长爪鼩鼱和细鼩鼱分子系统地理学初步分析
2023-12-22梅秀峰蔡赫韩美凤赵鑫旭张春凤时艳菁陈欢田新民张隽晟刘铸
梅秀峰 蔡赫 韩美凤 赵鑫旭 张春凤 时艳菁陈欢 田新民 张隽晟 刘铸*
(1 牡丹江师范学院生命科学与技术学院,牡丹江 157011)(2 黑龙江农业经济职业学院,牡丹江 157041)
长爪鼩鼱 (Sorex unguiculatus) 和细鼩鼱 (Sorex gracillimus) 隶属劳亚食虫目 (Eulipotyphla) 鼩鼱科(Soricidae) 鼩鼱属 (Sorex)。长爪鼩鼱国内分布于黑龙江、内蒙古东部。国外分布于俄罗斯符拉迪沃斯托克 (海参崴)、萨哈林岛 (库页岛)、日本 (北海道) (Hutterer, 2005;刘铸等,2019)。长爪鼩鼱未见亚种报道 (Hutterer, 2005; Burgin and He, 2018)。细鼩鼱国内分布于黑龙江、吉林、内蒙古东北部。国外分布于从鄂霍次克海东海岸到朝鲜半岛北部、俄罗斯萨哈林岛 (库页岛)、日本 (北海道) (Hutterer, 2005;刘铸等,2016)。细鼩鼱目前分为3 个亚种:S.g.gracillimus分布于萨哈林岛和库纳施尔岛;S.g.granti分布于南千岛群岛的齿舞岛和色丹岛;S.g.minor分布于俄罗斯远东 (马加丹南部到滨海边疆区)、中国东北和朝鲜半岛北部 (Burgin and He, 2018)。分布于日本北海道和北部一些小岛上的细鼩鼱亚种归属还未确定 (Burgin and He,2018)。
长爪鼩鼱和细鼩鼱在分布区域上存在较大重叠 (Hoffmann, 1987; Hutterer, 2005; Burgin and He,2018;刘铸等,2019),然而两个物种的地理系统分化格局可能并不相同。Ohdachi 等 (1997a,1997b) 利用Cytb基因片段 (402 bp) 和化石记录分析了东北亚地区分布的鼩鼱亚科动物的地理史,认为细鼩鼱是分布于东北亚地区的原始祖先成员之一,而长爪鼩鼱在细鼩鼱迁移到日本群岛和萨哈林岛之前,分布于原始东西伯利亚的局域避难所中。Ohdachi 等 (2001) 报道了长爪鼩鼱的分子系统关系和地理分布关系无相关性,而细鼩鼱的分子系统关系和地理分布关系存在一定相关性,尤其是俄罗斯马加丹地区分布的细鼩鼱与其他各分布地域存在明显分子系统分化。可见,已报道的细鼩鼱分子系统地理分化与其亚种分化不一致。我国东北分布细鼩鼱新记录报道中暗示着我国东北分布区的细鼩鼱存在特异化 (刘铸等,2016)。
本研究目的:(1) 明确长爪鼩鼱和细鼩鼱的种群系统地理关系;(2) 进一步明确长爪鼩鼱和细鼩鼱2个物种的亚种分化问题。
1 研究方法
1.1 样本采集
