麻阳河国家级自然保护区黑叶猴野外投食群动态观察
2023-12-22张鹏颜修刚石运杨德王彬李伟杨远露杨朝辉吴光阳蔡鸷李思尧邹浩张璠
张鹏 颜修刚 石运 杨德 王彬 李伟 杨远露 杨朝辉 吴光阳 蔡鸷 李思尧 邹浩 张璠
(贵州麻阳河国家级自然保护区管理局,沿河 565300)
黑叶猴 (Trachypithecus francoisi) 属灵长目猴科疣猴亚科乌叶猴属 (Trachypithecus)。全球范围内分布很窄,仅分布于中国的贵州、重庆、广西以及越南、老挝的部分地区,分布范围窄与其生活的石山环境和物种形成机制有密切关系 (Nadler and Brockman, 2014),目前全球黑叶猴数量接近2 500 ~ 3 000 只 (黄乘明等,2018)。《国家重点保护野生动物名录》(2021) 中黑叶猴为国家一级重点保护野生动物,《世界自然保护联盟濒危物种红色名录》(IUCN) (2020-2) 为濒危 (EN),列入濒危野生动植物种国际贸易公约 (CITES) 附录 (2019)Ⅱ物种。随着栖息地质量改善,人为猎杀减少,全球黑叶猴种群正在恢复。贵州麻阳河国家级自然保护区 (以下简称:麻阳河保护区) 是我国野生黑叶猴分布最密集、种群数量最多的地区,保护区及周边区域共有黑叶猴72 群,554 只 (牛克峰等,2016)。
黑叶猴社会结构通常为一雄多雌群体,由1只成年雄猴,几只成年雌猴及其未成年后代组成,猴群一般3 ~ 10只,较大的群体达20只左右,有一定的活动规律和较为固定的住所,每群均具有相对固定的家域。目前,针对黑叶猴种群行为生态学在笼养黑叶猴的饲养与繁殖 (姚琳和毕延台,2017;郭爱霞等,2020)、种群现状 (高喜凤,2014)、寄生虫防治与发病规律 (胡巍堃等2017;农汝等,2020)、日活动时间分配 (韩家亮等,2013;田明亮等,2019)、个体行为 (黎大勇等,2013;张嘉欣等,2019) 等方面进行了比较全面的研究。自然生境条件下,主要针对黑叶猴种群数量与分布 (Huet al., 2011;邓怀庆和周江,2018)、食性及行为 (王静等,2013;李生强等,2016)、活动节律与时间分配 (周岐海等,2007;洪伟和刘宁,2014)、保护现状与研究进展 (胡刚,2011;周岐海和黄乘明,2021) 等方面。关于野生黑叶猴繁殖行为及社群结构动态研究较少,在自然生境条件下,仅有黑叶猴繁殖周期的初步研究 (吴安康等,2006) 和黑叶猴生态行为研究 (胡刚,2018) 报道。
