年,发生下降态势的时间为2009—2010年和2012—2013年,这4次时间节点和跃变态势均与资源丰度发生跃变的情况相吻合。
2.4 影响产卵场变动的环境因子
相关性分析结果如表1所示,MP24与MVSS显著相关(P<0.05),与SSWA和MP18呈极显著相关(P<0.01)。梯度森林被用来分析产卵指数及其1年滞后期对太平洋褶柔鱼秋生群资源丰度的相对重要性,排序结果如图8所示。模型确定MP24_Lag1是最重要的产卵场指数,其次是MP24、MVSS、SSWA_Lag1、MP18、MVSS_Lag1、SSWA、MP18_Lag1。为了进一步提高模型性能,将R2<0.02的产卵场指数SSWA和MP18_Lag1排除在累积重要性分析之外。
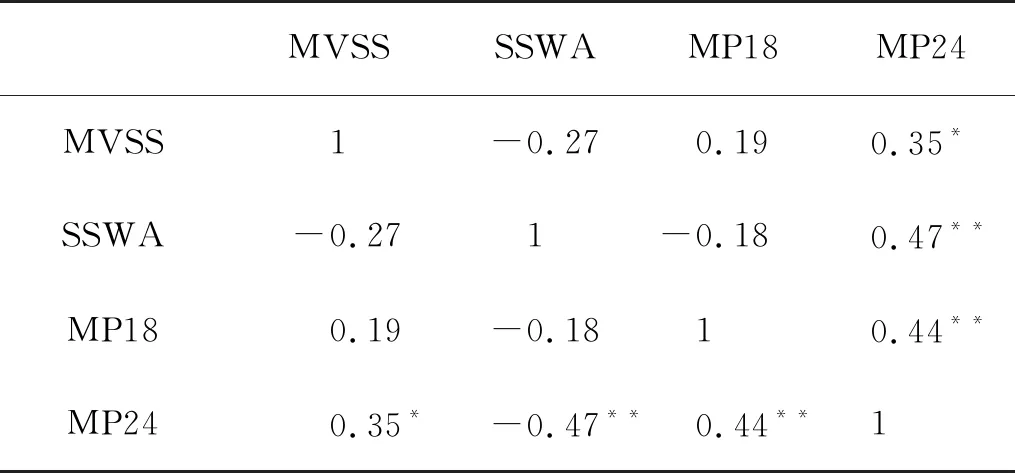
表1 1979—2018年西北太平洋海域太平洋褶柔鱼秋生群各产卵场指数的相关性矩阵

(MVSS为适宜海表温度均值,SSWA为适宜性海表温度加权面积,MP18为18 ℃等温线经向指数,MP24为24 ℃等温线经向指数。MVSS is mean value of suitability sea surface temperature(SST), SSWA is suitability SST weighted area and MP is meridional position of SST isoline. )
