瘤胃中添加剂的甲烷减排作用及甲烷厌氧氧化
2023-12-04朱晓雯周振明
朱晓雯 刘 月 周振明 吴 浩
(中国农业大学动物科学技术学院,动物营养学国家重点实验室,北京 100193)
甲烷(CH4)是产量仅低于二氧化碳(CO2)的温室气体,占温室气体排放总量的16%。联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)指出,CH4的全球变暖潜能(global warming potential,GWP)值是CO2的28倍[1],具有捕获大气红外辐射的能力,加速全球变暖,对生态环境造成威胁。反刍动物作为温室气体的主要贡献者,其产生的温室气体占畜牧业总排放量的80%,约占全球人为CH4排放量的16%,其胃肠道发酵产生的CH4约占CH4排放总量的47%[2-3]。反刍动物CH4生成不仅加剧温室效应,还会损失2%~12%的饲料总能,造成饲料可消化能的浪费。因此,减少反刍动物CH4排放对提高饲料利用率、缓解温室效应都具有重要意义。
大量研究证明,在饲粮中加入不同类型添加剂可通过减少产甲烷菌(methanogen)数量[4]、抑制产甲烷菌活性[5-7]以及抑制关键酶活性[8]等方式降低CH4产量,但都存在一定局限性;而甲烷厌氧氧化作为自然环境中重要的甲烷汇,不仅CH4利用效率颇高,而且反应发生条件基本与瘤胃环境相符。有研究人员从微生物角度进行探究,从侧面说明甲烷厌氧氧化过程可能存在于瘤胃中,但未进行直接的过程验证。基于此,本文综述了不同添加剂对反刍动物CH4减排的效果以及甲烷厌氧氧化在瘤胃中的可能性,旨在为反刍动物CH4减排提供新思路。
1 反刍动物瘤胃中的产甲烷菌与CH4生成机制
1.1 产甲烷菌的种类及特点
产甲烷菌与细菌虽同为原核生物,但其16S rRNA序列中并不存在细菌特征序列,且进化树上与真菌的亲缘更近[9]。产甲烷菌严格厌氧,属水生古细菌门。目前已知的产甲烷菌可分为4个纲,包括7个目、14个科和35个属,其中甲烷杆菌属(Methanobacterium)、甲烷短杆菌属(Methanobrevibacter)、甲烷微菌属(Methanomicrobium)和甲烷八叠球菌属(Methanosarcine)广泛存在于反刍动物胃肠道中[10],这些产甲烷菌分布在瘤胃液、原虫表面、原虫胞内的氢化酶体、瘤胃壁以及其他微生物或饲料颗粒上[11]。不同反刍动物在不同生理阶段核心产甲烷菌群略有不同,但奶牛、肉牛、水牛和牦牛等的优势菌属都是甲烷短杆菌属,而具体菌种有所区别[12-14]。反刍动物瘤胃中产甲烷菌分类见图1。

图1 反刍动物瘤胃中产甲烷菌分类Fig.1 Classification of methanogens in rumen of ruminants
1.2 CH4生成机制
碳水化合物在反刍动物瘤胃中被微生物降解为单糖,单糖再进一步降解生成丙酮酸,继而发酵产生挥发性脂肪酸(volatile fatty acid,VFA),同时产生氢气(H2)。瘤胃中的H2有2种存在形式,分别为溶解H2和气态H2,产甲烷菌可以利用溶解H2产生CH4,该过程也称为氢(H)营养型途径;除此之外,瘤胃中还存在其他2种产CH4途径,即乙酸发酵型途径和甲基营养型途径,以上3种途径中,由甲烷短杆菌经CO2和H2还原途径产生的CH4占瘤胃CH4产量的82%[15]。
1.2.1 H营养型途径
H营养型途径是瘤胃中主要的产CH4途径,其中甲烷短杆菌属为主要的产甲烷菌[16]。H2为产CH4过程提供电子,将CO2还原为甲酰甲烷呋喃,然后与四氢甲烷蝶呤结合,形成甲酰四氢甲烷喋呤,在一系列酶的作用下被还原为甲基四氢甲烷蝶呤,再由辅酶M甲基转移酶将甲基转移,最终在甲基还原酶的作用下生成CH4。在此过程中,还有F420、F430、辅酶B和辅酶M的参与[17]。此外,甲酸脱氢酶能将甲酸转化为CO2和H2,CO2被还原产生CH4,该路径中能利用甲酸的产甲烷菌占18%[18]。H营养型途径图2[19]。

