鸭梨PbADC 基因的鉴定及在果心褐变中的表达
2023-12-16李庆秀寇晓敏吴海清梁丽雅闫师杰
李庆秀,邓 冰,寇晓敏,李 萍,吴海清,4,梁丽雅,闫师杰,4*
(1 天津农学院 食品科学与生物工程学院 天津300384 2 山西农业大学食品科学与工程学院 山西晋中030801 3 天津农学院 基础科学学院 天津300384 4 天津市农副产品深加工技术工程中心 天津300384 5 天津农学院 农学与资源环境学院 天津 300384)
鸭梨(Pyrus bretschneideri Rehd cv.Yali)属于蔷薇科白梨品系的一种,是我国最早的出口商品之一[1-2]。鸭梨作为我国出口梨的主打品种,以其鲜嫩多汁、酸甜可口、香气浓郁、耐贮藏等优点而备受消费者青睐,然而其在贮藏期间极易发生逆境伤害,导致果实内部褐变,严重降低了果实的贮藏质量和食用口感,从而造成严重的经济损失[3-4],因此解决鸭梨褐变问题十分必要。
有研究显示,内源多胺在植物细胞中的累积水平越高,对植物的胁迫抗逆性就越强,而多胺合成酶基因表达水平的上调是其累积的先决条件[5-6]。多胺(Polyamines,PAs)是一类含有2 个或更多氨基组成的带正电荷的小分子化合物,在生物体内普遍分布[7-10]。高等植物体内的主要多胺有腐胺、亚精胺和精胺,它们对植物的生长发育及逆境响应起着非常重要的调控作用[11-14]。当果蔬受到逆境伤害时,其游离态腐胺会转变成亚精胺和精胺,从而得到较高含量的游离态亚精胺和精胺来抵御逆境胁迫及自身衰老过程中的一系列伤害[15]。王颉等[16]研究表明外源多胺处理可以显著降低鸭梨果心组织CO2含量,抑制其褐变的发生。多胺的生物合成主要有两条途径,一条是通过鸟氨酸脱羧酶(ODC)将鸟氨酸转化为腐胺;另一条精氨酸脱羧酶(Arginine decarboxylase,ADC)催化精氨酸经脱羧和酶促反应后,再形成腐胺(Putrescine,Put)。ADC 是植物在逆境胁迫条件下合成多胺的关键限速酶,能够将精氨酸转化为Put,该途径为高等植物所独有,其结构与功能备受关注[17-20]。精氨酸脱羧酶(ADC)是依赖磷酸吡哆醛(PLP)的一类酶,广泛存在于微生物和动植物中,且ADC 一般存在于植物的细胞质中,对外界各种胁迫反应都十分敏感。目前研究发现ADC 在植物抗逆、根系发育及细胞伸长等方面具有重要的功能,而其在果蔬,特别是鸭梨等果蔬采后褐变中的功能尚未见报道。
本研究以鸭梨果心为材料,从鸭梨转录组数据中筛选并鉴定PbADC 基因,通过生物软件来预测其基本生物学特性,并利用qRT-PCR 技术对其在鸭梨采后贮藏期间的表达模式进行分析。通过以上研究为探索PbADC 基因在鸭梨采后果心褐变中的调控机制提供依据,为进一步解析鸭梨果心褐变机理奠定分子和理论基础。
1 材料与方法
1.1 试验材料
鸭梨于2020 年采自河北省辛集市,采摘时间为8 月26 日(盛花期后145 d),即早采鸭梨:挑选大小均一、无机械伤和病虫害的果实,采摘后当天运回冷库,预冷24 h 后进行急速降温【直接放于(0±0.5)℃】和缓慢降温处理【先放入12 ℃的冷库中,每5 d 降2 ℃,30 d 降到(0±0.5)℃】,每个处理各10 箱。分别在贮藏0,30,60,90,120,150,180 d 和210 d 进行果心组织取样,用液氮将所有样品速冻后置于-80 ℃超低温冰箱中保存。