长爪鼩鼱和细鼩鼱样本的采集主要采用陷阱法,辅以铗捕法。在黑龙江省横道河子地区 (采集地海拔约740 m,北纬44°48′44″ ,东经129°02′04″) 采集长爪鼩鼱样本6 号 (HD21、HD26、HD43、HD91、HD94、HD99) 和细鼩鼱样本11 号(HD12、HD19、HD25、HD52、HD106、HD107、HD115、HD135、HD138、HD157、HD183),牡丹峰地区 (采集地海拔约480 m,北纬44°47′48″,东经129°04′52″) 采集长爪鼩鼱样本3 号 (MD60、MD91、MD125) 和细鼩鼱样本1 号 (MD7),双峰地区 (采集地海拔约640 m,北纬44°30′04″,东经128°51′14″) 采集长爪鼩鼱样本2 号 (SF1、SF2),凤凰山地区 (采集地海拔约520 m,北纬44°27′48″,东经128°12′40″) 采集细鼩鼱样本5 号(FH1、FH14、FH102、FH103、FH120)。基于现有数据情况,从GenBank 下载了东北亚其他地区长爪鼩鼱的Cytb基因全序列和细鼩鼱Cytb基因的序列片段 (基本信息见附录A)。
1.2 实验方法
用剪刀剪取样本腿部肌肉组织,肌肉组织用酚-氯仿法进行基因组DNA 提取。线粒体DNA(Mitochondrial DNA, mtDNA) 的Cytb基因全序列PCR 反应体系为50 μL,含200 μmol/L 的dNTP,1.5 mmol/L 的MgCl2,引物L14724/H15915 (Irwinet al., 1991) 各10 pmol,Taq DNA 聚合酶2.5 U(大连宝生物),模板为250 ng。PCR 扩增的反应条件:94 ℃预变性3 min;94 ℃变性45 s,50 ℃退火50 s,72 ℃延伸45 s,共30 个循环;72 ℃延伸7 min,4 ℃保存。PCR 扩增产物经纯化回收,进行测序 (哈尔滨博仕生物技术有限公司)。
1.3 数据处理
鉴于GenBank 现有长爪鼩鼱和细鼩鼱的Cytb基因序列,利用Cytb基因全序列 (1 140 bp) 作为长爪鼩鼱的分子标记;在GenBank 中细鼩鼱仅有Cytb序列片断,为便于比较,本文利用Cytb基因前端序列片段 (630 bp) 作为细鼩鼱的分子标记。使用EditSeq (DNASTAR 软件包) 对实验获得mtDNA Cytb基因序列和GenBank下载序列一并进行编辑,再利用Clustal W (Thompsonet al., 1997) 软件进行序列聚类比对。利用DnaSP 5.10 (Librado and Rozas, 2009) 分析多态位点数量 (Number of polymorphic sites,s)、单倍型数量 (Number of haplotypes,k)、核苷酸多态性 (Nucleotide diversity, π)、核苷酸差异平均数 (Average number of nucleotide differences,i) 及单倍型多态性 (Haplotype diversity,h)。利用软件RAxML 采用密码子分区段分析方式构建最大似然法 (Maximum likelihood, ML) 系统发生树(Stamatakis, 2014)。