本研究选择麻阳河保护区生境干扰强度大、黑叶猴种群分布密集、活动空间受限制、食物极度匮乏的大河坝区域。该区域分布5 群黑叶猴 (汆洞群、雷家群、香菇坝群、猴王洞群及温泉投食群),温泉投食群与香菇坝群和猴王洞群比邻,生活家域相互重叠。在该区域开展人工投食半驯化监测,以缓解猴群季节性食物缺乏压力,减少区域人为及牲畜活动产生的威胁,消除对种群发展的不利因素。人工半驯化猴群不仅方便生态学观察研究,也有利于动物保护,及时掌握动物资源消长情况,有利于开展科学研究和合理利用动物资源 (刘振河等,1986)。此外,人工投食可以近距离观察猴群个体,能有效进行个体辨认,极大地方便了野生群体的行为生态学研究,从而能更加真实反映野生种群动态特征。黑叶猴个体识别一直是深入研究的瓶颈,本文旨在人工投食个体识别的基础上,对一个半驯化猴群13 年繁殖情况和种群结构进行观察研究,为保护该物种提供参考。
1 研究方法
1.1 研究地点概况
麻阳河保护区位于北纬28°37′30″ ~ 28°54′20″,东经108°03′58″ ~ 108°19′45″,总面积31 113 hm2。区域地貌属黔北中山峡谷,以石灰岩山地为主,单面山和箱状谷发育;海拔800 ~ 1 000 m 以上地势平坦开阔,800 m 以下则多为深切沟谷,向下侵蚀作用强烈,村寨多分布在平缓地带;气候类型属中亚热带温暖湿润季风气候,年均降水量1 158.7 mm,年均温17.6 ℃,极端最高温41.5 ℃,极端最低温-2.3 ℃,无霜期306 d 左右。全区森林覆盖率77.9%,森林植被为中亚热带湿润常绿阔叶林,森林群落类型多样,原生植被多遭破坏,现存植被大多处于不同演替阶段。
1.2 观察对象
本研究以栖息在麻阳河保护区大河坝区域的一个黑叶猴群 (温泉投食群) 为观察对象。猴群活动范围以温泉为中心 (北纬28°39′29.74″,东经108°15′3.45″),北至老鹰岩下岩,南至香菇坝公路,西至猴王洞,东至大河坝桥,日常活动面积约为66.7 hm2,共有5 个固定夜宿点 (图1)。猴群家域南北两岸为陡峭悬崖,垂直高度160 m 左右,峡谷内次生植被保存较完整,竹类资源丰富,深切谷底溪流不断,形成了适于黑叶猴生存的独特栖息环境。从1999 年开始,投食人员对该群4 只(雄性1 只,雌性3 只) 个体进行长期追踪,并人工定期定点适度投食,探索野外生境下黑叶猴的招引驯化方法。投食少量科学,确保不妨碍野生种群的自然觅食能力。通过多年的人工管理和投食驯化,2009 年成功实现了与该群体近距离接触,在不影响其自然生活习性的情况下,投食人员对猴群个体及群体行为活动进行观察记录。

图1 麻阳河保护区黑叶猴投食群生活家域地理位置示意图Fig.1 Sketch of geographical location of provisioned group of Trachypithecus francoisi in Mayanghe National Nature Reserve