CO2:二氧化碳 carbon dioxide;H2:氢气 hydrogen;CH4:甲烷 methane;H+:氢离子 hydrogen ion。图2 H营养型途径Fig.2 H nutritional type pathway[19]
1.2.2 乙酸发酵型途径
产甲烷菌将乙酸分解生成的羧基(—COOH)和甲基(—CH3)分别氧化还原为CO2和CH4。参与反应过程的辅酶主要有辅酶A、辅酶M和辅酶B。此类途径产CH4占比较少,主要涉及的产甲烷菌是甲烷八叠球菌属和甲烷鬃菌属(Methanosaeta)。
1.2.3 甲基营养型途径
甲烷八叠球菌目(Methanosarcinales)、甲烷球菌属(Methanosphaera)、嗜甲基甲烷菌科(Methanomassiliicoccaceae)是该途径的主要产甲烷菌类型,其需要H2作为共基质生成CH4[20]。在此过程中,产甲烷菌利用甲醇、甲硫醇和甲胺等甲基化合物为底物生成CH4。
2 缓解反刍动物胃肠道CH4排放的策略
瘤胃CH4产量受动物个体、饲粮组成、瘤胃发酵模式、瘤胃微生物区系差异、饲养管理和环境温度等多方面因素影响。基于以上几点制定针对反刍动物的CH4减排方案,起到多种措施并用、共同发挥作用的目的。CH4减排途径见图3[21]。

CO2:二氧化碳 carbon dioxide;H2:氢气 hydrogen;CH4:甲烷 methane。图3 CH4减排途径Fig.3 CH4 emission reduction pathways[21]
饲料添加剂主要通过以下几种途径缓解瘤胃CH4排放:1)抑制产甲烷菌生长,降低其数量及活性,减少CH4生成;2)与产甲烷菌竞争H2,减少CH4转化;3)抑制产甲烷菌合成CH4途径中某些酶的活性,抑制CH4生成。
2.1 抑制产甲烷菌生长和活性
2.1.1 植物提取物
植物提取物是一类天然的、毒副作用低的、有生物活性的混合物,具有抗菌活性,有改善瘤胃发酵、提高生产性能、抑制CH4生成等功能,主要有植物精油、单宁和皂苷等。
植物精油是有芳香味的油状液体,分为萜类和丙苯素类[22],其活性物质大多为亲脂性,与产甲烷菌的细胞膜有高亲和力,可直接破坏膜蛋白,使细胞膜通透性增加,干扰产甲烷菌生长和活性。体外试验中,迷迭香精油能使CH4产量随剂量增加而线性下降,但对VFA和营养物质降解率无影响[23];不同植物[波叶海桐(Pittosporumundulatum)、金姜花(Hedychiumgardnerianum)和日本柳杉(Cryptomeriajaponica)]来源精油均能显著降低CH4排放量,但总VFA含量显著降低[24]。体内试验中,柑橘精油[25]能够降低湖羊瘤胃产甲烷菌丰度,精油混合物[5]能显著降低奶牛的CH4产量,且不影响干物质采食量和生产性能;大蒜油[26]、肉桂油[26]和牛至油[27]也能降低CH4产量但会对瘤胃发酵参数产生不利影响,使VFA含量降低。以上研究证明,植物精油能抑制瘤胃CH4生成,具有抗氧化等优点,但植物精油中活性成分较多,作用效果主要取决于来源和剂量;此外,还应考虑植物精油对瘤胃微生物发酵的影响,避免对动物健康和生产性能造成负面影响,因此需进一步研究确定在实际生产中植物精油的有效种类及具体剂量。
单宁分为水解型单宁(hydrolysable tannin,HT)和缩合型单宁(condensed tannin,CT)。其中,HT是含羟基的碳水化合物,与蛋白质结合能力强,能与微生物细胞膜上的蛋白质结合形成复合物沉淀,抑制产甲烷菌生长;CT是高分子黄酮类化合物,主要通过减少纤维消化减少H的生成,从而减少CH4生成。在反刍动物饲粮中适量添加单宁对生产性能及CH4减排有益。Ngámbi等[6]研究发现,在公羊饲粮中添加HT能降低CH4产量,并提高羊肉抗氧化活性;Torres等[28]利用Meta分析证明单宁的添加能降低绵羊CH4产量,并提高肉中脂肪酸含量;此外,用富含单宁的印揀叶饲喂山羊能提高氮利用率和生长性能[29];但金合欢提取物的添加会对营养物质的消化率产生负面影响[7],单宁的涩味和苦味会对适口性会产生影响[30-31]。因此,使用单宁饲喂反刍动物时应注意其添加剂量和比例,不同来源的含单宁饲料应首先确定其单宁组成和含量,含量过高可能对微生物产生毒性,使饲粮的消化率和降解率降低,而对适口性的影响可采取包被技术起到中和收敛作用[32]。