利用Network 10.2 (Bandeltet al., 1999) 构建中介网络图 (median-joining network) 分析单倍型间的进化关系。利用DnaSP 5.10软件计算Tajima’s D 和 Fu’s Fs 参数,检测中性选择情况,进行显著性检测,并结合种群的增减模型 (A population growth-decline model) 分析错配分布来检测种群的数量扩张情况 (Rozaset al., 2003)。
2 结果
2.1 Cyt b基因的遗传多样性
长爪鼩鼱的Cytb基因全序列1 140 bp,包含77 个变异位点,其中41 个简约信息位点 (Parsimony informative sites),36 个单变异位点 (Singleton variable sites),变异位点数占总位点数的6.75%,未见插入和缺失。细鼩鼱的Cytb基因序列630 bp,包含35 个变异位点,其中27 个简约信息位点,8 个单变异位点,变异位点数占总位点数的5.56%,未见插入和缺失。
长爪鼩鼱和细鼩鼱的遗传多样性参数见表1。长爪鼩鼱在43 个样本中发现36 个单倍型,核苷酸多态性0.015 5。细鼩鼱在39个样本中发现22个单倍型,核苷酸多态性0.016 3。

表1 长爪鼩鼱和细鼩鼱的遗传多样性参数Table 1 The genetic diversity indices of Sorex unguiculatus and Sorex gracillimus
2.2 长爪鼩鼱和细鼩鼱的系统发育分析
系统发生树显示长爪鼩鼱存在2 个进化支系,说明不同样本之间存在一定的遗传差异 (图1a),但是这2个支系的后验支持率较低 (支持率27),因此长爪鼩鼱种内不存在稳定的遗传分化。其次,来自同一地区的样本,分别聚类于不同进化分支上。我国横道河子地区的6个样本分别聚类于分化关系较远的2个进化分支上,HD21和HD26与地理分布关系较远的3 个日本北海道样本 (Hokkaido 8、Hokkaido 21 和Hokkaido 23) 和1 个俄罗斯萨哈林岛样本 (Sakhalin 2) 聚类于同一小分支上,而HD43、HD91、HD94 和HD99 与我国其他分布区样本以及1 个日本北海道样本 (Hokkaido 17) 聚类于同一小分支上 (图1a)。说明长爪鼩鼱的分子系统关系与地理分布无相关性。
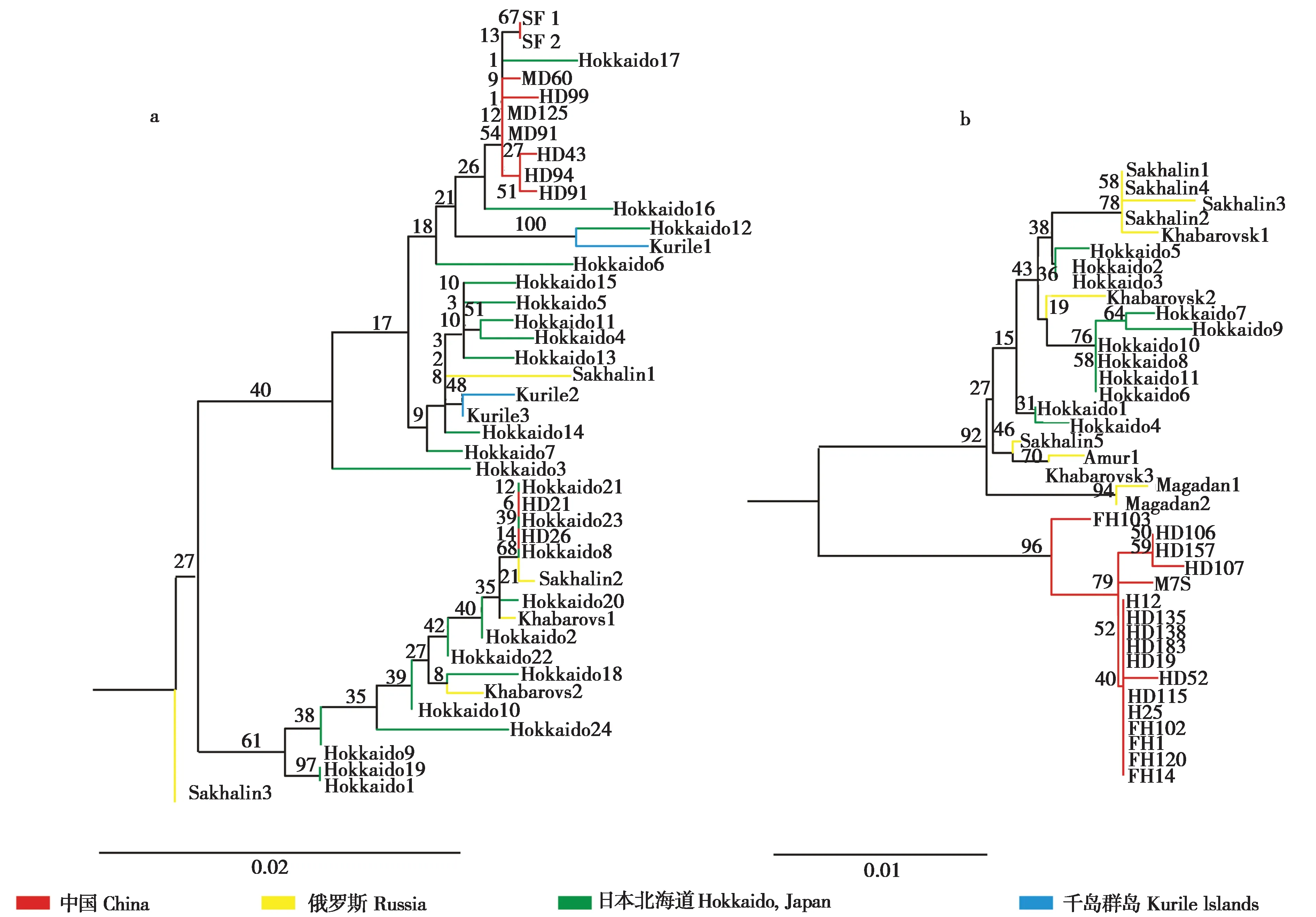
图1 基于Cyt b 基因构建的长爪鼩鼱 (a) 和细鼩鼱 (b) 的ML系统进化树Fig.1 The ML Phylogenetic tree of S. unguiculatus (a) and S. gracillimus (b) based on the Cyt b gene