1.3 观察方法
采取直接观察法对投食群及个体行为进行直接观察和记录。投食猴群经过长期野外投食驯化,对投食的食物及方式已适应,投食人员每日在其活动范围内,通过吹哨的方式召集,近距离观察猴群动态,记录猴群数量、个体特征、行为活动、繁殖情况、哺乳行为、毛色变化、进食状况、死亡及失踪 (家域内观察到尸体,即为死亡;一周内未发现个体,即为失踪) 等,并拍摄猴群个体照片。同时,记录天气情况、投食位置、投食人员、参观人员、食物类型及食物量等。投食观察每日1 次 (06: 00—08: 00),06: 00 到达猴群夜宿点附近区域,近距离提供食物或将食物放在易观察点,吹哨召唤黑叶猴至投食点。投食的食物主要以本地红薯为主,投食量1.5 ~ 2.5 kg,除了受天气影响、季节变化及其他不可抗拒因素影响外,全年投食监测观察。2009—2021 年,共观察1 846 d,平均每年142 d 左右。在野外投食观察过程中,根据体型大小、毛色变化及个体行为等,将黑叶猴划分为6 个年龄组:婴猴组 (0 ~1 月龄)、幼猴组 (2 ~ 12 月龄)、亚成年组 (1 ~3 岁) 、成年组 (4 ~ 5 岁)、中年组 (6 ~ 15 岁)、老年组 (大于16 岁) (高喜凤,2014;张嘉欣等,2019;郭爱霞等,2020)。
2 结果
2.1 投食群年度数量动态变化
投食群年度数量动态变化,以每年12 月底记录的现存个体数为依据,不包括年度死亡及失踪个体。2009—2021 年,年度种群平均约为9 只,年度种群最小5 只个体,最大13 只个体。在这期间观察到4次外来雄猴挑战投食群主雄事件,双方经过多次反复打斗、驱赶及杀戮,导致猴群结构发生较大变化,群体个体数量减少,但每个主雄统领阶段,猴群较稳定,正常繁殖,群体数量呈现正增长。其中,2009—2014 年,种群数量呈现逐步上升,平均每年净增1 只个体;2015—2018 年,种群数量增长速度较快,平均每年净增约2 只个体;2019—2021 年,猴群发生2 次主雄更替,因受其影响,猴群出生个体与损失个体相等,年度群体数量维持在9只左右 (表1)。
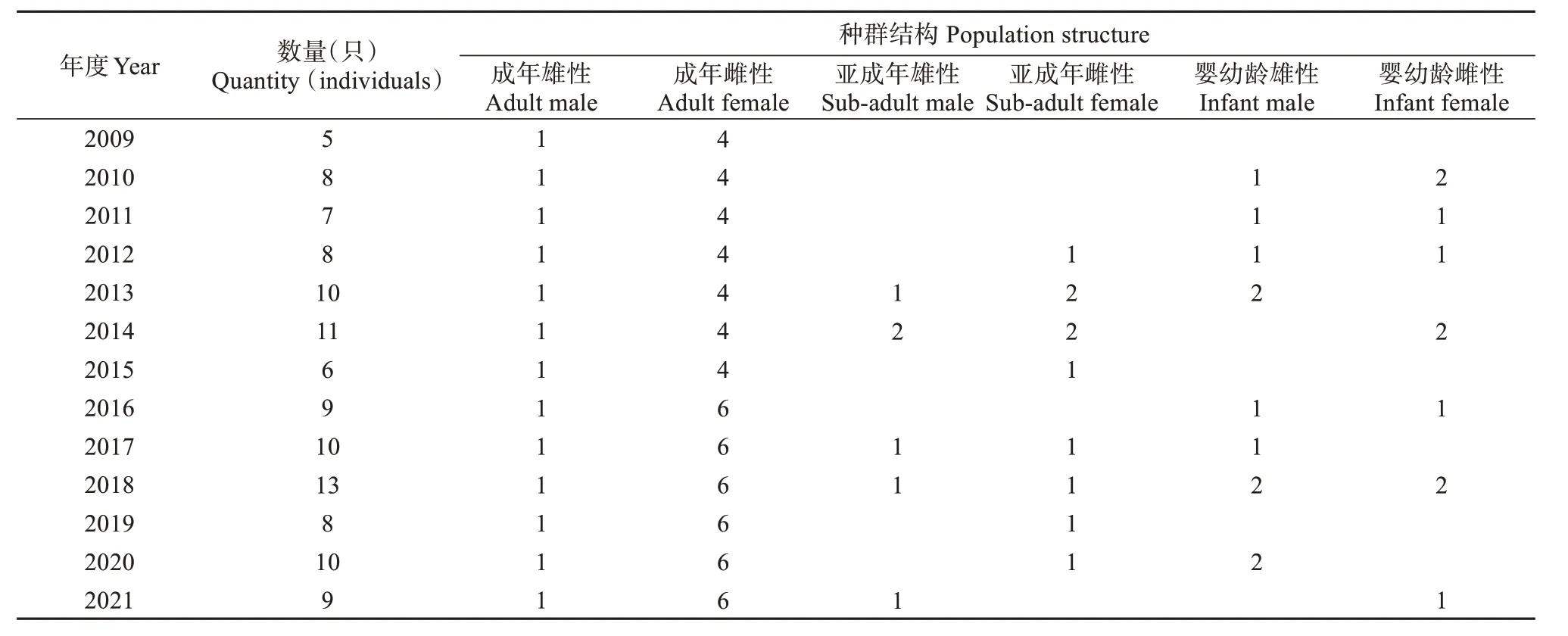
表1 麻阳河保护区黑叶猴投食群年度数量变化Table 1 Annual change in quantity of provisioned group of Trachypithecus francoisi in Mayanghe National Nature Reserve
2.2 投食群年度出生情况
