深圳坝光核坝路植被组成及其物种多样性研究
2023-12-17蒋呈曦余华金杨志明李鑫泰黄玉源李法民李建仪秦介堂
蒋呈曦 余华金 杨志明 李鑫泰 黄玉源 李法民 李建仪 秦介堂
摘要:采用样方调查法对深圳坝光核坝路自然林、半自然林和人工林进行了调查,分析了其物种组成、多样性水平以及不同林类差异的原因,为坝光区域植被与生态保护提供数据支持,也为市政道路建设与当地生态系统保护相结合提供科学依据。结果显示:(1)共记录有维管植物123种,隶属于57科98属;(2)自然林各物种多样性指数均高于人工林,科、属、种的综合多样性指数也高于人工林,人工林群落科、属、种的丰富度则略高于自然林;(3)自然林和半自然林的均匀度指数均高于人工林群落;(4)自然林与人工林的群落间相似性系数最低,而半自然林与人工林之间的物种相似性及科属种的综合相似性相对较高。
关键词:深圳坝光;物种组成;物种多样性
中图分类号:718.54+2 文献标志码:A
Study of Vegetation Composition and Species Diversity in Heba Road, Baguang District, Shenzhen
JIANG Chengxi1, YU Huajin2,3, YANG Zhiming2, LI Xintai1, HUANG Yuyuan2,3*, LI Famin1, LI Jianyi2, QIN Jietang2
(1Bureau of Public Works of Dapeng New District, Shenzhen, Guangdong 518120, China; 2Shenzhan Maosen
Ecological Sciences and Technology Co., Ltd., Shenzhen ,Guangdong 518120, China; 3College of Agriculture and
Biology, Zhongkai University of Agriculture and Technology, Guangzhou,Guangdong 510225, China)
Abstract: A sample survey was conducted using the quadrat method to investigate the species composition, diversity levels, and reasons for differences among natural forests, semi-natural forests, and artificial forests along the Heba Road in Baguang District, Shenzhen. The study would serve as data support for vegetation and ecological conservation in the Baguang District, as well as scientific evidence for integrating municipal road construction with local ecosystem protection. The results revealed that: (1) A total of 123 vascular plant species were observed, which belonged to 57 families and 98 genera. (2) Compared with artificial forests, natural forests exhibited higher diversity indices for all species, as well as higher composite diversity indices of families, genera, and species. However, the species richness of families, genera and species in artificial forests was slightly higher than in natural forests. (3) Both natural forests and semi-natural forests had higher evenness indices than artificial forests. (4) The similarity coefficient among all communities was the lowest between natural and artificial forests , while the species similarity and the comprehensive similarity of families, genera, and species were relatively higher between semi-natural and artificial forests.
Key words: Baguang District of Shenzhen; vegetation composition; species diversity
隨着全球城市化进程的加速,城市森林面临着严重的破碎化和外来物种侵入的危害[1,2],这些问题导致了一系列的负面的生态环境影响。城市森林在全球城市化景观中仍担任着提供重要生态系统服务的角色[3],而植物多样性对于一个地区生态系统的组成和稳定性具有关键的影响。坝光区域曾是深圳市生态保护最好的地区之一,这里植物资源丰富,古树成群,长期以来被誉为深圳“最美的渔村”“深圳的九寨沟”。许建新等[4]人对坝光的植物统计为109科285属376种,其中除去栽培植物的63种,野生植物为313种。梁鸿等[5]对坝光的25个植物群落进行调查,统计有植物317种,隶属于98科218属。保护好这里的植物资源与生态尤为重要。然而在近6年的国际生物谷经济开发区的开发建设中,如何平衡经济发展与生态保护成为了一个挑战,引起了社会各界和学术界的广泛关注。本研究以坝光区域为研究对象,通过对核坝路沿线的自然植物群落、半自然植物群落和人工植物群落进行调查研究和多样性分析,进一步了解和掌握该区域的植被组成和植物多样性特征,进而为坝光区域的植被与生态保护提供数据支持,为市政道路建设与当地生态系统保护相结合提供科学依据,实现经济发展与生态保护的平衡和可持续发展。
1 地点与方法
1.1 研究地概况
本研究地位于广东省深圳市大鹏新区大鹏半岛北端的坝光片区,西南临排牙山,东北与大亚湾海域相接,北与惠州交界。属于亚热带季风气候。年平均温度22.1 ℃,1月份均温13.9 ℃,极端最高温度36.6 ℃,极端最低温度1.4 ℃,年平均相对湿度79%,降水量1800.4 mm。
1.2 研究地点
本研究选取3个群落作为研究地,群落1为血桐-假鹰爪-海芋群落(Macaranga tanarius-Desmos chinensis-Alocasia macrorrhiza Comm.),属于自然林,位于东经114°33′34.9″,北纬22°39′05.26″,海拔24.00 m,群落样方总面积为1000 m2;群落2为台湾相思-银柴-山麦冬群落(Acacia confuse-Aporusa dioica-Liriope spicata Comm.),属于半自然林,位于东经114°34′01″,北纬22°39′07″,海拔为2.30 m,群落样方总面积为800 m2;群落3为荔枝-银柴-乌毛蕨群落(Litchi chinensis-Aporusa dioica-Blechnum orientale Comm.),属于人工林,位于东经114°31′51.10″,北纬22°39′00.93″,海拔为36.80 m,群落样方总面积为800 m2。