累积重要性可用来表示渔获量和CPUE对产卵场指数的敏感性。图9显示CPUE对产卵场指数MP24_Lag1、MP24、MP18、MVSS_Lag1的变化比渔获量敏感,二者对MVSS的敏感性相近,渔获量仅对SSWA_Lag1的变化比CPUE敏感。
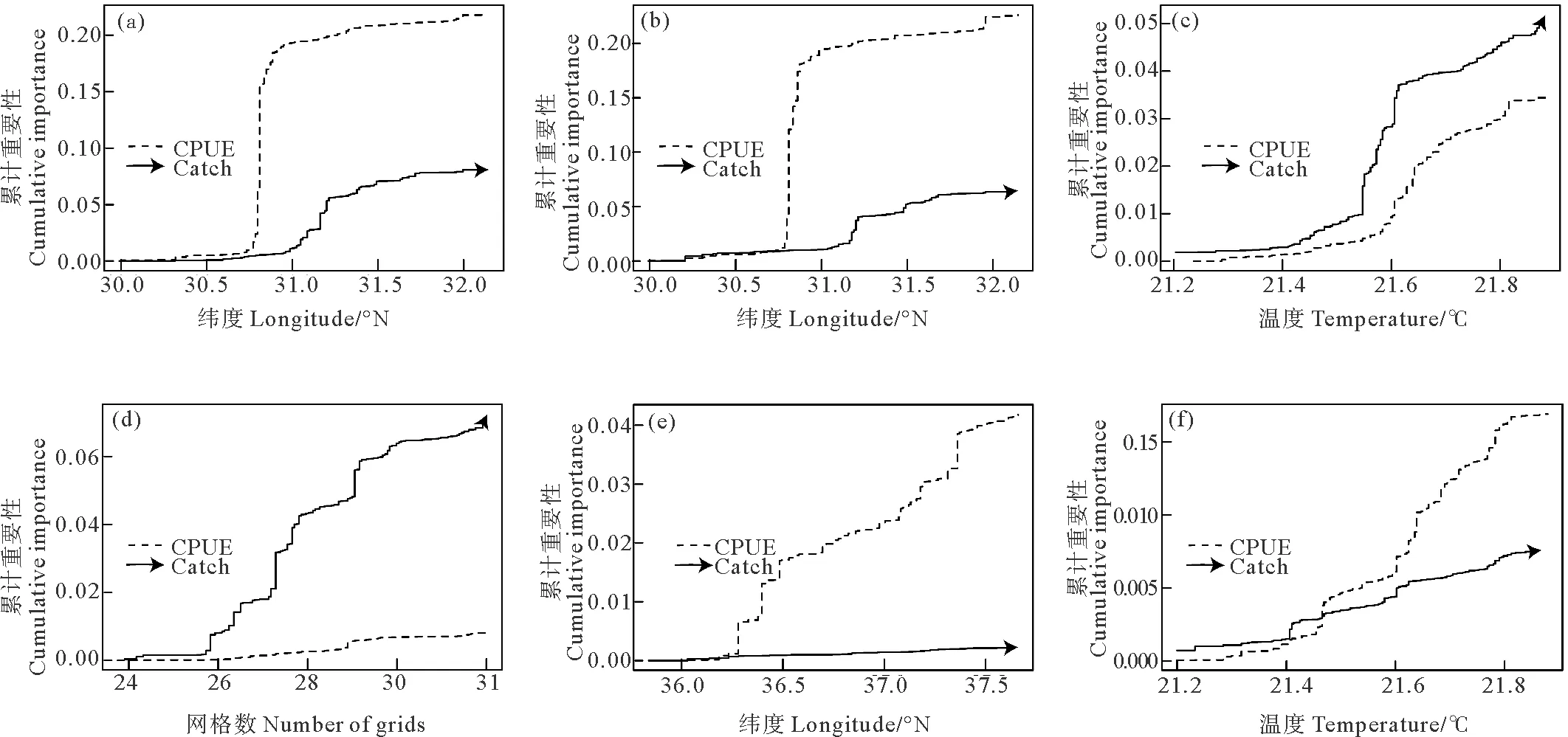
((a)MP24滞后一年;(b)24℃等温线经向位置;(c)适宜SST均值;(d)SSWA滞后一年;(e)18℃等温线经向位置;(f)MVSS滞后一年。黑色虚线为CPUE;黑色箭头线为渔获量。(a)Meridional position of 24 ℃ SST isoline with a 1-year lag;(b)Meridional position of 24 ℃ SST isoline;(c) Mean value of suitability sea surface temperature;(d)Suitability sea surface temperature weighted area with a 1-year lag;(e)Meridional position of 18 ℃ SST isoline;(f)Mean value of suitability sea surface temperature with a 1-year lag.Black dashed line is CPUE and black arrow line is catch.)