系统发生树显示细鼩鼱分子系统关系与地理分布表现出一定的相关性。细鼩鼱样本分为2 个分支,且表现出了较为显著的分化,来自于俄罗斯和日本北海道的细鼩鼱样本构成一个进化支,我国东北样本构成另一个独立的进化支 (图1b)。
2.3 长爪鼩鼱和细鼩鼱的中介网络分析
中介网络图显示我国长爪鼩鼱单倍型分布在中介网络图的两端,右端2 个样本与日本北海道3 个样本具有共同的单倍型和网络分化地位,左端7 个单倍型聚成一簇,同样表现出与地理距离较远的日本北海道单倍型具有较近的网络关系 (图2a)。中介网络图明显表现出长爪鼩鼱的分子系统关系与地理分布无相关性。
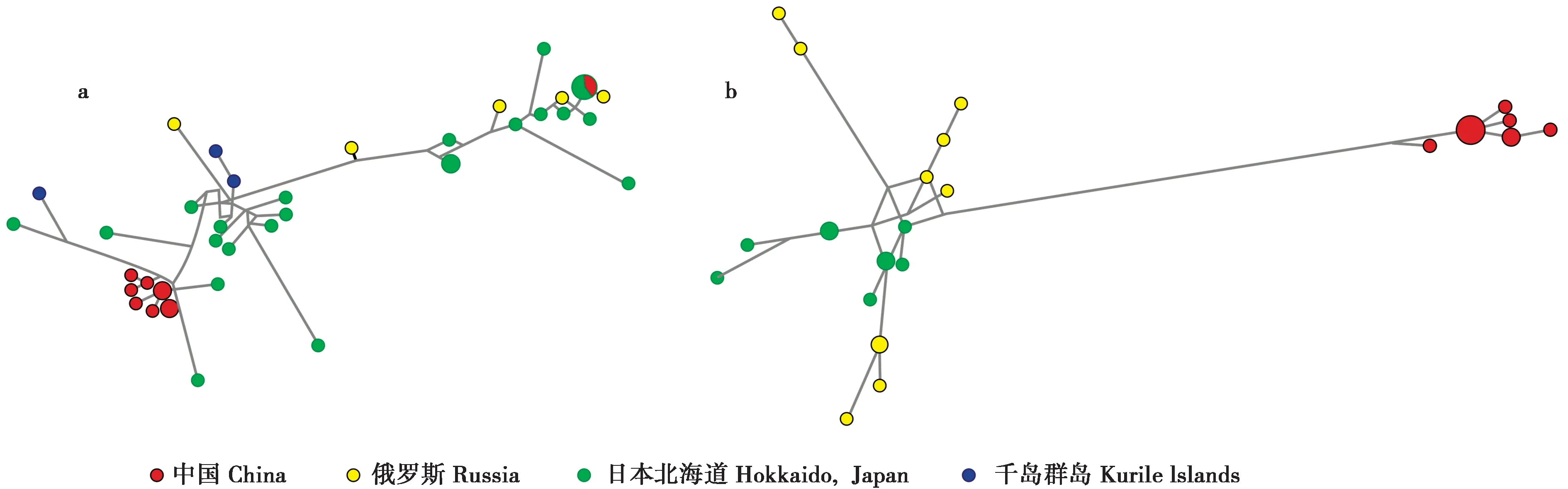
图2 长爪鼩鼱 (a) 和细鼩鼱 (b) 的单倍型中介网络图. 长爪鼩鼱最短树的突变数为141,细鼩鼱最短树的突变数为44,不同地理分布的样本单倍型用不同的颜色进行表示Fig. 2 Median-joining network of haplotypes of S. unguiculatus (a) and S. gracillimus (b). The number of mutations of the S. unguiculatus shortest tree = 141. The number of mutations of the S. gracillimus shortest tree = 44. Geographical origin of haplotypes of samples is indicated by color
细鼩鼱的分子系统关系与地理分布表现出一定的相关性。中介网络图分成两大部分,我国东北样本都分布在中介网络图的右端,其中出现由11 个样本构成的主要单倍型,位于中介网络图右端中央位置 (图2b)。俄罗斯和日本北海道样本分布于中介网络图的左端,表现出与我国东北样本存在明显的分化,日本北海道单倍型之间表现出较近的网络关系,左上方2个来自于俄罗斯马加丹地区的单倍型,表现出与其他单倍型具有较远的网络关系 (图2b)。
2.4 长爪鼩鼱和细鼩鼱的种群历史动态分析
长爪鼩鼱的Tajima’s D 结果 (D = -0.1164,P> 0.1) 尽管不显著,但为负值,Fu’s Fs 结果(Fs = -12.7320,P< 0.001) 为显著性负值 (表1)。中性检测结果显示长爪鼩鼱近期可能发生过种群扩张。细鼩鼱的Tajima’s D 结果 (D = 0.7558,P>0.1) 为非显著性正值,Fu’s Fs结果 (Fs = -3.3370,P> 0.01) 为非显著性负值 (表1),中性检测结果显示细鼩鼱未经历过数量急剧扩张。采用错配分布方法对种群历史进行检验,长爪鼩鼱的错配曲线是多峰曲线 (图3a),说明长爪鼩鼱近期尽管经历了种群扩张,但并非为单次急剧扩张。细鼩鼱的错配曲线是双峰曲线 (图3b),说明细鼩鼱经历了较为复杂的历史,但并未经历过单次数量急剧扩张。