1.3 研究方法
1.3.1 样方调查方法
本研究采用样方调查法,在随机抽样调查过程中,以能够实现所调查范围全部含盖所研究每个群落的所有种类为原则,因此,结合少量典型调查法,以补充在随机调查法设置样方过程中极少数漏掉测定的种类。植物群落生态学的研究对象是植物群落,植物群落有特定的组成、结构、范围等8个特征[6],按照国际最小面积法原理,即当不同的地理区域所测植物群落的样方总面积达到一定数值时,再增加样方总面积数则该群落的种类数也不会再增加的原理,采用随机调查法与典型调查法结合的方法,则亚热带地区所需样方总面积为400 m2即可满足上述要求,超过此面积数在500~600 m2也可[7–11],在特殊情况下也可根据情况设置较小的样带,如在城市中可以设置1 m×10 m的典型样带[12],某些研究仅设一个样方,如在埃塞俄比亚对某木本植物群落研究中则采用20 m × 20 m的一个样方即代表一个植物群落的研究特征[13],遵义市喀斯特地貌区传统村落植被结构的研究中采用50 m × 20 m的样地[14],样地里再设置样方。虽然样方的面积不同,但所选取样方面积里测定及统计的平均指标是所研究群落特征的代表,选取样方总面积的大小对群落物种多样性特征的掌握没有影响[15,16]。
本研究在3个植物群落中,每个群落设8~10个10 m × 10 m乔木层样方,测定及记录样方内所有植物的物种名称、数量、高度、冠幅,乔木还测量其1.3 m处的胸径;灌木层样方在每个乔木大样方内设1~2个4 m×4 m的小样方,草本样方在每个灌木的样方内设2个1 m×1 m的小样方。乔木幼苗归入灌木层植物统计,即结合植物在深圳的生态习性,把植株高度大于4 m的乔木划分为乔木层,4 m以下的划分为灌木层[17]。
1.3.2 重要值计算方法
重要值计算方法参考张金屯《数量生态学第2版》中的方法[11]:
[乔木层重要值=(相对密度+相对频度+相对显著度)300]
[灌木层和草本层重要值=(相对密度+相对频度+相对盖度)300]
科和属的重要值为科属内所有物种重要值之和。
1.3.3 多样性计算方法
多样性计算方法采用[17–19]的方法。其中,文中的H为Shannon-Wiener指数、D为Simpson指数、C为生态优势度(Ecological dominance)、R1為Odum指数、R2为Menhinnick指数、R3为Marglef指数、J为Pielou均匀度指数、Df和Dg分别为科和属的多样性指数,Dc为科、属、种综合多样性指数、Rf1和Rf2为科的丰富度指数、Rg1和Rg2为属的丰富度指数[18–20]。
2 植物群落组成特征
2.1 群落1:血桐-假鹰爪-海芋群落(M.tanarius-D.chinensis-A.macrorrhiza Comm.)
该群落是自然植物群落。此群落乔木层中有共11种植物,优势种为血桐(0.30)(括号内数值为重要值,下同),次优种为龙眼(Dimocarpus longan,0.22);灌木层中种类较多,有34个种,优势种为假鹰爪(0.17),次优种为假苹婆(0.12);草本层中有23种,海芋(0.27)为优势种,次优种为剑叶凤尾蕨(0.11)。
此群落有33科52属66种,种的组成情况如下。
乔木层:血桐、龙眼、青果榕(Ficus variegata,0.14)、八角枫(Alangium chinense,0.08)、假柿木姜子(Litsea monopetala,0.06)、秋枫(Bischofia javanica,0.05)、黄皮(Clausena lansium,0.02)、荔枝(0.02)、朴树(Celtis sinensis,0.02)、潺槁木姜子(Litsea glutinosa,0.02)、白桐树(Claoxylon indicum,0.02)。
灌木层:假鹰爪、假苹婆、荔枝(0.10)、秋枫(0.08)、假柿木姜子(0.06)、九节(Psychotria rubra,0.05)、银合欢(Leucaena leucocephala,0.05)、龙眼(0.04)、红鳞蒲桃(Syzygium hancei,0.03)、罗汉松(Podocarpus macrophyllus,0.02)、朴树(0.02)、罗伞树(Ardisia quinquegona,0.02)、山小橘(Glycosmis pentaphylla,0.02)、异叶鹅掌柴(Schefflera diversifoliolata,0.02)、香港大沙叶(Pavetta hongkongensis,0.01)、八角枫(0.01)、鲫鱼胆(Maesa perlarius,0.01)、润楠(Machilus nanmu,0.01)、土沉香(Aquilaria sinensis,0.01)、紫玉盘(Uvaria microcarpa,0.01)、朱砂根(Ardisia crenata,0.01)、木瓜(Carica papaya,0.01)、青果榕(0.00)、秤星树(Ilex asprella,0.00)、蒲桃(Syzygium jambos,0.00)、粗叶榕(Ficus hirta,0.00)、苎麻(Boehmeria nivea,0.00)、狗骨柴(Diplospora dubia,0.00)、土蜜树(Bridelia tomentosa,0.00)、三桠苦(Evodia lepta,0.00)、毛果算盘子(Glochidion eriocarpum,0.00)、银柴(0.00)、血桐(0.00)、茜树(Aidia cochinchinensis,0.00)。
草本层:海芋、剑叶凤尾蕨、华南毛蕨(Cyclosorus parasiticus,0.08)、马唐(Digitaria sanguinalis,0.07)、淡竹叶(Lophatherum gracile,0.05)、半边旗(Pteris semipinnata,0.04)、芭蕉(Musa basjoo,0.04)、砂仁(Amomum villosum,0.04)、山麦冬(0.04)、傅氏凤尾蕨(Pteris fauriei,0.03)、五节芒(Miscanthus floridulus,0.02)、酢浆草(Oxalis corniculata,0.02)、火炭母(Polygonum chinense,0.02)、求米草(Oplismenus undulatifolius,0.01)、鞭叶铁线蕨(Adiantum caudatum,0.01)、鬼针草(Bidens pilosa,0.01)、粽叶芦(Thysanolaena maxima,0.01)、紫马唐(Digitaria violascens,0.01)、烏毛蕨(0.01)、香附子(Cyperus rotundus,0.01)、水蜈蚣(Kyllinga polyphylla,0.00)、鸭跖草(Commelina communis,0.00)、鳞花草(Lepidagathis incurva,0.00)。
此群落33科中,大戟科有58株植物数量最多,此科重要值为0.49;樟科数量第二,有40株植物,但此科中所有植物重要值仅为0.17,略低;无患子科和天南星科重要值分别为0.39和0.27。52个属中,木姜子属、苹婆属、血桐属、银合欢属和凤尾蕨属均有30多株植物,海芋属、龙眼属有20多株植物。血桐属重要值为0.31,其次是海芋属和龙眼属,分别为0.27和0.26。大戟科血桐属的重要值均最大,说明此群落中这一科属的植物在群落构建过程中占有重要地位。
2.2 群落2:台湾相思-银柴-山麦冬群落(A.confuse-A.dioica-L.spicata Comm.)
