阿拉善荒漠啮齿动物优势种觅食活动对不同放牧方式的响应
2023-12-22刘丽娜袁帅李鑫付和平武晓东
刘丽娜 袁帅* 李鑫 付和平 武晓东
(1 内蒙古农业大学草原与资源环境学院,呼和浩特 010011) (2 草地啮齿动物生态学与鼠害控制自治区高等学校重点实验室,呼和浩特 010011) (3 草地资源教育部重点实验室,呼和浩特 010011)
在啮齿动物群落中,同域共存个体常通过调节空间、时间和食物分配来合理共享同一资源,以缓减潜在竞争从而实现共存 (Moriet al., 2019;Vivianoet al., 2022)。相关潜在竞争归根结底都与资源利用密切关联,其中食物是最重要的资源之一 (Brownet al., 1994)。觅食行为是动物获取食物资源,维持生存和自身正常代谢的一种基本活动(Moriet al., 2019)。啮齿动物的觅食行为包括选取觅食地点、搜寻食物来源、获取食物和处理食物等一系列行为。在不同生境中,动物在发现食物时一般会面临两种选择:原地取食,或者将食物搬运到其他地方贮藏以备后期取食 (Wanget al.,2013;郎政伟和巩合德,2016)。相关分配策略是各种因素综合作用的表现,也是动物长期适应生存环境自然选择的结果,动物在这些行为上所投入的时间和能量很大程度影响着动物的生存和繁殖,其由自身调控和外在因素作用共同决定 (尚玉昌,2006;张聪敏等,2019)。
而人类放牧活动是相对于自然生态环境的一种干扰,目前在荒漠生态系统中也最为常见 (陈利顶和傅伯杰,2000)。不同放牧干扰是导致植物群落和动物群落变化的主因,生境中食物资源和时空特异性会随着时间推移和干扰的不同而发生变化 (Carusoet al., 2016; Sévêqueet al., 2021)。阿拉善荒漠是我国的典型温性荒漠,生态系统结构简单,功能脆弱,对人为干扰反应较敏感 (付和平等,2007)。荒漠啮齿动物是该生态系统中的重要成员和环境变化的指示者 (Flowerdewet al., 2004)。三趾跳鼠 (Dipus sagitta)、五趾跳鼠 (Orientallactaga sibirica) 和子午沙鼠 (Merionesmeridianus) 是阿拉善荒漠区同域共存的3种夜行性优势鼠种,其中三趾跳鼠和五趾跳鼠均为两足跳跃且具有冬眠习性的啮齿动物;子午沙鼠擅长四足活动,常在巢区中心区觅食,无冬眠行为,具有储粮习性 (赵肯堂,1964;梁杰荣和肖运峰,1982;刘焕金等,1984)。有关同域分布荒漠啮齿动物在不同干扰下的觅食差异性及其对荒漠地区的生物多样性维持作用尚乏研究。
以往的研究表明,放牧干扰会对啮齿动物群落的自然分布、栖息地选择、行为节律、繁殖策略、种群数量和群落多样性等产生影响,且多集中在森林和高原景观系统 (Knight and Fitzner,1985;李俊生等,2005; Gaynoret al., 2018;李鑫等,2020)。但是有关温性荒漠中共存的啮齿动物物种对不同放牧干扰下的觅食行为响应的研究并不多见 (焦广强,2011;郎政伟和巩合德,2016;张彩军,2022;林天杰,2022)。
为此,本文通过红外相机陷阱法,以阿拉善盟荒漠区的夜行性优势鼠种为研究对象,对其在不同放牧干扰生境下各行为的时间分配进行研究,探讨各优势鼠种的摄食策略对不同放牧干扰的响应,并确定其主导因素。从研究动物的觅食生态学特征着手,从摄食行为策略方面丰富动物共存理论,有助于进一步认识不同放牧方式下阿拉善荒漠啮齿动物的觅食行为差异性,掌握动物的摄食策略,能更好地理解动物对生存环境的适应策略。从鼠害防治角度来看,针对优势鼠种的摄食策略适时进行干预,可能会提高对其种群数量的控制效率,对有害啮齿动物的生物防治具有重要的意义,也为荒漠区制定合理的放牧管理制度提供理论科学依据。
1 研究方法
1.1 研究地概况