图3 突然扩张模型下长爪鼩鼱 (a) 和细鼩鼱 (b) 的错配分布图Fig. 3 The observed pairwise difference and the expected mismatch distributions for S. unguiculatus (a) and S. gracillimus (b) under the sudden expansion model
3 讨论
3.1 长爪鼩鼱和细鼩鼱的遗传分化
本研究结果显示,长爪鼩鼱的系统发生树未表现出明显的地理遗传分化,来自俄罗斯萨哈林岛和日本北海道的样本并未表现出岛屿种群与大陆种群存在遗传分化,甚至是来自于千岛群岛,这些小岛的样本也未表现出明显的遗传分化。推测长爪鼩鼱种群离开冰期避难所后,占据目前的分布地域时间较短,还未形成进一步地理上的遗传分化。刘铸等 (2020a) 对中鼩鼱 (S.caecutiens)的遗传分化进行了分析,欧亚大陆分布的中鼩鼱的遗传分化结果与本研究得出的遗传分化结果类似,即未形成进一步的遗传分化,但分布于济州岛和北海道的中鼩鼱显现出一定的遗传分化,推测长爪鼩鼱种群离开冰期避难所后,占据目前的分布地域的时间要晚于中鼩鼱。
本研究结果显示,细鼩鼱的系统发生树表现出明显的地理遗传分化,特别是我国分布的细鼩鼱单独聚为一支。在俄罗斯和日本的进化分支上,分布于地理距离较远的马加丹的样本,也显现出了明显的遗传分化,可见细鼩鼱在欧亚大陆上存在的遗传分化,不同于长爪鼩鼱和中鼩鼱 (刘铸等,2020a),推测细鼩鼱种群离开冰期避难所后,占据目前的分布地域时间较长,已经形成进一步的遗传分化。本研究结果与Ohdachi等 (2001) 认为细鼩鼱分布于现在的分布区要久远于长爪鼩鼱的结论一致。
3.2 长爪鼩鼱和细鼩鼱的分子系统地理学分析
Hutchison 和Templeton (1999) 诠释了种群结构的踏石模型 (stepping-stone model),认为在一个物种 (种群) 的分布区内,遗传距离和地理距离的关系能够说明基因流和遗传漂变在物种 (种群) 的分布区扩张或扩散中的影响。也就是说,如果遗传距离和地理距离表现出正相关,说明这个物种 (种群) 占据目前的分布区已经很长一段时间了,这段时间足够使基因流和遗传漂变达到平衡。长爪鼩鼱的遗传分化与地理距离无相关性,推测其占据目前的分布区时间较短,基因流和遗传漂变未达到平衡。细鼩鼱的遗传分化与地理距离表现出相关性,推测其占据目前的分布区域时间较长,已经形成进一步的遗传分化。
系统发生树和中介网络图显示长爪鼩鼱的分子系统地理关系基本一致,所有样本聚集在一个大支上,进化支内部缺乏明显的进一步的遗传分化,与以往的报道结果类似 (Ohdachiet al., 1997a,1997b, 2001),表现出长爪鼩鼱遗传分化与地理分布关联性较弱。在第四纪冰期利用同一个冰期避难所的哺乳动物,例如灰狼 (Canis lupus)、貂熊(Gulo gulo)、紫貂(Martes zibellina) 和北松鼠 (Sciurus vulgaris) 具有类似的研究结果 (Vilaet al.,1999; Walkeret al., 2001; Liet al., 2013; Liuet al.,2014),推测长爪鼩鼱源自于相同的冰期避难所。