该群落为恢复多年的人工植物群落,属于半自然植物群落。此群落乔木层中共有16种植物,优势种为台湾相思(0.35),次优种为荔枝(0.09);灌木层中种类较多,有42个种,优势种为银柴(0.23),次优种为九节(0.11);草本层中有17种,山麦冬(0.22)为优势种,次优种为五节芒(0.17)。
此群落有40科55属65种,种的组成情况如下。
乔木层:台湾相思、荔枝、鹅掌柴(Schefflera octophylla,0.08)、八角枫(0.07)、山乌桕(Sapium discolor,0.06)、银合欢(0.05)、红鳞蒲桃(0.03)、铁冬青(Ilex rotunda,0.02)、乌桕(Sapium sebiferum,0.02)、椿叶花椒(Zanthoxylum ailanthoides,0.02)、簕欓花椒(Zanthoxylum avicennae,0.02)、黑面神(Breynia fruticosa,0.02)、野漆(Toxicodendron succedaneum,0.02)、土蜜树(0.02)、朴树(0.02)、牡荆(Vitex negundo,0.02)。
灌木层:银柴、九节、假鹰爪(0.05)、红鳞蒲桃(0.05)、秤星树(0.04)、假苹婆(0.03)、紫玉盘(0.03)、异叶鹅掌柴(0.03)、草珊瑚(Sarcandra glabra,0.03)、豺皮樟(Litsea rotundifolia,0.03)、米碎花(Eurya chinensis,0.03)、土沉香(0.02)、栀子(Gardenia jasminoides,0.02)、石斑木(Rhaphiolepis indica,0.02)、越南叶下珠(Phyllanthus cochinchinensis,0.01)、山乌桕(0.01)、朱砂根(0.01)、盐肤木(Rhus chinensis,0.01)、狗骨柴(0.01)、八角枫(0.01)、荔枝(0.01)、土蜜树(0.01)、破布叶(Microcos paniculata,0.01)、黑面神(0.00)、南烛(Vaccinium bracteatum,0.00)、毛棯(Melastoma sanguineum,0.00)、朴树(0.00)、龙眼(0.00)、毛果算盘子(0.00)、柳叶石斑木(Rhaphiolepis salicifolia,0.00)、大叶桂樱(Laurocerasus zippeliana,0.00)、牡荆(0.00)、粗叶榕(0.00)、酸藤子(Embelia laeta,0.00)、牛耳枫(Daphniphyllum calycinum,0.00)、簕欓花椒(0.00)、潺槁木姜子(0.00)、罗伞树(0.00)、蒲桃(0.00)、天门冬(Asparagus cochinchinensis,0.00)、杜英(Elaeocarpus decipiens,0.00)、马占相思(Acacia mangium,0.00)。
草本层:山麦冬、五节芒、乌毛蕨(0.11)、半边旗(0.09)、团叶鳞始蕨(Lindsaea orbiculata,0.08)、淡竹叶(0.05)、芒萁(Dicranopteris dichotoma,0.04)、艳山姜(Alpinia zerumbet,0.03)、华南毛蕨(0.03)、扇叶铁线蕨(Adiantum flabellulatum,0.03)、黑莎草(Gahnia tristis,0.01)、山菅(Dianella ensifolia,0.01)、毛果珍珠茅(Scleria herbecarpa,0.01)、毛果算盘子(0.01)、剑叶凤尾蕨(0.01)、异叶鳞始蕨(Lindsaea heterophylla,0.01)、韩信草(Scutellaria indica,0.01)。
此群落大戟科有227株植物,数量极多,此科的重要值为0.45;茜草科数量次之,但其重要值较低,仅为0.15;重要值其次分别为豆科(0.41)和百合科(0.24),豆科、百合科植株较少,均有20多株,其外形较大,故其重要值也较大。55个属中,金合欢属的重要值最大,为0.36;其次是银柴属,此属的植物数量也最多,多达261株,多为灌木所以其重要值低于金合欢属;山麦冬属重要值为0.22,此属仅有山麦冬,数量较多。此群落中大戟科银柴属的植物均为灌木层的银柴,大戟科的重要值在所有科中位于第一,银柴属重要值在所有属中位于第二,而豆科、金合欢属重要值在所有科和属中则分别位于第二和第一,说明此群落大戟科中包含属更多,遗传多样性高于豆科。
2.3 群落3:荔枝-银柴-乌毛蕨群落(L.chinensis-A.dioica-B.orientale Comm.)