2009—2021 年观察记录,猴群出生婴猴累计26只 (雄性14只,雌性12只) 个体,出生婴猴性比约1.2∶1,猴群年度平均出生2只个体,年平均出生率为22.8% (年平均出生个体数 / 年平均个体数 ×100%)。除2015 年和2019 年主雄更替导致种群未出生婴猴外,其余各年度均有婴猴出生,年度出生婴猴最多达4 只,最少1 只,出生3 只婴猴年度占45.5%。出生婴猴存在奇偶年度差异,偶数年出生婴猴18只 (雄性8只,雌性10只),奇数年出生婴猴8只 (雄性6只,雌性2只),偶数年出生婴猴数量占统计年度出生婴猴总数的70%,奇数年仅占30%,且偶数年出生的雄性个体少于雌性个体,奇数年出生的雄性个体则多于雌性个体,投食群雌猴多在奇数年交配受孕,偶数年产仔 (表2)。
2.3 投食群年度死亡和失踪情况
投食群死亡和失踪累计13 只 (雄性8 只,雌性5 只) 个体。其中,婴猴2 只 (雌性)、幼猴9 只 (雄性7 只,雌性2 只) 和成年猴2 只 (雄性1 只,雌性1只),无亚成体猴死亡和失踪记录,猴群年均死亡和失踪1 只个体,年均死亡和失踪率为11.4% (年平均死亡和失踪个体 / 年平均个体数 × 100%)。投食群幼猴死亡和失踪高于婴猴和成年猴,且婴幼猴存活时间均短于1年 (12 ~ 226 d)。投食群雄性与雌性死亡和失踪比为1.6∶1,雄性死亡和失踪率高于雌性;雄性与雌性幼猴死亡和失踪比为3.5∶1,雄性死亡和失踪率同样高于雌性。观察发现,投食群婴幼猴死亡和失踪主要集中在1 ~ 8 月龄,且7 ~8月龄死亡和失踪达到最大值4只个体 (表2)。
2.4 投食群婴猴繁殖月份及干湿季影响
投食群婴猴出生高峰期在2—4 月、7 月和9—10 月。2—4 月合计出生11 只婴猴,占全年出生婴猴数量42.3%;7 月和9—10 月共出生10 只,占全年出生婴猴数量38.5%;其次,1 月出生2 只,5 月、11 月和12 月均出生1 只,4 个月仅占全年出生婴猴数量的19.2%。出生相对高峰期在2月、4月和10 月,其次是3 月、7 月和9 月,上、下半年各有3个月的出生高峰 (图2)。
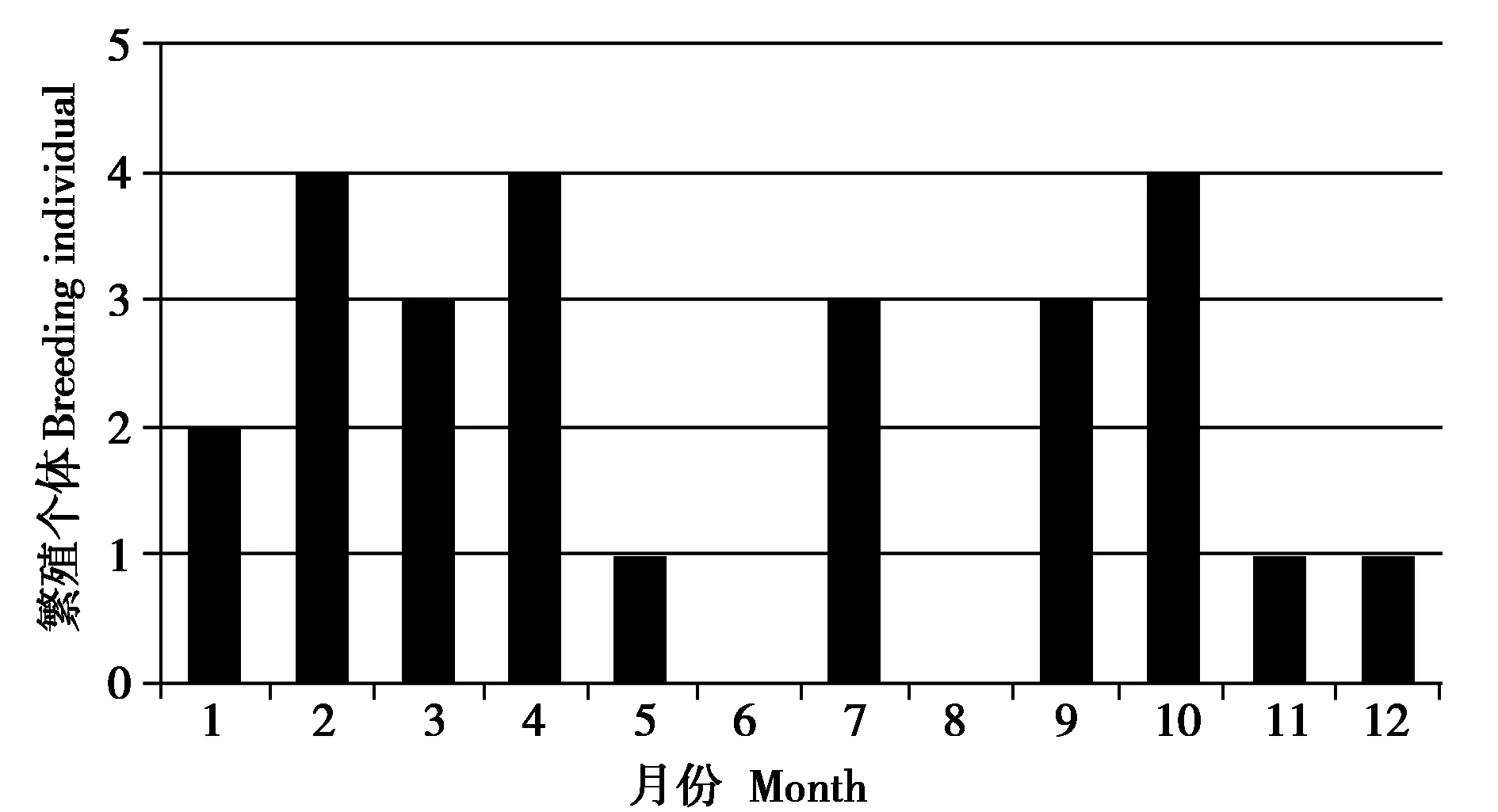
图2 麻阳河保护区黑叶猴投食群繁殖月份统计 (2009—2021)Fig. 2 Breeding month statistics of provisioned group of Trachypithecus francoisi in Mayanghe National Nature Reserve (2009-2021)
保护区秋冬干春夏湿,干湿季节较明显,秋冬降水量占全年的29.5%,春夏占70.5%,干季(秋冬季) 9 月至次年2 月,出生婴猴15 只,湿季(春夏季) 3—8 月,出生婴猴11 只,干季与湿季出生婴猴数量分别占全年出生婴猴数量的57.7%和42.3%,在水源和食物资源充足条件下,黑叶猴生殖行为相对适应较干旱的环境。
