携带松材线虫对松墨天牛成虫肠道和气管细菌的影响
2023-11-30顾宇彤朱浩诚樊建庭赵莉蔺
顾宇彤, 朱浩诚, 常 悦, 周 娇, 樊建庭, 赵莉蔺,3,4,*
(1. 浙江农林大学林业与生物技术学院, 生物农药高效制备技术国家地方联合工程实验室, 杭州 311300;2. 中国科学院动物研究所, 农业虫害鼠害综合治理研究国家重点实验室, 北京 100101;3. 中国科学院大学, 北京 100049; 4. 中国科学院生物互作卓越创新中心, 北京 100049)
在多物种互作以及复杂的营养级层面,宿主携带的微生物为其适应性进化提供了便利的条件。大部分的线虫与植物和动物的寄生、共生的紧密联系是通过一些特定的细菌建立起来的(Johnson and Rasmann, 2015)。昆虫病原线虫(entomopathogenic nematode, EPN)是昆虫的专性寄生线虫,可以通过自身携带的细菌完成寄主的侵染循环,最终可能导致寄主死亡。而非寄生关系中,微生物并不会致死寄主,微生物与昆虫互作的过程对其生活史的维持不可或缺(Poinar, 1993)。如腐生型的线虫Pristionchus和Diplogasteroides以滞育型虫态存在于活的金龟子当中。当金龟子死后,线虫则转变为繁殖型虫态,以寄主死后产生的各种细菌、真菌和其他线虫为食,并进行繁殖扩散(Herrmannetal., 2006; Raeetal., 2008)。
松材线虫Bursaphelenchusxylophilus是松材线虫病的病原体,是世界上对针叶树最严重的威胁之一(Vicenteetal., 2013a)。迄今已成为我国最为严重的林业外来有害生物之一,对我国的林业造成了严重的经济损失(Zhaoetal., 2014)。尽管采取了相应的预防和控制措施,我国的松材线虫病的疫区数量仍然在逐年增加。作为松材线虫病扩散的主要媒介松墨天牛Monochamusalternatus,在我国是危害针叶树的重要蛀干害虫,与松材线虫之间存在特殊的“气管共生”关系,松材线虫扩散型4龄幼虫在松墨天牛蛹期进入天牛气管,有时候在1头天牛的气管系统里会有二三十万头线虫(Zhaoetal., 2014, 2016)。松材线虫病的发生与扩散是多物种互作的结果,尽管寄主松树-松材线虫-媒介天牛复合体中的相关的微生物(细菌和真菌)并不能直接导致松材线虫病的发生,但是微生物在松材线虫病害体系中的作用不可忽视。媒介天牛蛹室和气管中的优势真菌——蓝变菌对松材线虫-媒介天牛入侵复合体的繁殖表型具有促进作用(Zhaoetal., 2014)。目前大多研究集中在松材线虫携带细菌对松树的致病及抗性影响(Vicenteetal., 2012, 2013b),并且细菌群落在干扰和调控松墨天牛免疫能力和携带松材线虫方面具有很大的生防潜力(Zhouetal., 2018),然而,对媒介携带的细菌种类及功能研究较少,尤其是对昆虫气管微生物的研究有待进一步探索。
本研究通过对松墨天牛成虫气管和肠道微生物多样性进行研究,探究在携带松材线虫的情况下,松墨天牛成虫气管和肠道微生物的变化,为进一步利用微生物防控松材线虫病提供可靠的理论依据。
1 材料与方法
1.1 松墨天牛成虫肠道和气管分离