此群落为人工植物群落,种植了大量荔枝,现已荒废。此群落乔木层共有23种植物,优势种为荔枝(0.48),次优势种为山烏桕(0.07);灌木层有46种,优势种为银柴(0.21),次优势种为九节(0.14);草本层有18种,优势种为乌毛蕨(0.23),次优势种为半边旗(0.16)。
此群落有42科61属74种,种的组成情况如下。
乔木层:荔枝、山乌桕、银柴(0.06)、簕欓花椒(0.04)、香蒲桃(Syzygium odoratum,0.03)、鹅掌柴(0.02)、朴树(0.02)、马尾松(Pinus massoniana,0.01)、鸭脚木(Schefflera octophylla,0.01)、楝叶吴萸(Evodia glabrifolia,0.01)、红鳞蒲桃(0.01)、豺皮樟(0.01)、毛八角枫(Alangium kurzii,0.01)、野漆(0.01)、土蜜树(0.01)、毛棯(Melastoma sanguineum,0.01)、乌材(Diospyros eriantha,0.01)、潺槁木姜子(0.01)、青果榕(0.01)、黄牛木(Cratoxylum cochinchinense,0.01)、石斑木(0.01)、竹节树(Carallia brachiata,0.01)、狗骨柴(0.01)。
灌木层:银柴(0.21)、九节(0.14)、蒲桃(0.13)、假鹰爪(0.05)、豺皮樟(0.03)、栀子(0.02)、石斑木(0.02)、异叶鹅掌柴(0.02)、草珊瑚(0.02)、狗骨柴(0.02)、假苹婆(0.02)、毛冬青(Ilex pubescens,0.01)、黑面神(0.01)、鹅掌柴(0.01)、毛果算盘子(0.01)、雅榕(Ficus concinna,0.01)、潺槁木姜子(0.01)、两粤黄檀(Dalbergia benthami,0.01)、粗叶榕(0.00)、土沉香(0.00)、红鳞蒲桃(0.00)、土蜜树(0.00)、荔枝(0.00)、秤星树(0.00)、罗伞树(0.00)、杂色榕(Ficus variegata,0.00)、日本杜英(Elaeocarpus japonicus,0.00)、假鱼骨木(Canthium dicoccum,0.00)、竹节树(0.00)、锡叶藤(Tetracera asiatica,0.00)、柞木(Xylosma racemosum,0.00)、野漆(0.00)、变叶榕(Ficus variolosa,0.00)、绒毛润楠(Machilus velutina,0.00)、白花灯笼(Clerodendrum fortunatum,0.00)、米碎花(0.00)、润楠(0.00)、紫玉盘(0.00)、羊角拗(Strophanthus divaricatus,0.00)、九里香(Murraya exotica,0.00)、朴树(0.00)、黄樟(Cinnamomum porrectum,0.00)、八角枫(0.00)、簕欓花椒(0.00)、罗汉松(0.00)、鲫鱼胆(0.00)。
草本层:乌毛蕨、半边旗、山麦冬(0.13)、箬竹(Indocalamus tessellatus,0.1)、金毛狗(Cibotium barometz,0.08)、异叶鳞始蕨(0.07)、团叶鳞始蕨(0.05)、扇叶铁线蕨(0.02)、黑莎草(0.02)、五节芒(0.01)、毛果珍珠茅(0.01)、类芦(Neyraudia reynaudiana,0.01)、华南毛蕨(0.01)、凤丫蕨(Coniogramme japonica,0.01)、傅氏凤尾蕨(0.01)、山菅(0.01)、姬蕨(Hypolepis punctata,0.01)、粽叶芦(0.01)。
此群落43科中无患子科的重要值最高,为0.49;其次是大戟科,大戟科数量极多,有236株;乌毛蕨科重要值位于第三,为0.23。61个属中,荔枝属重要值最大,为0.72,占有重要地位;其次是银柴属,此属植物也最多,有216株,主要是灌木层中有大量银柴,蒲桃属、荔枝属、九节属植物。荔枝全为乔木层的优势种,所以其对应的科和属重要值最高,也说明荔枝在自然环境下没有幼苗,其自然更新能力没有红鳞蒲桃和香蒲桃强,属于短期优势种群。蒲桃属仅有几株是乔木层植物,大部分为灌木层植物。
2.4 植物科属种分析
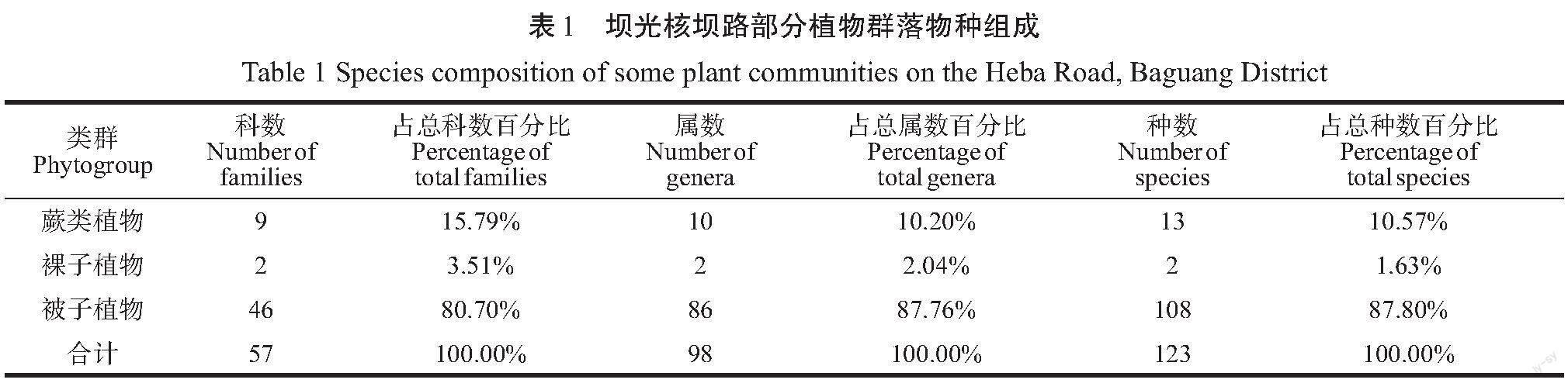