皂苷是一类天然表面活性糖苷,能与原虫表面甾醇结合使其裂解死亡,同时能够抑制与原虫相关的产甲烷菌活性,广泛存在于苜蓿、丝兰的提取物中。茶皂素是皂苷的一种,饲粮中添加10和20 g/kg DM的茶皂素能显著降低绵羊的CH4排放量,但添加5 g/kg DM茶皂素则无显著作用[33];在泌乳奶牛上添加0.52%DM的茶皂素,体外呈现减排效果,但在体内并无作用[34];体外添加苜蓿皂苷能有效抑制CH4生成[35],且CH4产量和原虫数随剂量增加(0~0.5 g/L)线性减少,最高可达29%和50%[34]。由此可见,皂苷在适当范围内具有CH4减排的能力,但体内和体外对CH4的抑制效果并不一致,需要更多的体内试验来验证体外试验结果,保障在生产上达到有效的添加剂量。
2.1.2 其他添加剂
大型海藻中含有溴仿和多种活性物质,溴仿能减少CH4排放并与维生素B12反应抑制产甲烷菌酶的活性,活性物质中包含皂苷、单宁和抗菌肽等能抑制产甲烷菌的生长。研究表明,海藻的添加与CH4生成呈负相关[36]。红藻提取物能降低CH4排放量并减少产甲烷菌数量[37],红海藻天门冬具有缓解反刍动物CH4排放的潜力[38],Roque等[39]用天门冬饲喂奶牛使产甲烷菌的代谢功能受限,CH4产量降低;Choi等[40]发现,马尾藻也能够显著降低总产气量以及CH4和CO2产量。海藻的作用效果显著且具有多种生物活性物质,但其生产成本较高,不易贮存和运输,若想广泛使用还需开发更好的贮存运输办法,降低成本并确定具体的添加量。
益生菌(probiotics)是对宿主有益的活性微生物,常见有乳杆菌、芽孢杆菌和酵母等。大部分益生菌通过促进瘤胃发酵、减少产甲烷菌数量以及影响瘤胃微生物活性来减少CH4生成[4];酵母菌通过刺激产乙酸菌与产甲烷菌竞争H2,减少CH4生成[41]。在羔羊饲粮中添加粉末或液体形式的益生菌均能显著降低CH4产量[42];在肉牛饲粮中添加乳酸菌也能够有效降低CH4产量并提高总产气量[43]。
益生元(prebiotics)包括蛋白质、多肽、脂肪、低聚糖和多糖等不易消化的成分,能与病原菌结合抑制其生长,刺激有益菌增殖,对瘤胃发酵具有积极作用。Kido等[44]在犊牛断奶前后添加β-葡聚糖益生元,瘤胃中纤维分解菌比例增加,产甲烷菌比例降低。益生元主要通过改变菌群结构影响产甲烷菌的细胞壁通透性,以此来减少CH4生成,通常与益生菌合用发挥协同作用。
2.2 抑制CH4合成关键酶活性
3-硝基氧丙醇(3-nitroxypropanol,3-NOP)是反刍动物CH4减排的有效添加剂,能够抑制瘤胃中CH4生成且对生长性能无不良影响[45]。Hristov等[8]在奶牛上发现,3-NOP能抑制CH4产生;Van Gastelen等[46]在泌乳奶牛上也得到相同结论;Vyas等[47]在育肥肉牛饲粮中添加3-NOP使CH4产量显著降低,且饲料转化效率有所提高,当3-NOP添加量在0~280 mg/kg DMI时,CH4的减少与3-NOP剂量呈线性相关[48]。其机制主要是3-NOP的结构与甲基辅酶M相似,能够结合甲基辅酶M还原酶的活性位点,使甲基辅酶M还原酶失活,阻止其催化产甲烷菌产生CH4,从而降低CH4产量[49]。
2.3 与产甲烷菌竞争H2
2.3.1 有机酸
苹果酸、延胡索酸等均为有机酸,在饲粮中可作为丙酸前体抑制CH4生成,也能与产甲烷菌竞争H2。Li等[50]用延胡索酸饲喂山羊发现,其具有持续抑制CH4产量的作用,且能降低乙酸/丙酸比值。Bharathidhasan[51]的体外试验发现,添加超过0.26%的苹果酸能有效降低CH4产量。不同有机酸拥有不同的还原途径,对CH4的抑制效果不同,其中延胡索酸的效果最佳[52]。