为了消除不同地理样本之间由于温度和湿度海拔等非生物因素的影响,本研究松墨天牛成虫收集自贵州省遵义市播州区马尾松林(27°13′N, 106°17′E),在松林中利用诱捕器人工收集,松墨天牛的采集从5月持续至8月。解剖前先对天牛体表进行杀菌处理,先用75%乙醇浸泡30 s,再用无菌水冲洗消除残留乙醇,用昆虫针将其固定在蜡盘上,用解剖剪从腹部纵向剪开到头部,后用镊子取出肠道,置于1.5 mL离心管中,液氮速冻15 min,-80 ℃冰箱保存;收集肠道后将剩余组织浸泡于PBS中,待气管和体腔中的松材线虫溢出后,取出气管进一步冲洗至松材线虫完全去除后,将气管置于1.5 mL离心管中,液氮速冻15 min,收集的肠道和气管作为携带松材线虫的松墨天牛成虫肠道和气管,如果没有松材线虫溢出,则收集到的肠道和气管作为未携带松材线虫的松墨天牛成虫肠道和气管,并统计每头松墨天牛成虫气管和体腔的松材线虫数量。
1.2 松墨天牛成虫肠道和气管细菌16S rDNA基因测序
利用微生物DNA提取试剂盒QIAamp PowerFecal Pro DNA Kit (50)提取携带和未携带松材线虫的松墨天牛成虫肠道和气管细菌总DNA,使用1%的琼脂糖凝胶电泳检测DNA的提取质量,每个样本3个重复,完成基因组DNA抽提后,利用1%琼脂糖凝胶电泳检测抽提的基因组DNA。基于16S rDNA基因序列,设计合成带有barcode的特异引物:799F (5′-AACMGGATTAGATACCCKG-3′);1193R (5′-ACGTCATCCCCACCTTCC-3′),PCR扩增16S rDNA基因序列。扩增PCR体系(25 μL): 2×KAPA HiFi HotStart ReadyMix 12.5 μL, 上下游引物各5 μL, 模板DNA 2.5 μL。PCR反应程序: 95 ℃预变性3 min; 95 ℃变性30 s, 55 ℃退火30 s, 72 ℃延伸30 min, 共25个循环;72 ℃延伸5 min。使用AxyPrepDNA凝胶回收试剂盒(Axygen Biosciences, Union City, CA, 美国)切胶回收PCR产物,Tris-HCl洗脱;2%琼脂糖电泳检测。参照电泳初步定量结果,将PCR产物用QuantiFluorTM-ST蓝色荧光定量系统(Promega, 美国)进行定量检测,之后按照每个样本的测序量要求,进行相应比例的混合。利用Illumina公司的MiseqPE300平台进行测序。
1.3 物种注释与评估
根据不同相似度水平,对携带和未携带松材线虫的松墨天牛成虫肠道和气管细菌16S rDNA基因测序读段序列进行可操作分类单元(operational taxonomic units, OTUs)划分,通常在97%的相似水平下的OTU进行生物信息统计分析。OTU聚类步骤如下:对优化序列提取非重复序列,便于降低分析中间过程冗余计算量(http:∥drive5.com/usearch/manual/dereplication.html)去除没有重复的单序列(http:∥drive5.com/usearch/manual/singletons.html);按照97%相似性对非重复序列(不含单序列)进行OTU聚类,在聚类过程中去除嵌合体,得到OTU的代表序列。将所有优化序列map至OTU代表序列,选出与OTU代表序列相似性在97%以上的序列,生成OTU表格。为了得到每个OTU对应的物种分类信息,采用RDP classifier贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析,并分别在各个分类学水平(域、界、门、纲、目、科、属和种)统计各样本的群落物种组成。
1.4 数据分析