从表1可以看出,蕨类植物有9科,裸子植物2科,被子植物46科,合计57科,分别占总科数的15.79%、3.51%和80.70%。蕨类植物有10属,裸子植物2属,被子植物86属,合计98属,分别占总属数的10.20%、2.04%和87.76%。一共有蕨类植物13种,裸子植物2种,被子植物108种,分别占总种数的10.57%、1.63%和87.80%。
3 群落的植物多样性研究
3.1 植物群落的α-多样性
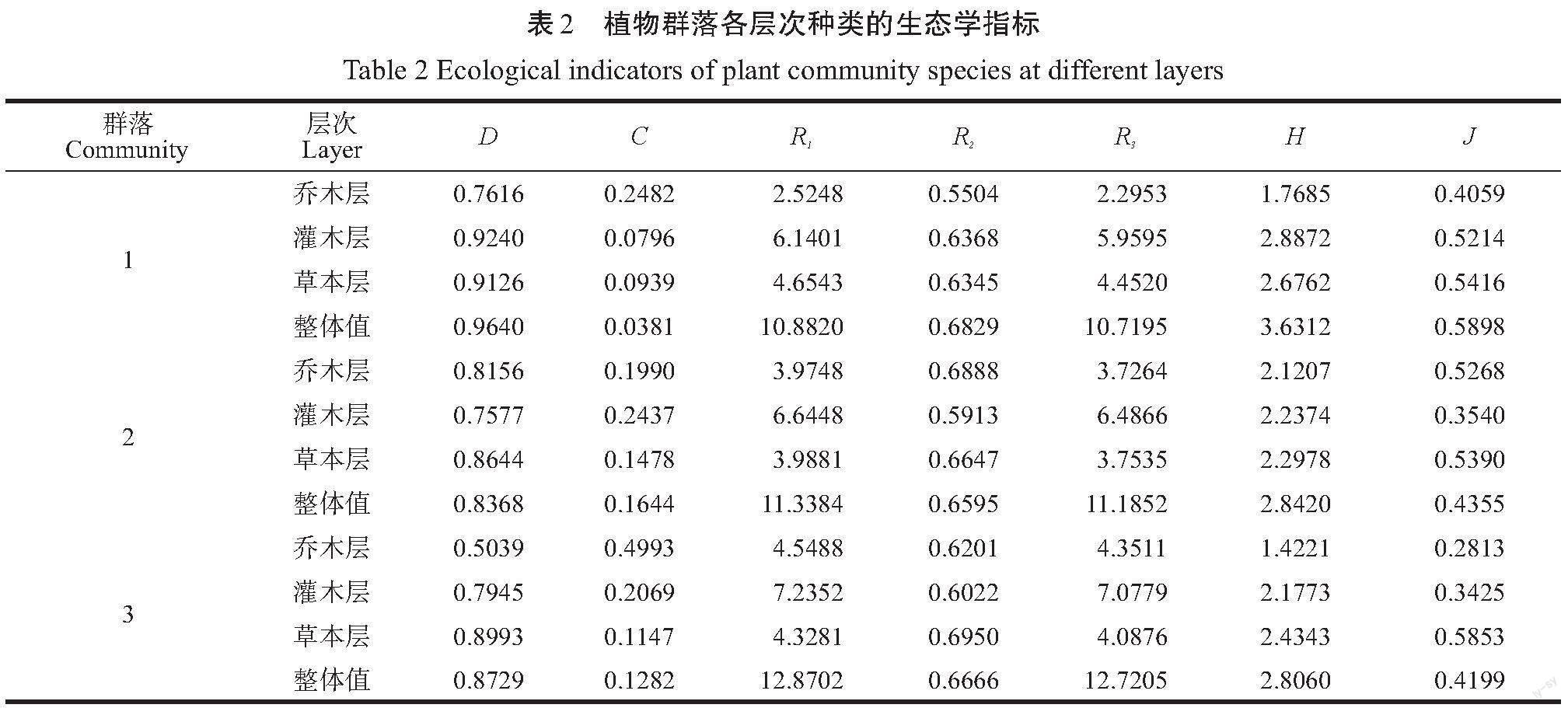
各植物群落物种多样性、丰富度及均匀度指数见表2。
从表2可以看出,群落1的乔木层多样性指数没有群落2高,但是其灌木层和草本层多样性均高于群落2;群落3乔木层多样性最低,从群落结构也可看出,此群落乔木层荔枝的重要值达0.48,是非常高的,导致乔木层多样性最低;灌木层和草本层中群落1多样性最高,群落3的多样性基本略高于群落2;在整体上,D值的排序为群落1>群落3>群落2,而H的排序为群落1>群落2>群落3,群落3的D值略高于群落2的主要原因可能是荔枝林荒废后其灌木和草本层有物种迁入。均匀度指数J值的排序也为群落1>群落2>群落3,同样是自然林高于半自然林,人工林最低。
从生态优势度指数来看,群落2的生态优势度指数整体上略高于群落3,而这两个群落又远高于群落1,说明自然群落的植物各种类的分布要比半自然群落和人工群落均匀。因为生态优势度指群落内某种类或极少数种类所占的优势程度的强弱,其值越高,则说明大部分种类的数量和分布等方面处于劣势或非常的劣势,因而群落的生物多样性是低或很低的。特别是群落3的乔木层,其生态优势度指数达到了0.4993,是自然群落的一倍以上。在群落2的灌木层中,生态优势度指数达0.2437,结合上文该群落灌木层科属种数量可以发现,此群落中银柴的株数约占总植株数的38.21%,说明群落2灌木层的物种分布很不均匀,这一结果与均匀度指数刚好相反。单就丰富度而言,科属种的丰富度指数基本上是群落3>群落2>群落1,因为丰富度的计算虽然也考虑了每个物种的个体数情况,但是相比各多样性指数和均匀度指数所含的这方面因素程度弱很多,因此,在此情况下,科属种的种类数均为群落3>群落2>群落1。但是从表2中可以看出,科属种的Simpson多样性指数排序为群落1>群落3>群落2,而种的Shannon-Weiner是群落1>群落2>群落3,均匀度指数也是群落1>群落2>群落3,这说明就多样性而言,自然群落>半自然群落>人工群落,在自然群落中不同物种个体分布更加均匀,遗传多样性也更高。
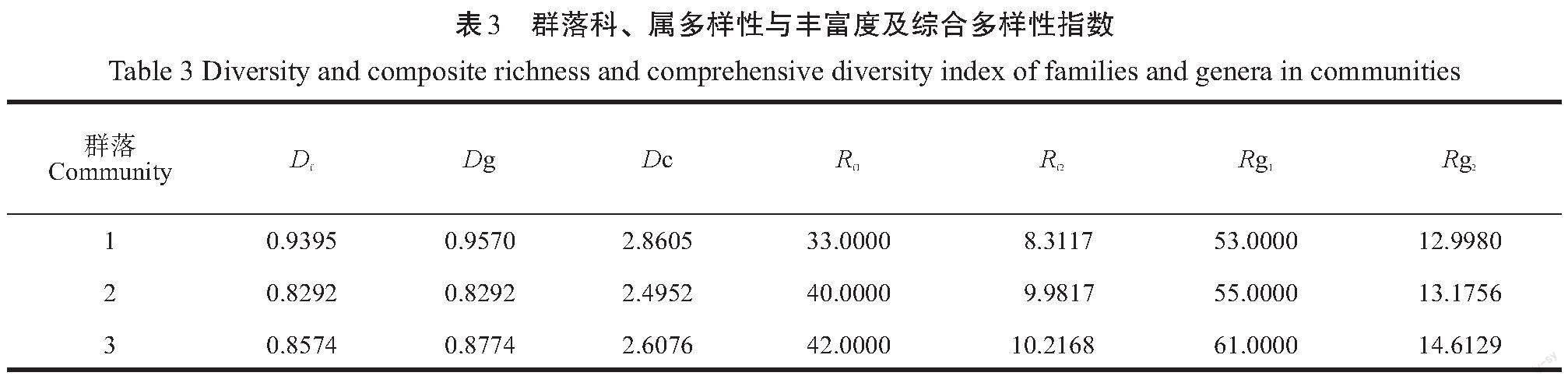
各植物群落科、属多样性与丰富度及综合多样性指数见表3。
從表3可以看出,科、属的多样性指数都是自然林的最高,而人工林的则居中,而综合多样性指数则是群落1>群落2>群落3,表现为即便考虑了科、属的多样性因素,还是自然林>半自然林>人工林。
3.2 植物群落β-多样性研究
从表4可以看出,各群落科的相似性系数在0.6133至0.7805之间,群落2和群落3科的相似性系数最高,达0.7805;最低是群落1和群落3,为0.6133,整体上比属的相似性系数要高。
各群落属的相似性系数在0.4860至0.6724之间,群落2和群落3属的相似性系数达0.6724,群落1和群落2属的相似性系数为0.4860。整体上比种的相似性系数要高。
群落2和群落3种相似性系数达0.5755,群落1和群落2的物种相似性系数为0.4640,群落1和群落3相似性系数最小,为0.4478,考虑为群落在人工干预之后有新的物种迁入,导致人工群落和自然群落之间的相似度较低。
群落1和群落3综合相似性系数最低,为0.5132;群落2和群落3综合相似性系数最高,达0.6647。这几个群落的综合相似性系数在0.5047至0.6647之间,部分群落的科属相似性系数高于0.6000,说明群落间有较多相同的科属。考虑到这三个群落的相隔距离较近,在相似性系数中融入科与属的要素,群落之间能有这样的差异也是较好的,说明这里的较大范围内的物种多样性和遗传多样性水平还是较高的。
4 讨论
4.1 物种组成特征