2.3.2 脂质
在反刍动物饲粮中添加油脂能够抑制瘤胃CH4产生,抑制效果取决于脂质来源、数量、饱和度以及脂肪酸碳链长短等。花生油、菜籽油和大豆油在体外试验中均能降低CH4产量[53];棕榈油能使绵羊的CH4产量降低[54];玉米油能降低山羊瘤胃中的溶解氢浓度及CH4产量[55]。不饱和脂肪酸(unsaturated fatty acid,UFA)在瘤胃中的氢化作用下消耗H2,与产甲烷菌形成竞争,从而减少CH4排放。Yang等[56]在奶牛高脂饲粮中添加UFA发现,随UFA添加比例的升高,CH4产量逐渐降低且纤维杆菌丰度显著降低,螺旋体科细菌丰度显著提高。椰子油属于中链脂肪酸,能通过改变微生物结构和瘤胃发酵模式降低CH4产量[57]。脂质易得且不易对动物造成危害,但其高能量会导致采食量下降,且过量添加油脂还可能对瘤胃发酵参数和瘤胃菌群产生不利影响,因此其添加量还需针对不同反刍动物的不同生长阶段进行调整和验证。
2.3.3 硫酸盐和硝酸盐
硫酸盐和硝酸盐属于化学合成抑制剂,从热力学上看(表1),硫酸盐和硝酸盐的还原作用强于瘤胃内CH4生成途径,可与产甲烷菌竞争H2从而减少瘤胃CH4排放。

表1 利用不同电子受体进行的还原反应的吉布斯自由能Table 1 Gibbs free energy of reduction reaction with different electron acceptors
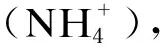
在饲粮中添加硫酸盐后,当瘤胃内硫酸盐含量高时,硫酸盐还原菌(sulfate reducing bacteria,SRB)能与产甲烷菌竞争H2并生成硫氢根离子(HS-),由此减少CH4生成。Wu等[61]发现,在饲粮中添加硫能够提高SRB丰度并减少CH4排放;干酒糟及其可溶物(distillers dried grains with solubles,DDGS)和硫酸钙能够增加奶牛产奶量并显著降低CH4产量[62];山羊饲粮中添加硫酸钠和SRB也能使CH4产量降低[63];在肉牛上,CH4产量随硫酸钠的增加而降低,且瘤胃上皮屏障的通透性增加[64]。因此,在不影响其生产性能的基础上添加硫酸盐能够有效降低反刍动物的CH4产量。但由于其终产物可能产生的硫化氢(H2S)具有毒性,动物吸入后会产生脑脊髓灰质软化症(PEM),且目前为止对于硫酸盐能够完成CH4减排的研究多为体外试验,需要更多的体内试验进一步确定硫酸盐最适添加剂量,避免对动物造成不可逆的损伤;除此之外,硫酸盐在瘤胃中通过何种路径完成CH4减排还尚不明确,还需更多试验验证。
3 甲烷厌氧氧化
甲烷厌氧氧化是湖泊、沼泽、海洋以及湿地等生态系统中CH4减排的重要途径,其CH4减排能力最高可达90%,是一类极具潜力的CH4减排途径。根据电子受体类型不同,甲烷厌氧氧化可分为硫酸盐依赖型甲烷厌氧氧化(sulfate-dependent anaerobic methane oxidation,S-DAMO)、硝酸盐/亚硝酸盐依赖型甲烷厌氧氧化(nitrate/nitrite dependent anaerobic methane oxidation,N-DAMO)和金属依赖型甲烷厌氧氧化(metal-dependent anaerobic methane oxidation,M-DAMO)。甲烷厌氧氧化在严格厌氧条件下完成,主要由甲烷厌氧氧化古菌(anaerobic methanotrophic archaea,ANME)发挥作用,若能证明在瘤胃中存在,将为反刍动物CH4减排工作提供新思路。表2列举了不同终端电子受体的甲烷厌氧氧化反应及所提供的吉布斯自由能。

表2 不同终端电子受体的甲烷厌氧氧化反应及所提供的吉布斯自由能Table 2 Anaerobic methane oxidation reactions with different terminal electron acceptors and Gibbs free energy provided by them