利用上海美吉生物医药科技有限公司的微生物多样性云分析平台对携带和未携带松材线虫的松墨天牛成虫肠道和气管细菌进行物种组成分析:基于物种注释结果,使用群落柱形图分析和韦恩图分析等分析样本中物种的组成,以探究物种与样本之间的关系,分析样本中的优势菌群/关键菌群等;物种比较分析:基于物种注释结果,通过单样本的多样性Alpha多样性分析反映微生物群落的丰度和多样性,包括Sobs指数、Chao指数(反映群落丰度)、Shannon指数、Simpson指数(反映群落多样性);Beta多样性分析可以通过对不同生境或微生物群落间的物种多样性进行组间比较分析,探索不同分组样本间群落组成的相似性或差异性,常用的方法有非度量多维排列(non-metric multidimensional scaling, NMDS)分析,将对象间的相似性或相异性数据看成点间距离的单调函数,在保持原始数据次序关系的基础上,用新的相同次序的数据列替换原始数据进行度量型多维尺度分析;物种差异分析:基于物种组成及比较分析,进一步使用多组比较/两组比较分析,筛选不同组别间具有显著差异的物种。
2 结果
2.1 松墨天牛成虫肠道和气管细菌物种注释与评估
从携带和未携带松材线虫的松墨天牛成虫42个肠道和气管样本中获得的16S rDNA基因原始序列读数,经过质量过滤和去除嵌合体后,得到2 108 404条高质量序列,每条序列平均长432 bp(表1)。按最小样本序列数抽平,对OTU进行物种分类学注释,共注释到15门26纲66目110科201属296种,OTU数目为444。各组的稀释曲线(Shannon指数)随着测序量增加曲线平缓,说明测序量足够反映微生物多样性;当测序数据量达到一定值时,所有曲线变得平缓,OTU数量不会随着测序量增加而明显增加,表明样品序列数充分(图1)。

图1 松墨天牛成虫肠道和气管的细菌组成稀释曲线Fig. 1 Rarefaction curves of bacterial composition in the gut and trachea of Monochamus alternatus adults

表1 松墨天牛成虫肠道和气管细菌16S rDNA基因测序数据Table 1 Sequencing data of bacterial 16S rDNA gene in the gut and trachea of Monochamus alternatus adults
2.2 松墨天牛成虫肠道和气管的细菌群落结构
由图2可知,携带松材线虫的松墨天牛成虫肠道和气管OTU数量分别为432和413个,分别高于未携带松材线虫的松墨天牛成虫的肠道和气管的OTU数量(分别为410和75个)。4种样本共有的OTU数量为52个,其中携带松材线虫的松墨天牛成虫肠道和未携带松材线虫的松墨天牛成虫的肠道所共有的OTU数量为351个,相似度较高;携带松材松材线虫的松墨天牛成虫气管和未携带松材线虫的松墨天牛成虫气管所共有的OTU数量为60个,相似度较低。
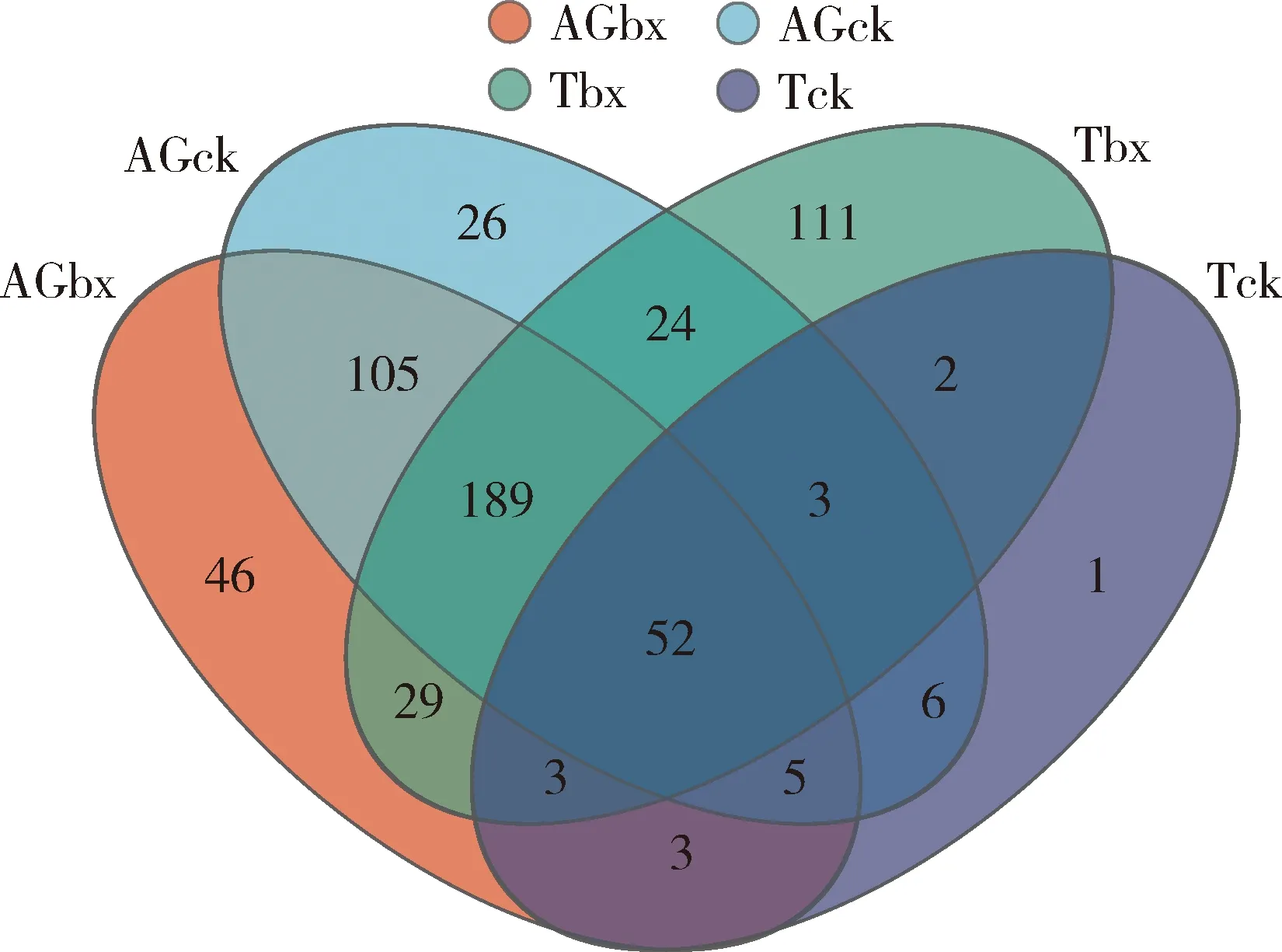
图2 松墨天牛成虫肠道和气管细菌可操作分类单元(OTUs)的韦恩图Fig. 2 Venn diagram of operational taxonomic units (OTUs) of bacteria in the gut and trachea of Monochamus alternatus adults