在坝光的这3个群落中,群落1为自然植物群落,群落2为半自然植物群落,群落3为人工植物群落。物种组成在一定程度上可以反映出植物群落的结构特点。这几个群落中,共有维管植物57科98属123种,其中蕨类植物9科10属13种,裸子植物2科2属2种,被子植物46科86属108种。在植物科的组成方面,大戟科在群落1和群落2中为优势科,无患子科在群落3中为优势科。在植物属的组成方面,这三个群落优势属分别为血桐属、金合欢属和荔枝属,其中群落2中的优势属不隶属于优势科,说明群落2中大戟科包含属的种类更多。在群落3科属种层面占据优势的分别为无患子科、荔枝属、荔枝,荔枝在乔木层中占有极大的优势地位,但是其在灌木层中则无,说明人工种植的荔枝在无人管理的情况下其自然更新能力极差,可以认为是一种短期优势种群。
4.2 物种多样性特征
在种类水平的Simpson 指数和Shannon-Weiner指数方面,多样性指标的排序均为群落1>群落2>群落3,说明这三个群落的多样性水平是自然林>半自然林>人工林,这与前文的研究一致[17,18,21];但从丰富度指标看群落3>群落2>群落1,而各群落的植物种类数也是如此。群落3的植物种类数大于群落2,群落2的植物种类数会略多于群落1,从群落3科、属包含的植株数量可以看出,群落3中大戟科植物数量最多,有236株,基本上均为灌木层中的银柴,大戟科的植物占总植株数量的29.57%,占比相当高,但是大戟科重要值位于第二,乔木层中无患子科的荔枝占乔木层植物数量的70.89%,占比极大,无患子科重要值位于第一;从群落2的科、属的所包含的植物株数也可以看出,群落2中大戟科有多达227株且均为银柴,占此群落总株数的38.21%;群落1中也是大戟科植株数量最多,共58株,包含了白桐树属、秋枫属、算盘子属、土蜜树属、血桐属、银柴属等6个属,大戟科的植株数量仅占群落1的25.55%,说明群落1虽然物种的种类数略少于群落2和群落3,但是其数量分布较为均匀,没有特别的单优物种。这也表明,虽然说群落3和群落2的丰富度要略高于群落1,但是因为这两个群落植物的数量分布不均匀,表现出了少部分种类的个体数占据着大量的比例,即其生态优势度指标过高,而丰富度指标虽然也考虑的各种类个体数的因素,但其为所有多样性指数里所占分量最轻的一类,有的丰富度指数还仅仅是指种类的数量,而上述的Simpson指数和Shannon-Weiner指数及均匀度指数等则融入各种类个体数的成分比重高很多,所以表现出了群落2和群落3的多样性并没有自然群落的群落1高。
另外從均匀度指标看,其排序也为群落1>群落2>群落3,与Simpson指数和Shannon-Weiner指数指标值的顺序一样,也进一步印证了自然群落的多样性高于半自然群落,半自然群落高于人工群落,这与黄玉源等、江小蕾等、毛志宏等人的研究结果一致[21-23]。
几个群落做比较,如果种的多样性各指数相同,则某一个群落有比其他群落多出较多乃至几倍的科与属,则这个群落的组成的遗传差异更大,其多样性的水平比其他群落的高出很多。因此,只比较各群落的种水平的多样性指标是不全面,不够合理的。因此,考虑各群落科、属层面的多样性,可以更加实际、更能全面展示群落多样性特征和水平[20]。从科、属、种的Simpson多样性指数来看,高低排序均为群落1>群落3>群落2;综合多样性指数的高低顺序也是群落1>群落3>群落2;而科属种的丰富度指数排序均为群落3>群落2>群落1,说明自然林的丰富度虽然没有人工林高,但是由于其物种的个体数分布更加均匀,因此自然林在科与属的等级方面,其多样性都是比人工林高。人工群落的丰富度较高可能是因为在人工群落荒废早期,林下空间较多,有较多对中小尺度资源利用率较高的灌木和草本迁入,这些植物会暂时增加群落内植物的物种数量,表现出丰富度略高,这种荔枝林的更新能力也较弱,从长期来看,其属于短期优势种,最终会被其他物种取代。物种多样性不能光要看其物种数量,还要看每个种类个体数量的比例分布是否均匀。如群落3,在荔枝林荒废初期,乔木层荔枝占据了明显的优势地位,导致乔木层其他植物的个体数等指标很低,群落的多样性不高;而随着群落的演替,群落3中灌木层和草本层有较多孤数异种植物,尤其是灌木层和草本层,这就表现出了群落的丰富度较高,但是比半自然群落的群落2也仅多了4个种,比自然群落的群落1也仅多了6个种。但如果在自然条件下,在长期演替的过程中,小尺度资源的竞争,部分偏阳生植物失去竞争优势,这些植物的迁出,使得群落的丰富度没有演替初期高,但是其物种个体数比例的分配会更加均匀,在分布上也会更加均匀。如群落2,经过了自然演替,其多样性比群落3要高。群落3科和属的Simpson多样性指数指数高于群落2,这可能也是因为荔枝林荒废后,在具有较多空间的林地中,不同科属的灌木和草本植物更容易迁入导致。