Ohdachi 等 (1997a, 1997b) 认为长爪鼩鼱在细鼩鼱迁移到日本群岛和萨哈林岛之前,存在于原始东西伯利亚的局域避难所中,本研究的分析结果与其一致。系统发生树和中介网络图显示细鼩鼱的分子系统地理关系基本一致,我国东北分布区样本与俄罗斯和日本北海道样本存在明显的遗传分化,推测细鼩鼱在第四纪冰期利用了2个冰期避难所。鼩鼱属已有研究显示诸多物种的冰期避难所和冰期后扩散与其分布的北方针叶林 (扁颅鼩鼱S.roboratus、中鼩鼱、远东鼩鼱S.isodon、姬鼩鼱S.minutissimus) 或温带针阔混交林 (普通鼩鼱S.araneus) 度过冰期的历史事件有着密切联系(Hopeet al., 2010; Bannikovaet al., 2010, 2018; Raspopovaet al., 2018)。目前细鼩鼱主要分布于温带针阔混交林 (刘铸等,2019),推测细鼩鼱冰期避难所的利用与其生境中的主要树种避难所的利用有着密切关系。我国东北地区的樟子松 (Pinus sylvestris)、赤松 (P.densiflora)、长白松 (P.funebris)、兴凯湖松 (P.takahasii)、臭冷杉 (Abies nephrolepis) 和杉松 (A.holophylla) 的系统地理学分析都认为长白山避难所是东北亚地区第四纪冰期的一个重要避难所 (Jianget al., 2011; Renet al.,2012),远东鼩鼱的分子系统地理学分析也显示其利用了长白山避难所 (刘铸等,2020b)。推测我国东北分布区的细鼩鼱来源于冰期的长白山避难所。对蒙古栎 (Quercus mongolica) 的系统地理学分析发现了第四纪冰期俄罗斯远东避难所的存在(Zenget al., 2015),推测俄罗斯和日本北海道的细鼩鼱来源于冰期俄罗斯远东避难所。
长爪鼩鼱目前未见亚种报道 (Hutterer, 2005;Burgin and He, 2018),本研究系统发生树和中介网络图结果支持长爪鼩鼱无亚种分化的观点。细鼩鼱目前分为3 个亚种:S.g.gracillimus分布于萨哈林岛和库纳施尔岛;S.g.granti分布于南千岛群岛的齿舞岛和色丹岛;S.g.minor分布于俄罗斯远东 (马加丹南部到滨海边疆区)、中国东北和朝鲜半岛北部 (Burgin and He, 2018)。目前分布于日本北海道和日本北部的一些小岛上的细鼩鼱还未见亚种归属的报道 (Burgin and He, 2018)。由于本研究未包含分布于南千岛群岛的齿舞岛和色丹岛的细鼩鼱样本,其亚种分类地位不能确定。本研究的细鼩鼱系统发生树和中介网络图结果支持将本研究包含的地理区域分成两个地理亚种,一个亚种分布于中国东北,另一个亚种分布于俄罗斯远东 (马加丹南部到滨海边疆区,包括萨哈林岛)和日本北海道。