研究地位于内蒙古自治区阿拉善盟东南部的嘉尔嘎勒赛汉镇孪井滩生态移民示范区。海拔1 399 m,地理坐标为北纬 37°24′ ~ 38°25′,东经 104°10′ ~ 105°30′,是我国西北部典型的温带荒漠干旱脆弱生态系统。研究区为典型的高原大陆性气候,冬季严寒、干燥,夏季酷热,昼夜温差大;年降水量约45 ~ 215 mm,主要集中在7—8 月;年蒸发量约3 000 ~ 4 700 mm;当地植被稀疏、种类贫乏、结构简单、覆盖度极低 (约1% ~20%),其植物类型多为旱生、超旱生、盐生的灌木、小灌木、半灌木和小半灌木。灌木优势种包括:红砂 (Reaumuria songarica)、霸王 (Zygophyllum xanthoxylon)、白刺 (Nitraria tangutorum)、短脚锦鸡儿 (Caragana brachypoda)、猫头刺 (Oxytropis aciphylla) 和梭梭 (Haloxylon ammodendron);多年生优良禾本科牧草和豆科牧草较少,草本优势植物多为:糙隐子草 (Cleistogenes squarrosa)、刺蓬 (Salsola tragus)、条叶车前 (Plantago lessingii)和雾冰藜 (Bassia dasyphylla) 等。地形主要包括荒漠、沙漠、低山丘陵和戈壁等。
1.2 实验设计
本实验于2020 年在阿拉善荒漠研究区内,依据草地利用方式的不同选择3种不同干扰类型的生境,并在每类干扰生境中设置3块重复样地作为啮齿动物活动监测样地,每块样地的面积为1 hm2。不同干扰类型分别为禁牧区 (1997 年开始围封禁牧,植被盖度约23.0%)、过牧区 (放牧强度为每公顷3.75 ~ 4.23只羊,植被盖度约16.4%) 和轮牧区 (1995 年起,50 ~ 60 只成年羊在3 个划分小区,每1.5 个月轮替放牧,植被盖度为19.7%) (武晓东等,2016)。选取2020 年5 月、7 月和9 月红外相机观测数据和样地中植物、土壤数据和啮齿动物群落调查数据进行分析。
1.2.1 植物、土壤数据采集
在每个啮齿动物活动痕迹监测样地内随机选取3 个10 m × 10 m 的灌木样方,在每个灌木样方中随机选取3 个1 m × 1 m 的样方作为草本样方,分别测量样方内灌木、草本植物的植被“四度一量”。并同步使用土壤硬度仪 (TYD-2 数字式土壤硬度计) 对 [0, 5) cm、[5, 10) cm、[10, 15) cm、[15, 20] cm 4 个土层的土壤硬度进行测定,采用烘干法 (105 ℃, 12 h) 对4个土层的含水量进行测定。
1.2.2 动物识别与监测
预实验在每类生境的3个重复样区内各随机选取5个点以新鲜花生米作为饵料,次日检查饵料取食情况,每个样区选取1个饵料取食最多的点作为正式实验红外相机架设点,3 种干扰类型生境的9 个样区中共设置9 个红外相机架设点。每个点架设1 ~ 2 台相机 (型号:东方红鹰ERE-E1B)。在相机布设样地,将花生米饵料与过筛的沙土 (10 L)均匀混合放入食盘 (50 cm × 50 cm × 4 cm) 中,调整相机位置 (相机距离地面30 ~ 40 cm);调整镜头,避免逆光拍摄。固定完成后将相机参数统一为录像模式。每天18: 00 布置食盘,次日06: 00 对食盘中的内容物进行过筛称重,记录剩余饵料数并继续重新布置食盘,监测时间为24 h,每月连续观测4 d。
1.2.3 选取研究对象
2020 年5 月、7 月和9 月在阿拉善荒漠过牧、禁牧及轮牧3 种不同放牧处理下共获得红外相机视频数4 845 个,有效视频3 689 个。在所有有效视频中共观测到13 个物种,其中仓鼠科2 种,分别为小毛足鼠 (Phodopus roborovskii)、子午沙鼠;跳鼠科3 种,分别为蒙古羽尾跳鼠 (Stylodipus andrewsi)、三趾跳鼠、五趾跳鼠;兔形目1 种,为蒙古兔 (Lepus tolai)。在有效视频中共拍摄到三趾跳鼠1 760 次、五趾跳鼠1 155 次、子午沙鼠1 046 次。通过有效视频观测频次结果初步选取研究对象为阿拉善荒漠区同域共存的夜行性优势鼠种:三趾跳鼠、五趾跳鼠和子午沙鼠 (图1)。

图1 荒漠区啮齿动物优势种红外相机拍摄图. A:三趾跳鼠;B:五趾跳鼠;C:子午沙鼠Fig. 1 Photographed by infrared camera of the dominant rodent species in desert. A: Dipus sagitta; B: Orientallactaga sibirica; C: Meriones meridianus