在一些研究中,自然群落和人工群落哪种群落结构好、多样性高有一定的争议,有些研究认为经过人工干扰后的其多样性变高[24–26]。但是通过研究可以发现,这类干扰后多样性升高主要有两个方面的原因:一方面这类研究多为1至2年,研究时间短,只能说是多样性暂时有所升高;一方面则是群落内主要为灌木层和草本层多样性有所升高,灌木层和草本层的波动很容易受到外界影响,短期的波动并不能说明多样性长期内都会升高,干扰导致的丰富度高只是一个短暂的特性[27,28]],而随着群落的演替,许多新进入的种类会在原来的种类的发育和遮蔽下而退出群落,其多样性又恢复到被间伐或片伐前的水平,但是这样严重影响了群落结构的稳定和生物量的保持,相当多的乔木和灌木被砍伐,会导致一系列生态效益的下降,新迁入的幼苗其生态的效益较低。多样性维护不仅指固定物种的共存,而且指在较长时间内维持物种丰富度和均匀性,因此需要考虑物种形成和灭绝率以及不频繁的殖民化[29],因此,上述暂时群落多样性的增高是得不偿失的做法[5,17,21]。上述所指的是把自然林或较好的半自然林或草地经人为强烈干扰和砍伐对其生物多样性的负面影响,而从另一个角度来看,人工林经过人为的林分改造,让其能够接纳野外的自然种类进入而生长发育,则群落的多样性会较明显地提高。多个间伐实验表明,通过对杉木林和马尾松林的间伐,将原来的人工林向自然林转变,经过5到10年,其多样性会有所提高[30,31]。韩国国家公园人工林通过大量间伐,经过5年演替,物种组成类似于自然林,多样性也极大提高[32]。Widenfal通过对针叶林间伐的分析,也建议通过间伐那些密度较大的人工林来提高其生物的多样性[33]。人工林需要通过间伐的手段推动其向自然林演替,从而提高原人工林的多样性,这也更进一步说明人工林的多样性明显低于自然林。反之亦然,自然林经过干扰,其多样性则会降低。对库尔德斯坦省(伊朗)的自然森林施加干扰,总体上植物其多样性会降低[34]。人为干扰和森林碎片已经改变了埃塞俄比亚北部森林的结构和组成,导致多样性的减少[35]。在本研究中,群落3为人工林,群落2经过多年演替从人工林转为半自然林,群落1为自然林,结果也证明了自然林的多样性高于半自然林,半自然林高于人工林。
这3个群落的科、属、种的综合相似性系在0.5047至0.6647,对于考虑了科与属的成分,其多样性水平属于中等的状态。与莲花山的几个群落相比[17],这3个群落的综合相似性系数还是稍低一些,说明这3个群落间的物种差异比莲花山的要大。杨梅坑和赤坳的综合相似性系数在0.1920至0.4490[18],总体上比本研究中的相似性系数要低一些,特别是杨梅坑的自然群落之间,其相似性系数更低,说明本研究中的物种没有自然群落中差异大。尽管本研究的几个群落相隔较近,它们之间的种类相似性水平仍然较低,表现出其在较大区域范围内的多样性还是较高,表现出一定的群落组成的隔离性。
参考文献
[1] FULLER D O. Forest fragmentation in Loudoun County, Virginia, USA evaluated with multitemporal Landsat imagery[J]. Landscape Ecology, 2001, 16(7): 627-642.
[2] DUGUAY S, EIGENBROD F, FAHRIG L. Effects of surrounding urbanization on non-native flora in small forest patches[J]. Landscape Ecology, 2007, 22(4): 589-599.
[3] SENA K L, HACKWORTH Z J, MAUGANS J, et al. Twenty years of urban reforestation: Overstory development structures understory plant communities in Lexington, KY, USA[J]. Sustainability, 2023, 15(3): 1985.
[4] 许建新, 钱瑭璜, 冯志坚, 等. 深圳坝光精细化工园区植物资源及植被类型分析[J]. 福建林业科技, 2012, 39(1): 108-111.