门水平上(图3: A),携带松材线虫的松墨天牛成虫肠道的优势菌门为变形菌门(Proteobacteria),占74.7%,次优势菌门为蓝细菌门(Cyanobacteria),占13.0%;未携带松材线虫的松墨天牛成虫肠道的优势菌门为变形菌门,占58.1%,次优势菌门为厚壁菌门(Firmicutes),占21.1%。携带松材线虫的松墨天牛成虫气管的优势菌门为变形菌门,占50.7%,次优势菌门为蓝细菌门,占40.4%;未携带松材线虫的松墨天牛成虫气管的优势菌门为厚壁菌门,占54.0%,次优势菌门为变形菌门,占45.9%。

图3 松墨天牛成虫肠道和气管细菌优势门(A)、目(B)和属(C)相对丰度Fig. 3 Relative abundance of bacteria at the phylum (A), order (B) and genus (C) levels in the gut and trachea of Monochamus alternatus adults
从目水平上(图3: B),携带松材线虫的松墨天牛成虫肠道的优势菌目为肠杆菌目(Enterobacterales),占58.4%,次优势菌目为Chloroplast,占13.0%;未携带松材线虫的松墨天牛成虫肠道的优势菌目为肠杆菌目,占44.9%,次优势菌目为乳杆菌目(Lactobacillales),占15.7%。携带松材线虫的松墨天牛成虫气管的优势菌目为肠杆菌目,占41.9%,次优势菌目为Chloroplast,占40.4%;未携带松材线虫的松墨天牛成虫气管的优势菌目为乳杆菌目,占53.9%,次优势菌目为肠杆菌目,占32.4%。
在属水平上(3: C),丰度排序前10名的各优势属在携带松材线虫的松墨天牛成虫肠道、未携带松材线虫的松墨天牛成虫的肠道、携带松材线虫的松墨天牛成虫气管和未携带松材线虫的松墨天牛成虫的气管中所占比例分别为肠杆菌科未分类属(分别占40.8%, 33.1%, 7.2%和25.4%),乳球菌属Lactococcus(分别占0.1%, 14.3%, 0.5%和49.4%),沙雷氏菌属Serratia(分别占10.3%, 2.4%, 32.0%和5.0%),Chloroplast目未分类属(分别占13.1%, 13.4%, 40.4%和0),假单胞菌属Pseudomonas(分别占0.5%, 0.9%, 0.8%和0.4%),布鲁氏菌属Brucella(分别占7.1%, 2.4%, 0.2%和10.7%),大肠埃氏菌属-志贺氏菌属Escherichia-Shigella(分别占2.7%, 8.2%, 0.1%和0.2%),不动杆菌属Acinetobacter(分别占1.5%, 3.2%, 3.7%和0.3%), 肠球菌属Enterococcus(分别占0.0%, 1.2%, 0.0%和4.6%),沃尔巴克氏菌属Wolbachia(分别占0, 0.2%, 0和0.6%)。松墨天牛携带或未携带松材线虫的肠道主要优势菌相似,但其丰度有较大差异;携带或未携带松材线虫的松墨天牛成虫气管在主要优势菌和丰度上都有较大差异。
2.3 松墨天牛成虫肠道和气管的细菌群落多样性
属水平Beta多样性分析结果表明(图4),携带松材线虫的松墨天牛成虫肠道细菌群落结构与未携带松材线虫的松墨天牛成虫肠道细菌群落结构相近,但携带松材线虫的松墨天牛成虫气管细菌群落结构与未携带松材线虫的松墨天牛成虫气管细菌群落结构明显区分开来,说明携带松材线虫对松墨天牛成虫气管细菌群落结构有显著影响。

图4 利用非度量多维排列(NMDS)对松墨天牛成虫肠道和气管细菌进行属水平Beta多样性分析Fig. 4 Beta diversity of bacteria in the gut and trachea of Monochamus alternatus adults at the genus level analyzed by non-metric multidimensional scaling (NMDS)