1.2 试验设备与仪器
TAdvanced 96 温度梯度PCR 仪,德国耶拿分析仪器公司;Quantagene q225 荧光定量PCR 仪,北京酷博科技有限公司;GenSens 2100 凝胶成像仪,上海勤翔科学仪器有限公司;PowerPac Basic 1645050 电泳仪,美国伯乐公司;Eppendorf 5418小型台式高速离心机,德国艾本德股份公司;NanoDrop one 超微量分光光度计,美国赛默飞世尔科技公司。
1.3 试验方法
1.3.1 基因鉴定 参考韩云云等[21]的方法,并略作修改。利用在线软件SMART 和Pfam 对从转录组数据中筛选的基因进行注释,使用NCBI 中的Blast P 分析ADC 蛋白的保守结构域,基因结构用IBS 1.0 软件进行可视化。
1.3.2 系统发育树构建及多序列比对 PbADC蛋白和其部分同源序列使用NCBI 数据库中的BLAST 检索并下载,系统发育树使用MEGA 6.0软件进行构建,PbADC 蛋白与其亲缘关系较近的ADC 蛋白使用DNAMAN 软件进行多序列比对。
1.3.3 蛋白生物信息学分析 参考韩云云等[21]的方法对PbADC 蛋白进行生物信息学分析。

表1 蛋白生物信息学分析软件Table 1 The software of protein bioinformatics analysis
1.3.4 果心褐变指数的测定 参考闫师杰等[22]的方法,并略作修改;分别在贮藏0,30,60,90,120,150,180 d 和210 d 随机从急降和缓降处理的10箱鸭梨中各选取30 个鸭梨,每组10 个,把鸭梨切开后统计其果心子房室的褐变情况。褐变指数计算方法见式(1)。
1.3.5 总RNA 提取和相对表达量测定 参考张引引[2]和樊晓岚等[23]的方法,略作改动。
1.3.5.1 总RNA 提取和反转录 根据植物多糖多酚试剂盒说明书进行鸭梨果心组织样品总RNA 的提取;制备1%的琼脂糖凝胶,并用水平电泳检测RNA 提取的质量;采用Nanodrop 超微量分光光度计测定RNA 在不同波长下(260 nm 和280 nm 处)的OD 值,来确定RNA 的浓度及纯度。使用宝生物的反转录试剂盒(RR047A)将1 μL 的总RNA 为模板进行反转录,合成cDNA(具体操作参照说明书),反应结束后保存于-20 ℃冰箱中备用。
1.3.5.2 相对表达量测定 将获得的cDNA 参照确定的基因序列用NCBI blast primer 设计用于荧光定量PCR 反应的引物(表2)。参照试剂盒说明书把荧光定量PCR 反应体系充分混匀后,放于荧光定量PCR 仪中进行反应,反应结束后根据得到的溶解曲线和扩增曲线确定数据的可靠性,并进行计算。将PbActin(登录号GU830958.1)作为内参基因,用降温前(贮藏0 d)样品中的相关基因表达量作为标准,使用2-△△Ct方法计算出不同样品的基因相对表达量[21]。

表2 本研究所用引物Table 2 Primers used in this study
1.3.6 精氨酸脱羧酶(ADC)活性的测定 采用植物精氨酸脱羧酶试剂盒进行精氨酸脱羧酶(ADC)活性的测定。
1.3.7 数据统计及分析 利用Excel 2016 和IBM SPSS 统计软件进行数据处理和方差分析,使用Origin 进行绘图。