2.5 现投食群性比与年龄结构
截至2021年12月31日,野外投食群仅有9只个体,雄雌比为2∶7。其中,老年雌猴3只 (均大于17 岁),中年雌猴2 只 (1 岁和9 岁),中年雄猴1只 (2018年的外来挑战者,大于8岁),成年雌猴1 只 (5 岁),亚成年雄猴1 只 (1 岁),幼雌猴1 只(8个月),种群中无婴猴及2 ~ 4岁年龄段亚成年猴。
2.6 投食群生殖间隔周期
2009年5月,投食群主雄更替。2009年5 月—2015 年7 月,该群无外来雄猴挑战干扰,猴群相对稳定,这期间共计出生12 只婴猴 (雄性7 只,雌性5只),每只雌猴均生产3只,共记录4 只雌猴12 次生殖情况 (雌猴的生产记录是以观察时间内,发现新生小婴猴时间为准)。其中,前四次为新主雄迁入后杀死婴猴,使4 只终止哺乳成年雌猴(D1 ~ D4) 提前怀孕并产仔,平均生殖间隔周期为(488 ± 2) d,约16.3个月 (D1 12.3个月,D2 16.7个月,D3 16.9 个月,D4 19.2 个月);后八次为投食群正常情况下雌猴的生殖记录,生殖间隔周期平均为 (559 ± 3) d,约18.7个月。
3 讨论
观察期间,投食群发生了4次主雄更替,时间在5 月和8 月。主雄地位都是通过反复打斗获得,打斗期间没有婴猴出生和成年雌猴怀孕。雄性更替发生在雌性受孕之前,可以增加新主雄的繁殖成功机会 (Xianget al., 2014)。主雄更替打斗现象在疣猴亚科的白头叶猴 (Presbytis leucocephalus)(于凤琴等,2022)、花斑疣猴 (Colobus vellerosus)(Sicotte and Macintosh, 2004) 和神农架川金丝猴(Rhinopithecus roxellana) (杨万吉等,2022) 等物种中均有报道。而秦岭川金丝猴主雄替换模式中,没有观察到新旧主雄的打斗,推测是单元内雌性主导了主雄更替,入侵雄性采取不断靠近的策略吸引雌性迁移到自己身边形成新单元 (Renet al.,2007)。非人灵长类主雄替换方式多种多样,其中通过引诱雌性的方式建立家庭的可能性不大,最终只能是短暂的接触和聚集,比如埃及狒狒 (Papio hamadryas) 和戴帽叶猴 (Trachuypithecus pileatus) 繁殖群雄性会花费大量精力阻止其他雄性个体接近群内的雌性 (吴秦伟等,2011)。与之相比,直接挑战繁殖群主雄更易成功,但也伴随诸多危险。本研究2015 年8 月观察到投食群主雄与挑战者打斗,造成双方伤残。入侵者直接挑战主雄是猴群重要的新老更替形式,动物界优胜劣汰的自然选择,有利于种群保存更优秀的遗传基因 (胡刚,2011)。