1.2.4 行为识别
将获得的视频等数据按时间段进行分类储存,逐一查看和分析红外相机中的视频和照片数据,详细记录相机位点、视频编号、温度、触发日期和时间、个体数、GPS位点、行为类型等数据。
对行为的定义和描述是定量开展动物行为学研究的前提,对于已有研究记录的引用原定义,其他根据观察结果进行补充定义 (Boake, 1998; Ferrer-i-Cancho, 2013;车烨和李忠秋,2014;王涛涛等,2015) (表1)。

表1 啮齿动物行为分类Table 1 Classification of rodent behavior
1.3 数据处理
1.3.1 种群密度
统计2020 年5 月、7 月和9 月,每一鼠种在各不同放牧样地中出现频次,使用捕获率模型 (capture rate model, CRM) 计算不同优势鼠种在不同放牧下的种群平均密度 (Liuet al., 2013)。
式中Di为第i月啮齿动物种群平均密度 (ind./hm2);Nik表示第i月第k台相机所拍摄的某种啮齿动物总数;Sk代表第k台红外相机所拍摄范围 (本研究中相机拍摄范围均为2 m × 2 m);Tik表示第i月第k台红外相机的监测时间 (单位:d);n表示实验中参与监测啮齿动物的红外相机数量 (Liuet al., 2013;袁帅等,2021)。
选取2020 年5 月、7 月和9 月不同放牧方式下啮齿动物优势种的种群密度,采用IBM SPSS Statistics 25 软件的一般线性模型 (general linear model, GLM) 将不同物种作为固定因子 (fixed factor),3 种放牧方式下的种群密度作为响应变量 (dependent Variable, DV),月份作为随机因子 (random factor),比较在不同放牧方式下各啮齿动物种群密度的差异性。所有数据以平均数 ± 标准差(mean ± SD) 表示。利用Origin 2021 绘制种群密度图。
1.3.2 行为频次所占比例
统计啮齿动物5种主要行为的出现频次,以此来计算不同时空尺度下各种行为出现频次所占比例,计算公式如下:
式中P为某一时间段的出现频次,A为观测到目标行为的数目,N为观测天数,B为观测行为总数(赵运林等,2018)。将啮齿动物各行为频次导入IBM SPSS Statistics 25 软件,使用一般线性模型,将不同月份作为随机因子,分别将不同物种和不同放牧处理作为固定因子,并将5种行为作为响应变量,比较啮齿动物优势种各行为在不同放牧方式下的差异性。所有数据以平均数 ± 标准差 (mean ±SD) 表示,所有图使用Origin 2021进行绘制。
1.4 相关分析
对不同放牧方式下各啮齿动物种群密度以及植物、土壤等环境因子和啮齿动物行为做相关分析,使用Spearman 相关系数来衡量相关性大小,探究啮齿动物行为对不同放牧方式的响应。
Spearman相关系数表达式如下:
式中n为样方数,dk=xik-xjk,xik和xjk分别为种i与种j在样方k中的秩。*表示0.05 水平 (双侧) 显著相关,**表示0.01 水平 (双侧) 显著相关。使用Tbtools 软件对环境因子和种群密度结果进行可视化处理 (Chenet al., 2020)。
2 结果
2.1 不同放牧方式下荒漠区啮齿动物优势种种群密度
对不同放牧方式下的3种荒漠区优势啮齿动物的种群密度统计结果表明:过牧区,3 种啮齿动物的种群密度存在种间差异 (F2,24= 3.375,P=0.049),其中五趾跳鼠的种群密度显著高于子午沙鼠;禁牧区,3 种啮齿动物的种群密度存在种间差异 (F2,24= 3.245,P= 0.021),其中五趾跳鼠的种群密度显著低于三趾跳鼠;轮牧区,3 种啮齿动物的种群密度种间差异不显著 (F2,24= 0.750,P=0.529,图2)。