2 结果与分析
2.1 PbADC 基因鉴定
从鸭梨转录组数据中得到1 条与ADC 基因具有同源性的编码序列,通过SMART 在线工具对该序列所编码的蛋白进行结构域分析,结果表明PbADC 蛋白第 127~399 位氨基酸残基为Orn_Arg_deC_N 结构域(图1),再结合CDD 注释结果确定该序列为PbADC 基因的编码序列。而后对PbADC 基因的DNA 序列进行分析,结果显示该基因DNA 序列长度为3 020 bp,其最长开放阅读框为2 193 bp,编码730 个氨基酸。
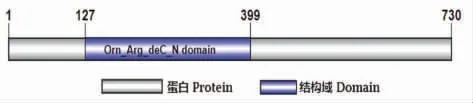
图1 PbADC 基因结构域预测Fig.1 Domain prediction of PbADC gene
2.2 PbADC 氨基酸序列比对和系统发育树构建
为分析PbADC 与其它植物ADC 蛋白的进化关系,通过MEGA 6.0 软件,用ML 法进行构建鸭梨(Pyrus bretschneideri)、杜梨(Pyrus betulifolia)、苹果(Malus domestica)、秋子梨×西洋梨(Pyrus ussuriensis × Pyrus communis)、湖北海棠(Malus hupehensis)、桃(Prunus persica)、月季(Rosa chinensis)、葡萄(Vitis vinifera)、雷公藤(Tripterygium wilfordii)、枣(Ziziphus jujuba)、川桑(Morus notabilis)、栓皮槠(Quercus suber)、核桃(Juglans regia)、木薯(Manihot esculenta)ADC 蛋白系统发育树。结果(图2)表明,PbADC 蛋白与杜梨、秋子梨×西洋梨、苹果和海棠ADC 亲缘关系较近。同时使用DNAMAN 软件对PbADC、杜梨、秋子梨×西洋梨、苹果和湖北海棠ADC 蛋白的氨基酸序列进行多重序列比对分析,结果(图3)显示其整体一致性为96.66%,且均含一个完整的Orn_Arg_deC_N结构域,并具有2 个关键的酶活位点:氨基末端的赖氨酸(K)和羧基末端的半胱氨酸(C),并在第151~169 位残基发现精氨酸脱羧酶家族2-磷酸吡哆醛结合位点。

图2 PbADC 蛋白与部分物种ADC 蛋白的系统发育树Fig.2 Phylogenetic tree of PbADC protein and ADC protein of some species

图3 PbADC 与其它同源蛋白的序列比对Fig.3 Sequence alignment of PbADC with other homologous proteins
2.3 PbADC 蛋白生物信息学分析
2.3.1 PbADC 理化性质分析 采用在线软件ProtParam 预测了PbADC 的理化性质。PbADC 氨基酸残基为730,其中甘氨酸(Gly,9.60%)、亮氨酸(Leu,10.10%)、丝氨酸(Ser,9.30%)、缬氨酸(Val,8.10%)和丙氨酸(Ala,8.60%)是该蛋白中最主要的几种氨基酸;分子式为C3463H5406N940O1063S37,分子质量为78.403 ku;含正电(Arg+Lys)的氨基酸数58 个,含负电(Asp+Glu)的氨基酸数87 个;理论等电点为5.23,不稳定系数为42.64(大于40),属于酸性不稳定蛋白。