本研究发现,投食群黑叶猴全年均可交配怀孕产仔,且婴猴出生月份存在双高峰期现象,上半年和下半年均有3 个月的出生高峰,仅6 月和8 月未出生婴猴。但在其他猴群的观察和保护区野外监控探头中,均发现黑叶猴6 月和8 月携带婴猴记录 (2014 年6 月野外观察贵阳坝群,2014 年6 月和2019 年8 月监控探头记录猴王洞群),红外线相机同样监测到了野外不同季节黑叶猴携带幼仔情况。这与李明晶等 (1989)、朱本仁和谢华彪(1999) 的黑叶猴不存在季节性繁殖结果一致。大多数亚洲叶猴类没有明显的繁殖季节,只有生育高峰期 (Poirier, 1970),如川金丝猴产仔期集中在3月和4 月 (陈服官等,1983)。随着对各种疣猴研究的不断深入,具有生育高峰被认为是亚洲疣猴类的种属特征 (任宝平,2002)。栖息地气候和食物的变化,可能是影响黑叶猴交配和产仔的重要因素,在降水量最多的5—6 月,猴群仅出生1 只婴猴;在食物相对匮乏的夏季和冬季,出生婴猴数量仅占全年出生婴猴数量的38.5%。这一情况符合野生动物产仔是在其生活条件较好时期的规律,有利哺乳和婴猴成长发育 (陈服官等,1983)。此外,投食群死亡和失踪率比例高,可能与喀斯特石山生态系统遭受破坏有很大关系。
本研究观察记录到野外投食群黑叶猴个体合计31 只 (雄性15 只,雌性16 只),实际存活个体18 只 (雄性7 只,雌性11 只),截至2021 年12 月底本群仅有9 只个体 (雄性2 只,雌性7 只),中老年个体所占比例大,猴群年龄结构不合理。
黑叶猴作为西南喀斯特地区的旗舰物种,其生存状态和保护成效直接反映了本地区的生物多样性现状和保护水平 (黄乘明等,2018)。研究区人口急剧增长,交通、通讯、住宅、电力、水利等基础设施建设,导致黑叶猴行为活动及生存空间风险增加。频繁的人类活动造成非人灵长类栖息地破碎化,导致栖息质量下降,进而影响其家域范围、食性、活动时间分配、繁殖率、死亡率以及存活情况 (Menon and Poirier, 1996)。区域生态系统的修复与维护、物种保护与社区发展、生态旅游与自然教育、人猴冲突与猴畜食物竞争以及病毒交叉感染等需要全社会广泛关注与参与。管理者在科学制定保护行动计划基础上,需积极开展各个领域多方合作研究,以应对栖息地严重干扰和破坏下黑叶猴行为适应策略变化,确保黑叶猴种群及栖息地得到有效保护。
致谢:黑叶猴投食驯化工作,得到了贵州麻阳河国家级自然保护区管理局的大力支持;在数据收集、整理和核实过程中,梁桥管理站专职护林员肖志坚、毛晓林、陈秋明等鼎力相助;贵州民族大学Nat Panda 博士对英文摘要进行了修改,特此致谢!