图2 荒漠区啮齿动物优势种种群密度. 不同字母表示不同物种种群密度在不同放牧方式下具有显著差异,显著水平P = 0.05Fig. 2 Population density of rodent dominant species in desert. Different letters indicate that the population density of different species has significant differences under different grazing patterns, and the significant level P = 0.05
2.2 不同放牧方式下荒漠区啮齿动物优势种行为频次比例
不同放牧方式对三趾跳鼠取食行为 (F2,109=2.352,P= 0.100)、观望行为 (F2,109= 0.085,P=0.919) 无显著影响;三趾跳鼠在禁牧区和过牧区的觅食行为所占比例显著高于轮牧区 (F2,109=7.315,P= 0.001);其躲避行为在禁牧区显著高于轮牧区 (F2,109= 3.500,P= 0.034);虽然3 种放牧方式下三趾跳鼠的贮食行为占比均较少,但是禁牧区三趾跳鼠的贮食行为显著高于过牧区和轮牧区(F2,109= 3.930,P= 0.022) (图3A)。

图3 荒漠区啮齿动物优势种三趾跳鼠 (A)、五趾跳鼠 (B)、子午沙鼠 (C) 在不同放牧方式下的行为频次比例. 不同字母表示啮齿动物优势种的行为占比具有显著差异,显著水平P = 0.05Fig. 3 Proportion of behavior frequency of Dipus sagitta (A), Orientallactaga sibirica (B), and Meriones meridianus (C) in desert under different grazing patterns. Different letters indicated that the proportion of rodent species behaviors were significantly different, and the significant level was P = 0.05
五趾跳鼠的觅食行为 (F2,109= 17.918,P=0.010)、取食行为 (F2,109= 3.800,P< 0.001)、躲避行为 (F2,109= 5.886,P= 0.040) 在禁牧区显著低于过牧区和轮牧区,这3种行为所占比例均在过牧区最高;其贮食行为 (F2,109= 0.962,P= 0.455) 和观望行为 (F2,109= 2.690,P= 0.182) 在3种放牧方式下无显著差异 (图3B)。
子午沙鼠的取食行为在3种放牧方式下占比均较低且无显著差异 (F2,109= 0.972,P= 0.453);子午沙鼠的其他行为均受放牧方式显著影响,在过牧区的觅食行为(F2,109= 10.101,P= 0.027)、贮食行为 (F2,109= 9.421,P= 0.031)、观望行为 (F2,109=6.774,P= 0.041) 和躲避行为 (F2,109= 10.325,P=0.026) 均显著低于禁牧区和轮牧区 (图3C)。
2.3 不同放牧方式下荒漠区啮齿动物优势种行为活动分配比例
在过牧区,五趾跳鼠的觅食行为 (F2,108=12.245,P= 0.020) 和取食行为比例 (F2,108=45.583,P< 0.001) 显著高于三趾跳鼠和子午沙鼠,其中子午沙鼠的这些行为所占比例均最低;五趾跳鼠和三趾跳鼠的躲避行为比例显著高于子午沙鼠 (F2,108= 7.067,P= 0.046);3 种啮齿动物的贮食行为 (F2,108= 0.973,P= 0.452) 和观望行为(F2,108= 3.104,P= 0.153) 占比较低且无显著差异(图4A)。
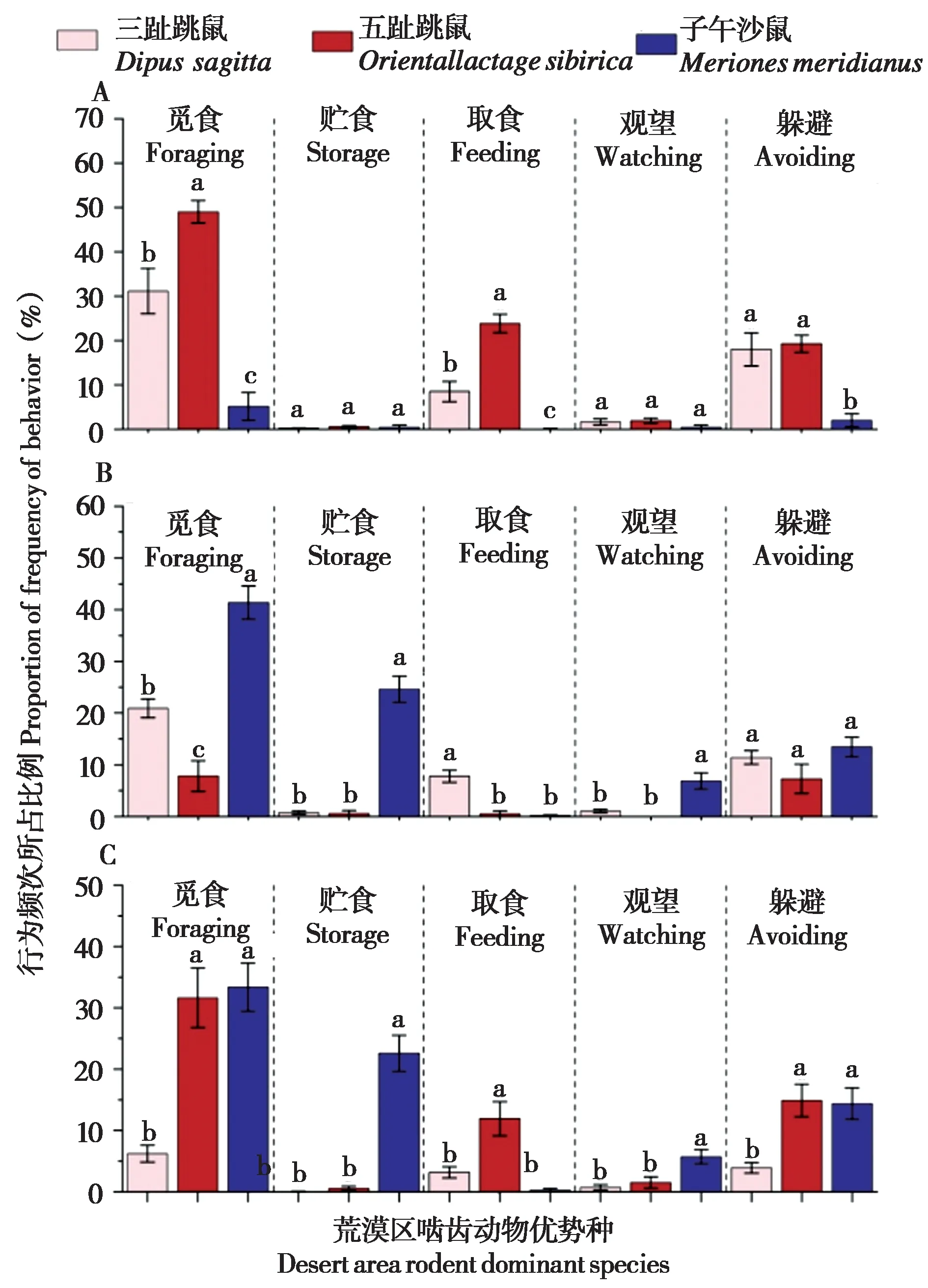
图4 过牧区 (A)、禁牧区 (B) 和轮牧区 (C) 啮齿动物优势种活动时间分配. 不同字母表示啮齿动物优势种的行为占比具有显著差异,显著水平P = 0.05Fig. 4 Time allocation of rodent dominant species in overgrazing(A), prohibited grazing (B), and rotational grazing areas (C). Different letters indicated that the proportion of rodent species behaviors were significantly different, and the significant level P = 0.05