2.3.2 PbADC 蛋白二、三级结构分析 使用SOPMA 和SWISS-MODEL 在线工具分析并预测了PbADC 的二、三级结构,结果显示PbADC 蛋白主要由α-螺旋和无规则卷曲构成,其中α-螺旋占42.05%、无规则卷曲占35.48%、延伸链和β-转角分别占15.48%和6.99%(图4a 和4b)。

图4 PbADC 蛋白的二、三级结构预测Fig.4 The prediction of secondary structure and tertiary structure of PbADC protein
2.3.3 PbADC 蛋白亲水性分析 使用ExPASy-ProtScale 在线工具分析了PbADC 的亲水性,结果表明PbADC 蛋白表面存在722 个亲疏水基团,以第188 位氨基酸残基(Met)的疏水能力最强,为2.567;在第143 位(Tyr)疏水能力最弱,为-2.111(图5)。从总体上看,PbADC 蛋白表面的总亲水位点数量(375 个)大于总疏水位点(347 个),且总平均亲水系数为-0.044,说明该蛋白为亲水蛋白。
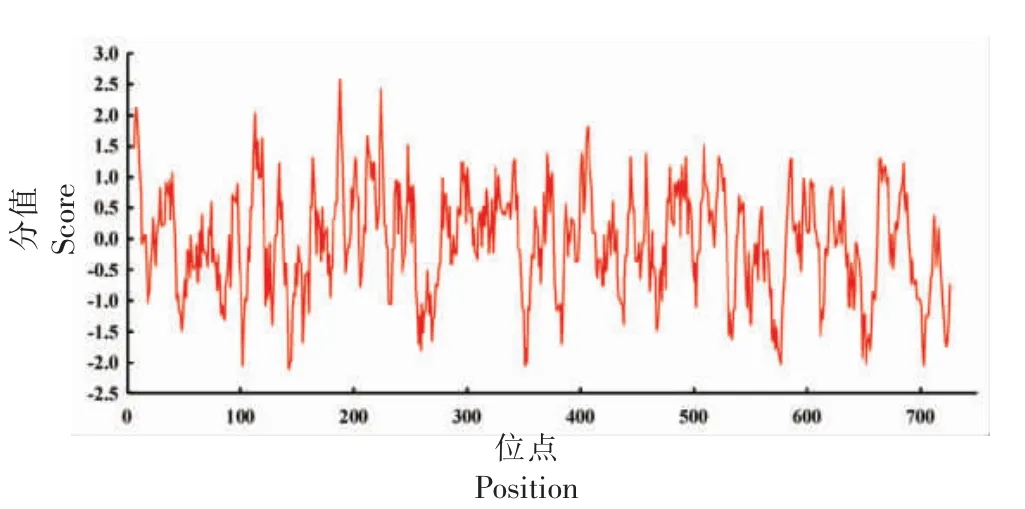
图5 PbADC 蛋白的亲水性分析Fig.5 Hydrophilic analysis of PbADC protein
2.3.4 PbADC 蛋白细胞定位与信号肽分析 利用SignalP 4.1 Server 对PbADC 蛋白的信号肽进行了预测,发现前60 位氨基酸中最高的信号肽、初始剪切位点和综合剪切位点的分值均低于0.5(图6a),且信号肽预测的结果显示为no,表明不存在信号肽。蛋白质的跨膜区域主要是指膜内在蛋白和细胞膜的膜脂相结合的部位,为了寻找跨膜结构域,用TMHMM Server 在线工具对PbADC蛋白进行了预测,结果表明该蛋白的跨膜螺旋数量为0,没有跨膜结构(图6b)。综上所述,推测PbADC 为一种非分泌蛋白,不会进入其它细胞器发挥作用。利用PSORT 和Cell-Ploc 在线软件对PbADC 在细胞中的定位情况进行了预测,结果发现PbADC 蛋白主要存在于细胞质中。

图6 PbADC 蛋白的细胞定位与信号肽分析Fig.6 Cell localization and signal peptide analysis of PbADC protein
2.4 PbADC 基因在鸭梨采后褐变过程中的表达模式分析
果心褐变指数是显示褐变严重程度的一个重要指标,它既可看出整体果实的褐变情况,也可看出单个果心的褐变程度。不同处理组鸭梨果心的褐变指数随贮藏时间的增加而增加(图7a)。在贮藏120 d 时,急降和缓降处理组的鸭梨均出现了不同程度的果心褐变,褐变指数分别为0.217 和0.167。急速降温和缓慢降温两个处理组相比,急速降温处理组的果心褐变指数均高于缓慢降温处理组(P<0.05),说明急速降温处理加速了鸭梨的果心褐变。

图7 PbADC 基因在鸭梨采后褐变过程中的表达模式分析Fig.7 Expression pattern analysis of PbADC gene during postharvest browning of Yali pear