[5] 梁鸿, 许斌, 温海洋, 等. 深圳坝光国际生物谷规划区域生态状况与保护策略研究(上篇): 坝光区域陆地植被生态学特征[J]. 环境与可持续发展, 2017, 42(3): 7-14.
[6] 牛翠娟, 娄安如, 孙儒泳, 等. 基础生态学[M]. 3版. 北京: 高等教育出版社, 2015: 148.
[7] 高贤明, 黄建辉, 万师强, 等. 秦岭太白山弃耕地植物群落演替的生态学研究 Ⅱ演替系列的群落α多样性特征[J]. 生态学报, 1997, 17(6): 619-625.
[8] 朱彪, 陈安平, 刘增力, 等. 广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局[J]. 生物多样性, 2004, 12(1): 44-52.
[9] 何东进, 洪滔, 胡海清, 等. 武夷山风景名胜区不同森林景观物种多样性特征研究[J]. 中国生态农业学报, 2007, 15(2): 9-13.
[10] 黄金燕, 周世强, 谭迎春, 等. 卧龙自然保护区大熊猫栖息地植物群落多样性研究: 丰富度、物种多样性指数和均匀度[J]. 林业科学, 2007, 43(3): 73-78.
[11] 张金屯. 数量生态学[M]. 2版. 北京: 科学出版社, 2011: 20.
[12] 邱园, 冯志坚, 翁殊斐. 广州滨水绿地自生植物群落调查与园林应用潜力分析[J]. 广东园林, 2022, 44(6): 18-22.
[13] ANAMO A, MAMMO S, TEMESGEN M. Floristic composition and community analysis of woody species in Hereje Natural Forest, southwest Ethiopia[J]. SN Applied Sciences, 2022, 5(1): 48.
[14] DENG C J, HUANG Z S, ZHANG X J, et al. Correlation between vegetation structure and species diversity in traditional villages in Karst topographic regions of the Zunyi city, China[J]. Plants, 2022, 11(22): 3161.
[15] 刘灿然, 马克平, 于顺利, 等. 北京东灵山地区植物群落多样性的研究 Ⅳ.样本大小对多样性测度的影响[J]. 生态学报, 1997, 17(6): 601-610.
[16] 余欣繁, 黄玉源, 曾清怀, 等. 深圳坝光区域自然林及半自然林植物多样性比较[J]. 亚热带植物科学, 2021, 50(5): 388-393.
[17] 黄玉源, 余欣繁, 梁鸿, 等. 深圳莲花山植被组成及植物多样性研究[J]. 农业研究与应用, 2016(2): 18-32, 34, 33.
[18] LIANG H, HUANG Y Y, LIN S Z, et al. Study on plant diversity of mountain areas of Yangmeikeng and Chiao, Shenzhen, China[J]. American Journal of Plant Sciences, 2016, 7(17): 2527-2552.
[19] 黄玉源, 招康赛, 杨立君. 深圳山地植物群落结构与植物多样性[M]. 北京: 科学出版社, 2017: 21-22.
[20] PIELOU E C. Ecological diversity[M]. New York: Wiley, 1975.
[21] 黄玉源, 余欣繁, 招康赛, 等. 深圳小南山与应人石山地植物多样性比较研究[J]. 广西植物, 2016, 36(7): 795-805.
[22] 江小蕾, 张卫国, 杨振宇, 等. 不同干扰类型对高寒草甸群落结构和植物多样性的影响[J]. 西北植物学报, 2003, 23(9): 1479-1485.
[23] 毛志宏, 朱教君. 干扰对植物群落物种组成及多样性的影响[J]. 生态学报, 2006, 26(8): 2695-2701.
[24] 牛莉芹, 程占红. 五台山森林群落中物种多样性对旅游干扰的生态响应[J]. 水土保持研究, 2012, 19(4): 106-111.
[25] 周光兵, 龙翠玲. 人为干扰对城市近郊区森林植物多样性的影响: 以贵阳市乌当区为例[J]. 西南师范大学学报(自然科学版), 2011, 36(5): 98-103.
[26] 郭子良, 刘欣艳, 张曼胤, 等. 大九湖湿地游步道两侧草本植物群落對旅游干扰的响应[J]. 生态科学, 2022, 41(6): 82-91.
[27] CONNELL J H. Diversity in tropical rain forests and coral reefs[J]. Science, 1978, 199(4335): 1302-1310.
[28] SHEA K, ROXBURGH S H, RAUSCHERT E S J. Moving from pattern to process: Coexistence mechanisms under intermediate disturbance regimes[J]. Ecology Letters, 2004, 7(6): 491-508.
[29] CHESSON P. Mechanisms of maintenance of species diversity[J]. Annual Review of Ecology and Systematics, 2000, 31: 343-366.
[30] 李瑞霞, 馬洪靖, 闵建刚, 等. 间伐对马尾松人工林林下植物多样性的短期和长期影响[J]. 生态环境学报, 2012, 21(5): 807-812.
[31] 徐雪蕾. 间伐对杉木人工林的生长调控作用研究[D]. 北京: 北京林业大学, 2020.
[32] CHO S, MYEONG H H, CHOUNG Y. Promotion of plant species diversity of artificial plantations in Korean National Parks through thinning[J]. Journal of Asia-Pacific Biodiversity, 2020, 13(4): 631-636.
[33] WIDENFALK O, WESLIEN J. Plant species richness in managed boreal forests—Effects of stand succession and thinning[J]. Forest Ecology and Management, 2009, 257(5): 1386-1394.
[34] ESHAGHI R J, VALADI G, SALEHZADEH O, et al. Effects of anthropogenic disturbance on plant composition, plant diversity and soil properties in oak forests, Iran[J]. Journal of Forest Science, 2018, 64(8): 358-370.
[35] MULUNEH M G, FEYISSA M T, WOLDE T M. Effect of forest fragmentation and disturbance on diversity and structure of woody species in dry Afromontane forests of northern Ethiopia[J]. Biodiversity and Conservation, 2021, 30(6): 1753-1779.