在禁牧区,子午沙鼠的觅食行为 (F2,108=24.638,P= 0.006)、贮食行为 (F2,108= 13.895,P=0.016) 和观望行为比例 (F2,108= 6.141,P= 0.006)均显著高于三趾跳鼠和五趾跳鼠;三趾跳鼠的取食行为比例显著高于五趾跳鼠和子午沙鼠 (F2,108=14.876,P= 0.014)。3 种啮齿动物躲避行为占比无显著差异 (F2,108= 3.269,P= 0.144) (图4B)。
在轮牧区,子午沙鼠的贮食行为 (F2,111=31.593,P= 0.004) 和观望行为比例 (F2,111=25.657,P= 0.005) 显著高于三趾跳鼠和五趾跳鼠;三趾跳鼠的觅食行为 (F2,111= 16.96,P< 0.001) 和躲避行为比例 (F2,111= 31.209,P= 0.003) 显著低于五趾跳鼠和子午沙鼠,五趾跳鼠的取食行为比例显著高于三趾跳鼠和子午沙鼠 (F2,111= 12.583,P<0.001) (图4C)。
2.4 不同放牧方式下环境特征描述
不同放牧方式对草本和灌木的不同指标产生了显著影响 (表2)。过度放牧显著降低了草本和灌木的高度 (草本:F2,24= 5.770,P= 0.009;灌木:F2,24= 34.880,P< 0.001)、盖度 (F2,24= 6.380,P=0.006) 和生物量 (F2,24= 5.170,P= 0.014)。禁牧显著降低了草本的盖度 (F2,24= 5.190,P= 0.013)。
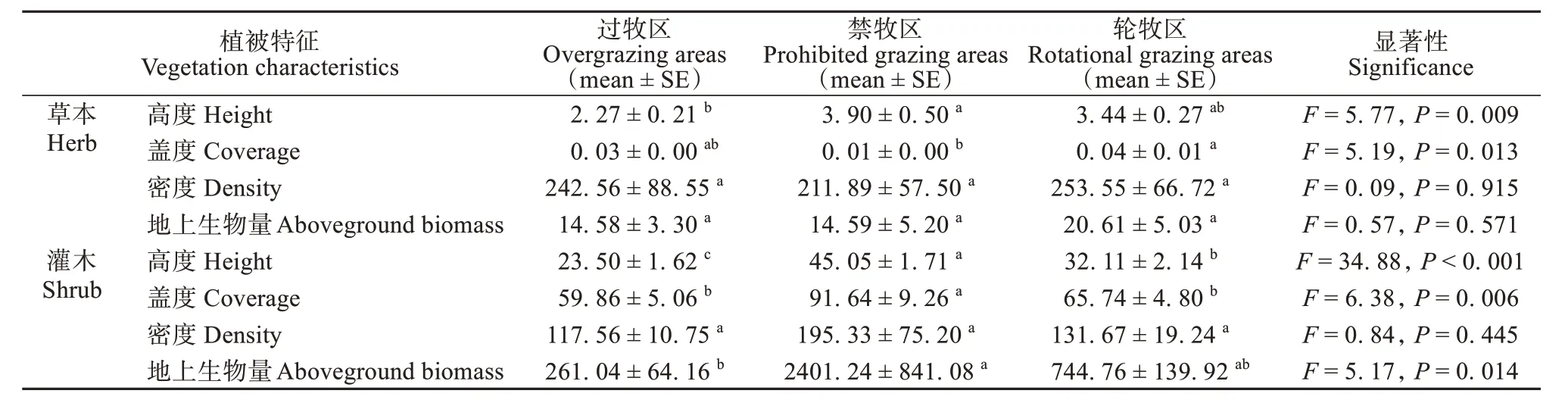
表2 植被因子指标差异性分析Table 2 Differential analysis of vegetation factor indicators
不同放牧方式对土层硬度的影响主要表现为过牧和轮牧同禁牧相比,显著增加了0 ~ 5 cm 土层硬度 (F2,24= 4.758,P= 0.011)。放牧方式显著改变了0 ~ 5 cm 土层含水量 (F2,24= 0.266,P= 0.018),其中过牧区0 ~ 5 cm 土层含水量最小,轮牧区含水量最大 (表3)。
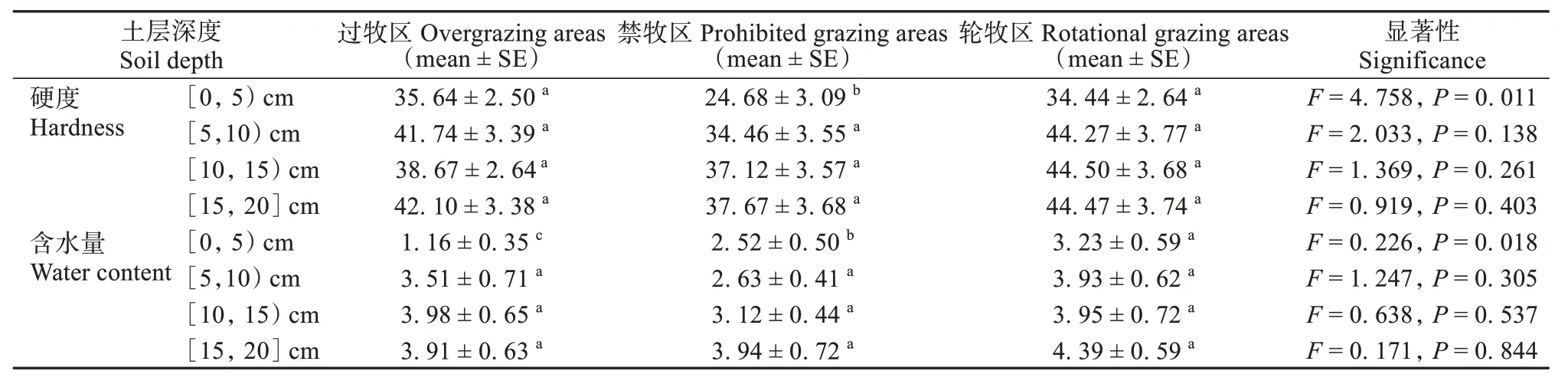
表3 过牧区、禁牧区和轮牧区不同土层土壤含水量、土壤硬度差异性分析Table 3 Analysis of soil water content and soil hardness differences in different soil layers in overgrazing areas, porhibited grazing areas and rotational grazing areas
2.5 不同放牧方式下优势鼠种行为的影响因素
过牧干扰下,荒漠区3种啮齿动物优势种的行为与种群密度的相关分析表明:过牧区三趾跳鼠的5 种行为均与其种群密度存在正相关关系;三趾跳鼠贮食行为随着其他两种啮齿动物种群密度增多而增多,三趾跳鼠的躲避行为与五趾跳鼠种群密度呈正相关;五趾跳鼠的贮食行为和观望行为随着三趾跳鼠种群密度的增多而增多,五趾跳鼠的觅食行为和躲避行为与其种群密度呈显著正相关;子午沙鼠取食行为与过牧区五趾跳鼠种群密度呈显著正相关;子午沙鼠躲避行为与三趾跳鼠和五趾跳鼠种群密度呈显著正相关 (图5)。综上,在过牧区啮齿动物的种内和种间个体密度对三趾跳鼠的行为影响较大,对子午沙鼠行为影响较小。
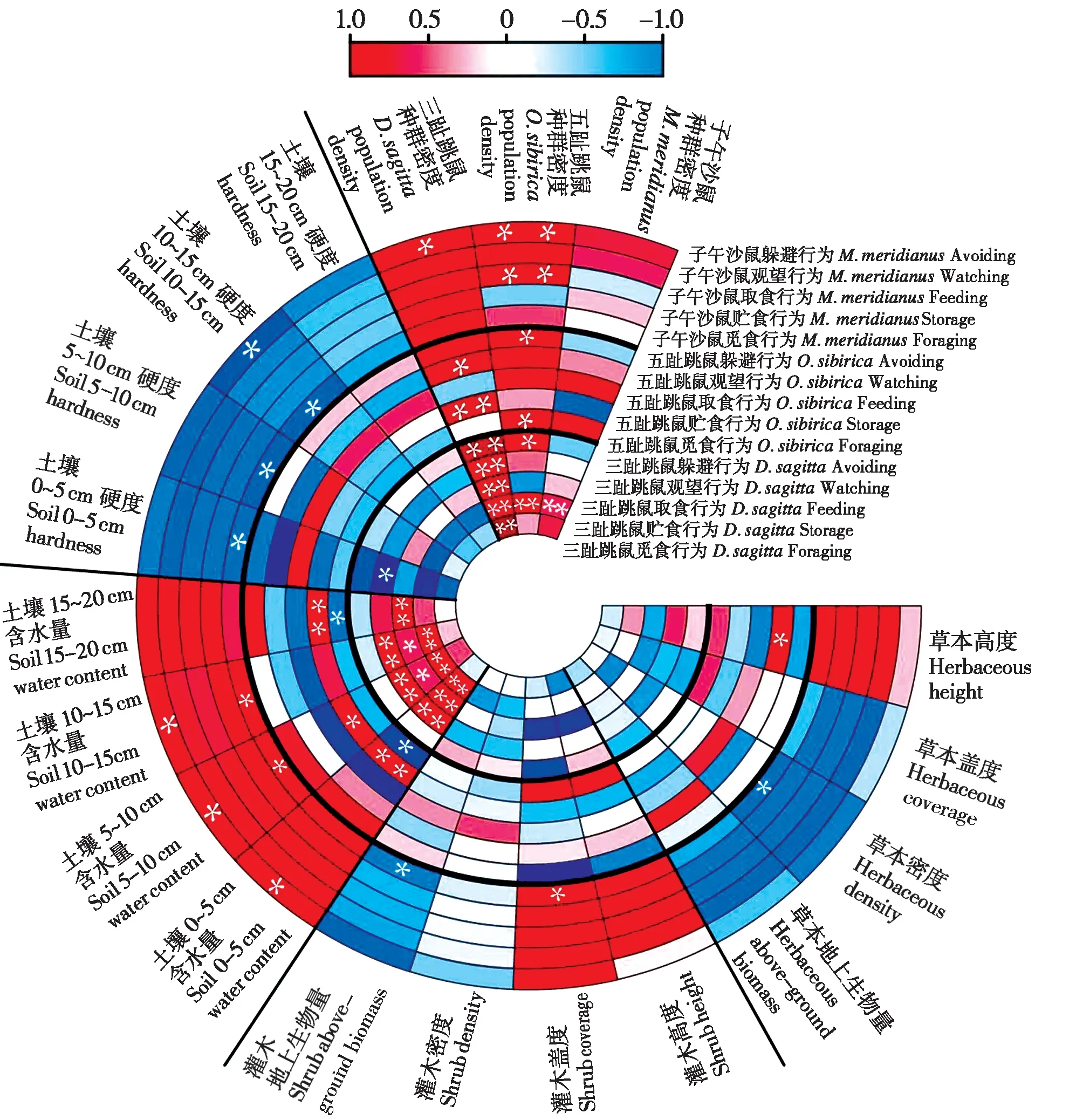