精氨酸脱羧酶(ADC)作为多胺合成通路中的一个重要酶,对外界环境的胁迫响应最为敏感[24]。结果(7b)表明,在贮藏的前60 d,鸭梨ADC 基因上调表达,其中ADC 基因在缓降处理中的表达比急降处理高(P<0.05),并且缓降鸭梨的表达量最高为376.7。ADC 基因在鸭梨果心中的表达量虽随贮藏时间延长而降低,但仍是缓降鸭梨的表达量高,说明缓慢降温能促进鸭梨ADC 基因的表达,有助于其抵抗低温胁迫导致的果心褐变。精氨酸脱羧酶在不同处理下均呈先升后降的变化趋势,这与ADC 基因的表达情况基本一致(图7c)。在整个贮藏期间,缓慢降温处理下的ADC 酶活性均高于急速降温处理(P<0.05),且在贮藏60 d 时,ADC 酶活性最高,为68.89 U/L。结合鸭梨果心褐变情况分析,整个贮藏期缓慢降温处理组鸭梨果心褐变指数低,褐变情况少,可能是因为鸭梨果实中的ADC 酶活力较高,能够合成大量的多胺,从而提高了鸭梨的抗冷性,降低了果心褐变的风险。
3 讨论与结论
作为一种新型植物生长的调节剂或“第二信使”,多胺具有稳定细胞膜、参与植物成熟衰老过程以及响应逆境等多种功能[25]。ADC 属于磷酸吡哆醛依赖性酶(Pyridoxal 5'-phosphate de-carboxylase,PLPDE)基因超家族Ⅲ型的重要成员,其含有1 个PLPDE 典型的Orn_Arg_deC_N 结构域,是生物体内多胺生物合成的一个重要限速酶[17,26]。本研究发现鸭梨PbADC 基因序列最长ORF 为2 193 bp,编码730 个氨基酸。鸭梨与柑橘和草莓等其它果树具有相似的ADC 基因序列,柑橘PtADC 基因的ORF 含2 256 bp,编码751 个氨基酸[27];森林草莓FvADC 基因的ORF 含2 154 bp,编码717 个氨基酸[28]。PbADC 蛋白是定位于细胞质的非分泌亲水性不稳定蛋白,二级结构以α-螺旋和无规则卷曲为主,该结果与其它ADC 蛋白的细胞定位情况一致[27-29]。除此之外,本研究通过同源序列比对发现 PbADC 包含完整的Orn_Arg_deC_N 结构域,含有家族保守的2-磷酸吡哆醛结合位点和两个重要的酶活位点,这些与已有的一些物种的ADC 同源蛋白相吻合,是鉴定磷酸吡哆醛依赖性酶基因超家族Ⅲ型PLPDE 家族成员的关键结构域。
有研究表明,ADC 基因在植物抗逆中的作用较大。孙培培[30]研究表明,枳的PtADC 基因能够提高柑橘的低温耐受力,降低H2O2和超氧阴离子的积累。刘荣等[31]通过RACE 技术,从芒果中分离得到MiADC 基因,并发现其在转录水平上能够受到低温的调控,是一类对低温具有应答作用的基因。孙虹丽[32]在梨褐皮与多胺代谢的研究中发现花后ADC 基因的表达量先增加后降低,这一现象与多胺的变化趋势相吻合,是造成果皮褐色的主要因素。在贮藏初期,ADC 基因的表达量增加,其通过上调表达来提高对低温的抗性,从而降低了缓降鸭梨果心褐变的发生。
本研究在鸭梨转录组数据中筛选并鉴定到了1 条由PbADC 基因编码的序列,通过对该序列编码的蛋白进行生物信息学分析,明确了其生物学特性,并利用qRT-PCR 方法分析了PbADC 基因在鸭梨采后果心褐变中的表达模式。综上所述为进一步揭示鸭梨采后果心褐变发生机制奠定了理论基础。