3 讨论
3.1 产卵场动态对太平洋褶柔鱼秋生群资源丰度的影响
本研究利用适宜太平洋褶柔鱼产卵的SST范围和水深计算了1979—2018年产卵场的年变化,而实际上影响其产卵场的环境因子很多,仅根据这2个参数会导致预测的潜在产卵场与实际存在一定的偏差,未来可以加入更多的环境因子提高预测的准确性。本研究中,产卵场指数共同反映了秋生群产卵场的变化。产卵场的环境条件决定了太平洋褶柔鱼产卵量和补充量的大小,由于其寿命仅1年,资源丰度很大程度取决于产卵量和补充量[3]。影响太平洋褶柔鱼资源丰度的环境因子很多,如Chla浓度、涡流动能(Eddy kinetic energy,EKE)和海面高度距平值(Sea surface height anomaly,SSHA)等[13,24],一般认为SST起着非常重要的作用[7,8,10-12]。1979—2018年产卵场MVSS与太平洋褶柔鱼秋生群资源丰度在跃变上表现出良好的对应关系,1995—1996年和2012—2013年同时发生了跃变,且两次变动态势相反,MVSS与资源丰度在1995—1996年同步增加,2012—2013年后同步减少。以前的研究认为升高的海水温度不仅有利于浮游植物的生长[25-26],而且能够增加浮游动物的种类[27],从而保证太平洋褶柔鱼的食物来源以增加补充量[1]。因此推测产卵场MVSS的跃变会导致太平洋褶柔鱼补充量的增减,从而导致资源丰度出现了相应的跃变。仅用MVSS无法解释资源丰度在1988—1989年、2009—2010年出现的跃变,于是本研究使用了产卵场指数SSWA与MP作进一步分析。
以往的研究认为产卵场的面积和空间分布可能会影响太平洋褶柔鱼冬生群资源丰度[2,3,5],SSWA被用来表示潜在产卵场的时空变化,能够解释SST变化对太平洋褶柔鱼产卵和存活的影响[16]。图3和图7显示SSWA与资源丰度在2009—2010年表现出相同的跃变均呈下降态势,同步跃变说明SSWA的减少对资源丰度具有不利的影响,以往的研究也认为最适产卵场面积的减少会导致资源丰度的降低[28]。
根据适宜产卵的临界温度18和24 ℃度等温线纬度计算的MP指数可以表示潜在产卵场整体位置的变化。MP18、MP24与资源丰度在1988—1989年和1995—1996年呈现相同的跃变且均呈上升态势,同步的跃变说明MP指数的升高对太平洋褶柔鱼的资源丰度具有正的作用。主要原因可能是太平洋褶柔鱼在产卵孵化后向北洄游进入日本海索饵[1],MP指数的升高表明潜在产卵场北移,使其更易于洄游到索饵场免受不利因素的干扰[12],从而使资源丰度增加。
梯度森林将MP24_Lag1定义为最重要的因子,指数包括1年滞后期是合理的,因为太平洋褶柔鱼的寿命为1年[3]。累积重要性分析中,除SSWA_Lag1,CPUE总体上对产卵场指数的变化比渔获量更为敏感,可能是因为CPUE是比渔获量更加适宜的资源丰度指数[3]。表1相关性分析表示,MP指数与MVSS和SSWA均显著相关。总之,3种产卵场指数共同解释了1979—2018年太平洋秋生群资源丰度的变化,特别是近年来资源丰度的不断下降。
3.2 太平洋褶柔鱼秋生群资源丰度的长期变化过程
太平洋褶柔鱼是高度依赖环境条件的机会主义物种,其资源丰度对海洋环境变化非常敏感[29]。潜在产卵场的MVSS不能完全解释太平洋褶柔鱼资源丰度的变化,故本研究构建了3种产卵场指数共同解释。基于SRSD分析,本研究构建的3种产卵场指数与资源丰度在1988—1989年、1995—1996年、2009—2010年、2012—2013年表现出相同的跃变,证明产卵场指数所反映的潜在产卵场的稳态变化与资源丰度有良好的对应关系。表1相关性分析表示,MP24与MVSS、SSWA、MP18均显著相关。基于以上分析,研究表明整个海域温度的变化导致了潜在产卵场位置的改变,从而影响潜在产卵场MVSS和SSWA,产卵场MVSS和SSWA的改变影响了产卵和存活的条件,从而导致补充量的变化,最终影响太平洋褶柔鱼资源丰度。资源丰度在1988—1989年和1995—1996年增加是因为产卵场指数发生上升态势的跃变,而2009—2010年和2012—2013年减少是因为产卵场指数发生下降态势的跃变,研究结果可以为解释太平洋褶柔鱼秋生群资源丰度的长期变化机理提供理论支撑。
4 结语
本文利用海表温度和水深开发了3种太平洋褶柔鱼秋生群产卵场指数,并分析了1979—2018年潜在产卵场动态变化,解析了太平洋褶柔鱼秋生群资源丰度的长期变动机制,证明了资源丰度和产卵场指数在1988—1989年、1995—1996年、2009—2010年、2012—2013年表现出同步的跃变,发现了太平洋褶柔鱼秋生群资源丰度的变化是由潜在产卵场环境条件是决定的。但本研究方法尚未解出2003—2004年资源丰度急剧减少的原因,未来可以加入更多的环境因子,如海表盐度、海流、Chla浓度、混合层深度等参数,用于综合探索太平洋褶柔鱼资源丰度变化机制。
致谢:感谢中国海洋大学水产学院渔业海洋学实验室诸位老师和同学提供的帮助。