图5 过牧区环境因子与啮齿动物优势种行为频次相关性. 图中横坐标表示不同鼠种行为;纵坐标为影响因子,不同颜色反应Sperman 相关性,红色代表正相关关系,蓝色代表负相关关系,颜色越深相关性越强. **表示在 0.01 级别 (双尾) 相关性显著,* 表示在 0.05 级别 (双尾) 相关性显著Fig. 5 Spearman correlation of environmental factors with behavioral frequency of rodent dominant species in overgrazing areas. The abscissa shows rodent species behaviors. The ordinates show effect factors in the overgrazing areas. Red represents a positive correlation,and blue represents a negative correlation. The darker the color, the stronger the relationship. ** indicates significant correlation at level 0.01 (two-tailed test), * indicates significant correlation at level 0.05(two-tailed test)
过牧干扰下,荒漠区3种啮齿动物优势种行为与环境因子的相关分析表明:过牧区显著影响三趾跳鼠的外部环境因子主要包括土壤含水量和0 ~5 cm 土层的硬度,这些因子主要影响三趾跳鼠的贮食、取食和观望行为;影响五趾跳鼠行为的外部环境因子主要是土壤含水量和草本高度,其主要影响了五趾跳鼠的觅食、贮食和观望行为;影响子午沙鼠行为的外部环境因子较多,主要包括0 ~ 15 cm 土层的硬度、0 ~ 15 cm 土层的含水量、灌木生物量及盖度和草本密度,其主要影响子午沙鼠的觅食行为和躲避行为 (图5)。
禁牧干扰下,荒漠区3种啮齿动物优势种行为与种群密度的相关分析表明:禁牧区,三趾跳鼠仅贮食行为和观望行为与其种群密度呈显著正相关;五趾跳鼠的贮食行与三趾跳鼠的种群密度呈显著正相关;五趾跳鼠的所有行为均与自身种群数量成正比;子午沙鼠的觅食行为和躲避行为与三趾跳鼠的种群密度正相关 (图6)。
禁牧干扰下,荒漠区3种啮齿动物优势种行为与环境因子的相关分析表明:禁牧区,显著影响三趾跳鼠的外部环境因子较多,主要包括土壤含水量、灌木高度和10 ~ 15 cm 土层的硬度;禁牧区未发现显著影响五趾跳鼠行为的外部环境因子;影响子午沙鼠行为的外部环境因子较多,主要包括土壤含水量、灌木高度和草本密度,其主要影响子午沙鼠的贮食、觅食和观望行为 (图6)。
轮牧干扰下,荒漠区3种啮齿动物优势种行为与种群密度的相关分析表明:在轮牧区,三趾跳鼠的贮食、取食和观望行为与其种群密度呈显著正相关,其贮食行为还与五趾跳鼠种群密度呈显著正相关;五趾跳鼠仅贮食行为与三趾跳鼠种群密度呈显著正相关;子午沙鼠贮食行为与其自身种群密度呈显著正相关,与五趾跳鼠种群密度呈显著负相关,其取食行为与五趾跳鼠种群密度呈显著正相关,其观望行为与其自身种群密度呈显著正相关,其躲避行为与三趾跳鼠和子午沙鼠的种群密度显著正相关 (图7)。
轮牧干扰下,荒漠区3种啮齿动物优势种行为与环境因子的相关分析表明:轮牧区显著影响三趾跳鼠的外部环境因子主要是各层土壤含水量,其主要影响三趾跳鼠除贮食行为外的其他行为;土壤含水量几乎影响五趾跳鼠在轮牧区的所有行为,草本高度和地上生物量主要影响五趾跳鼠的觅食行为;影响子午沙鼠行为的外部环境因子主要是5 ~ 15 cm 土层含水量,其显著影响子午沙鼠的躲避行为 (图7)。
3 讨论
在本研究中,过牧导致五趾跳鼠种群密度增加,同时过牧区五趾跳鼠表现出更高的摄食行为。相反,过牧抑制了子午沙鼠的种群密度,该区域内子午沙鼠的摄食行为表现较低,而在禁牧区占比较高。这与各放牧方式下植被及土壤理化性质等环境因素存在相关性。表明阿拉善荒漠3种啮齿动物优势种的觅食行为对不同放牧方式的响应存在差异。