3.1 S-DAMO


CH4:甲烷 methane;CO2:二氧化碳 carbon dioxide;ANME:甲烷厌氧氧化古菌 anaerobic methanotrophic archaea;SRB:硫酸盐还原菌 sulfate reducing bacteria;Methyl-S-CoM:甲基-S-辅酶M methyl-S-coenzyme M;Methyl-H4MPT:甲基-四氢甲基蝶呤 methyl-tetrahydromethylpterin;Methylene-H4MPT:亚甲基-四氢甲基蝶呤 methylene-tetrahydromethylpterin;Methenyl-H4MPT:次甲基-四氢甲基蝶呤 methyl-tetrahydromethylpterin;Formyl-H4MPT:甲酰-四氢甲基蝶呤 formyl-tetrahydromethylpterin;Formyl-MFR:甲酰-甲烷呋喃 formyl-methanofuran;Mcr:甲基辅酶M还原酶 methyl-coenzyme M reductase;Mtr:甲基辅酶M转移酶 methyl-coenzyme M transferase;Mer:亚甲基-四氢甲基蝶呤还原酶 methylene-tetrahydromethylpterin reductase;Mtd:F420型亚甲基-四氢甲基蝶呤还原酶 F420 type methylene-tetrahydromethylpterin reductase;Mch:亚甲基-四氢甲基蝶呤环化酶 methylene-tetrahydromethylpterin cyclase;Ftr:四氢甲基蝶呤甲酰转移酶 tetrahydromethylpterin formyltransferase;Fmd:甲酰-甲烷呋喃脱氢酶 formyl-methanofuran dehydrogenase。图5同 the same as Fig.5。图4 硫酸盐甲烷厌氧氧化的3种假说途径Fig.4 Three hypothetical approaches to anaerobic oxidation of sulfate methane[74]
3.2 N-DAMO


Hdr:杂二硫化物还原酶 heterodisulfide reductase;H2:氢气 hydrogen;N2:氮气 nitrogen。图5 N-DAMO反应过程Fig.5 N-DAMO reaction process[77]
3.3 M-DAMO
M-DAMO主要以三价铁离子(Fe3+)、四价锰离子(Mn4+)等金属离子作为电子受体,将其还原为二价铁离子(Fe2+)和二价锰离子(Mn2+),在河道沉积物、海洋沉积物以及沿海沉积物中普遍存在[2,65-66]。M-DAMO其主要功能微生物是海洋底栖生物群-D(marine benthic group-D,MBG-D),ANME-2及其亚类也均参与该反应[78-79]。
3.4 瘤胃中的甲烷厌氧氧化
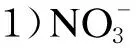
4 小 结
CH4产生是反刍动物瘤胃内不可规避的过程,其对环境的危害不可忽视,在饲粮中添加不同类型的添加剂的确能够降低反刍动物的CH4排放水平,但存在减排效果瞬时性、作用效果不稳定以及添加剂间联用效果及适宜添加量尚不明确等问题。而甲烷厌氧氧化作为自然界中重要甲烷汇,其CH4减排能力高达90%。瘤胃也属于严格厌氧环境,温度在39 ℃左右,pH 6.0~7.0符合甲烷厌氧氧化发生条件,且在饲粮中添加硫酸盐和硝酸盐使CH4排放量降低且SRB、NC10门菌以及古菌总数上升,这些能都证明甲烷厌氧氧化过程有在瘤胃中存在的可能,但现阶段并没有直接数据证明其存在,若能使用如同位素示踪等技术手段直接能证明甲烷厌氧氧化在瘤胃中存在,将为反刍动物甲烷减排工作提供新思路,对提高饲料利用效率以及减少CH4排放、缓解温室效应具有重要意义。