荒漠啮齿动物往往通过调节对同一资源在空间、时间和类型的合理分配,进而缓减潜在竞争得以实现同域共存,且该调节具有强烈的空间位置特异性,主要受动物自身适应特征和环境特征组合影响 (Mohret al., 2003)。不同物种对密度制约性的生境选择差异有助于物种共存。理论上,放牧家畜通过采食和践踏等方式,改变了荒漠植被密度、盖度、高度、生物量及土壤理化性质等环境因素,从而形成不同的生境类型,并直接或间接影响栖息于荒漠生态系统中啮齿动物的生境选择和摄食策略,即不同物种响应不同放牧方式会表现出生境选择差异和摄食行为时空分配差异(Carusoet al., 2016)。从生境选择来看,在阿拉善荒漠生境中,3 种同域共存的优势啮齿动物摄食行为的差异可能是为避免栖息地和食物选择相似而引起的种内和种间竞争而导致的 (Cregoet al.,2018)。通常,植被条件是制约啮齿动物种群数量消长和分布的重要因素之一,而在啮齿动物群落中,不同物种的活动同时也影响着植物群落的结构,进而对其他物种的分布与多度产生作用,而放牧干扰对上述过程具有明显效应 (Hooper and Vitousek, 1997)。首先,放牧强度的增加影响了生境中食物的数量和质量,生境中的植被种类是啮齿动物做出不同选择的原因之一 (Bejderet al.,2009)。我们之前的研究发现禁牧区有刺叶柄棘豆(Oxytropis aciphylla)、蒙古莸 (Caryopteris mongholica)、黑果枸杞 (Lycium ruthenicum)、柠条锦鸡儿 (Caragana korshinskii) 和沙冬青 (Ammopiptanthus mongolicus) 等植物,子午沙鼠多以这些植物种子为食,故其会选择放牧强度较低的生境摄取食物;相反,五趾跳鼠和三趾跳鼠主要以植物茎和叶为食 (娜日苏等,2009),放牧强度的增加导致沙蓬 (Agriophyllum pungens)、雾冰藜 (Bassia dasyphylla) 和虫实 (Corispermum mongolicum) 等植物密度和生物量显著增加,这些植物均为五趾跳鼠和三趾跳鼠优先选择的食物 (袁帅等,2017)。与禁牧区和过牧区不同,轮牧区的植被数量、盖度、密度以及地上生物量均处于较高稳定状态。本研究表明,在阿拉善荒漠区适当的放牧可以丰富生境内植被类型,有助于多种啮齿动物实现共存,这与以往的研究结论一致 (王艳芬和汪诗平,1999;杨素文,2021)。
不同放牧方式下家畜的啃食、踩踏等也可以导致生境中植被高度和盖度以及土壤质地变化,在荒漠生境中,灌木植被盖度的增加可能不会直接影响哺乳动物群落,但会通过减少草地开阔区间接改变哺乳动物群落状态 (Soto-Shoenderet al.,2018),其中,通过啮齿动物对避难地选择产生影响是重要原因之一。通常植被高度制约动物活动范围和活动速度,啮齿动物在避免生存风险和日常活动间权衡时会通过生理、形态和行为等多方面调节自身活动节律和各行为时间分配来做出响应。同时,这种调节和响应可能与动物的体型大小、是否冬眠等生活史特征有关。也就是说,动物在与自然的博弈中选择最有利的生态小区,从而进化出适合自身的、收益效率最大化的行为策略 (Falcy and Danielson, 2013; Gaynoret al., 2018)。本研究中,子午沙鼠属于四足运动的小型地栖啮齿动物类群,在郁闭度较高的环境中易于找到食物资源且活动不受限制,不容易暴露在天敌的视野中,因此其在禁牧生境中表现出较高的植被依赖性以及种群数量和活动时间优势 (赵天飙等,2001)。三趾跳鼠和五趾跳鼠均属于中等体型以上的啮齿类动物,喜开阔生境的啮齿动物并不适生存于郁闭度较高的环境中,所以它们更适合生存于低矮的草本植物较多且视野开阔的环境以便观察天敌从而降低被捕食的风险,同时最大限度地增加觅食和繁殖的机会 (戴昆等,1998; Logginset al., 2019)。过度放牧生境中草本种类、高度、密度以及地上生物量显著降低,生境开阔,而禁牧区的植被郁闭度高,所以本研究中过牧区比禁牧区更适合跳鼠活动。
以往的研究表明,荒漠区过度放牧时家畜的踩踏以及植被的缺失导致生境中土壤质地较硬且含水量较低,子午沙鼠并不喜选择这类生境 (杨素文,2021)。子午沙鼠对干燥且硬度较低的沙质地选择性更高。这可能与两个原因有关:(1) 子午沙鼠体型较小对灌木种子的采食不便,而沙质地表更多地滞留掉落的新鲜种子,更有利于子午沙鼠对目标食物的捡拾;(2) 子午沙鼠体型较小,沙蟒(Eryx tataricus) 等天敌靠近时沙粒被触动后产生的低频振动信号更容易被子午沙鼠所警惕 (地拉娜·艾力肯等,2010)。而五趾跳鼠和三趾跳鼠是擅于两足跳跃的啮齿动物类群 (梁杰荣和肖运峰,1982),其强有力的后腿需要稍硬的土质来完成跳跃运动,尾巴极长且可控制方向保持平衡,并能竖直敲打地面,增加弹跳力 (赵肯堂,1964)。禁牧土壤基质松软,使较大体重的活动者行动受限制,放牧为三趾跳鼠、五趾跳鼠提供了适宜的活动和摄食环境。此外,五趾跳鼠和三趾跳鼠的栖居洞穴常筑在较坚实的土质中 (甘红军等,2021)。由此可见,在阿拉善荒漠适当的放牧可以通过改变不同生境内动物庇护地和土壤质地,来影响动物栖息选择决策,增加群落物种多样性。
从摄食行为时空分配差异来看,栖息地的复杂性和差异性会影响物种的各种行为分配和摄食策略 (Baker and Brown, 2010),动物调节摄食行为在时间和空间上的投入是其应对种间竞争和种内竞争并平稳的度过食物匮乏期的有效手段 (Mohret al., 2003; Symeset al., 2013)。不同啮齿动物对植物种子的处理能力存在差异,而不同生境中啮齿动物对同类种子的选择和个体对不同行为所投入的时间比例也存在显著差异 (Hodgkisonet al.,2013; Wanget al., 2013)。荒漠生境中不同啮齿动物在面对有限的食物资源时,动物摄食行为可能会表现出差异性,并受环境干扰的影响 (Falcy and Danielson, 2013)。本研究中在相同放牧方式下,3 种啮齿动物呈现出就地取食和贮食两种摄食行为选择差异;而在不同放牧方式下,啮齿动物面对食物觅食、取食和贮食3种行为的分配比例也不同,这同样可能与动物的生活史特征相关联。例如三趾跳鼠和五趾跳鼠多采取就地取食行为,可能反映两者与其冬眠和非冬眠期的能量摄入需求以及形态结构相适应。三趾跳鼠和五趾跳鼠具有冬眠习性,非冬眠期的日常摄食只需满足活动时期的代谢即可,越冬前期需要大量觅食储备脂肪,而跳鼠强有力的后肢,为跳跃到更远的地方采食,获得足够的能量储备提供了形态学基础。对于无冬眠习性的子午沙鼠来说,面对的压力是在满足自身基本能量需求前提下,越冬前尽可能储备大量食物,以满足越冬能量需求 (Shuai and Song, 2011)。因此,子午沙鼠在长期适应进化过程中,更倾向于将食物搬运回巢内贮食,采取边收集边消耗边贮存的摄食行为对策,有利于保障洞内食物足以维持越冬之需 (刘焕金等,1984)。此外,子午沙鼠体型较小导致运动范围也较小,其在生境中的觅食时间明显小于三趾跳鼠和五趾跳鼠 (Schradinet al., 2010;袁帅等,2017)。荒漠区啮齿动物种类较多,本研究只分析了不同放牧方式对啮齿动物物种间摄食策略的影响,同种啮齿动物不同性别、年龄组也可能存在摄食策略差异,但由于红外相机对小型啮齿动物个体识别存在困难,本研究并未涉及性别及年龄组方面的分析。
本研究还发现,在3种放牧方式中种群数量较平均的三趾跳鼠在过牧区觅食频次更高。这可能反映了摄食时间分配受生境资源干扰,主要是食物资源丰富度和食物可利用性的影响 (Frafjord,2016;范言广,2020)。本研究区域,因过度放牧导致生境中食物可用性不高,啮齿动物可选择食物较少,动物个体因此需要转变摄食策略,扩大日常活动区域并付出更多的觅食努力以达到维持自身代谢和能量需求;而禁牧区和轮牧区较过牧区具有更高的植被盖度和多样性,可以为动物提供较高的食物可用性和选择性,因此,禁牧区啮齿动物采取付出较小觅食努力的摄食策略,是其生存适应收益-代价权衡的结果。
综上所述,本研究通过对不同放牧方式下阿拉善荒漠啮齿动物优势种的摄食行为分配研究发现:(1) 荒漠区啮齿动物优势种的摄食行为占比与不同放牧方式下种群密度成正比,子午沙鼠的摄食行为多集中于植被盖度较高的禁牧区,五趾跳鼠的摄食行为在过牧区占比较高;(2) 放牧强度的增加使荒漠区啮齿动物优势种觅食频次增多,花费的觅食努力更大;(3) 相同放牧方式下,子午沙鼠更倾向于“觅食 + 取食 + 贮食”相结合的摄食策略,而三趾跳鼠和五趾跳鼠更倾向于“觅食 + 原地取食”的摄食策略。放牧干扰会通过影响啮齿动物种群密度、栖息地植物和土壤特征间接影响其觅食行为。掌握动物的摄食策略,能更好地理解动物对生存环境的适应策略,从鼠害防治角度来看,根据优势鼠种的摄食策略,适时放牧干预可能会提高对其种群数量的控制效率。