属水平Alpha多样性分析结果表明(图5),Shannon指数和Simpson指数在不同组之间没有明显差异 (图5: A, B), 松墨天牛成虫肠道细菌多样性高于气管细菌多样性,携带松材线虫的松墨天牛成虫气管细菌多样性高于未携带松材线虫的松墨天牛成虫气管细菌多样性。Sobs和Chao指数在不同组之间有显著差异(图5: C, D),且携带松材线虫的松墨天牛成虫肠道和气管细菌Sobs和Chao指数均高于未携带松材线虫的松墨天牛成虫肠道和气管的,表明携带松材线虫后,松墨天牛成虫肠道和气管的细菌群落丰度升高。
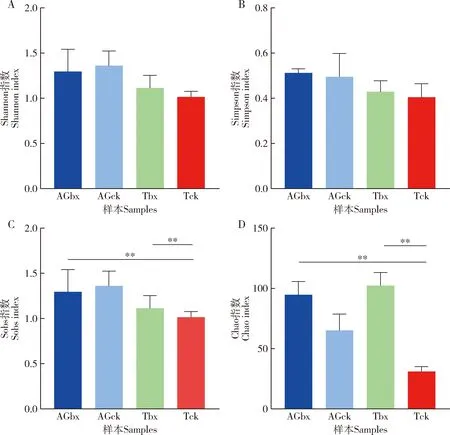
图5 松墨天牛成虫肠道和气管细菌Alpha多样性Fig. 5 Alpha diversity of bacteria in the gut and trachea of Monochamus alternatus adultsA: Shannon指数Shannon index; B: Simpson指数Simpson index; C: Sobs指数Sobs index; D: Chao指数Chao index. 图中数据为平均值±标准误;柱上双星号表示两组间差异显著(P<0.01, T检验)。Data in the figure are mean±SE. The double asterisk above bars indicates significant difference between two groups (P<0.01, T-test).
2.4 松墨天牛成虫肠道和气管细菌菌群丰度的差异
对携带和未携带松材线虫的松墨天牛成虫肠道、携带和未携带松材线虫的松墨天牛成虫气管中细菌进行优势属(图6)的组间差异显著性检验分析,结果表明,在属水平,沙雷氏菌属Serratia、布鲁氏菌属Brucella、经黏液真杆菌属Blautia和肠杆菌属Enterobacter在携带松材线虫的松墨天牛成虫肠道中富集;沙雷氏菌属Serratia和肠杆菌属Enterobacter在携带松材线虫的松墨天牛成虫气管中富集。乳球菌属Lactococcus、痤疮丙酸杆菌属Cutibacterium、鞘氨醇单孢菌属Sphingomonas在未携带松材线虫松墨天牛成虫肠道中富集;乳球菌属Lactococcus、布鲁氏菌属Brucella、肠球菌属Enterococcus在未携带松材线虫的松墨天牛成虫气管中富集。

图6 松墨天牛成虫肠道和气管细菌菌群丰度的差异分析(排名前10的属)Fig. 6 Differential abundance analysis of the bacterial flora in the gut and trachea of Monochamus alternatus adults among the top 10 genera图中P<0.05表示组间差异显著(Kruskal Wallis秩和检验)。The P-value lower than 0.05 in the figure indicates significant difference between groups (Kruskal Wallis H test)
3 讨论
本研究通过对携带松材线虫前后的松墨天牛成虫肠道和气管的细菌多样性测序结果进行分析,探讨松墨天牛肠道和气管的细菌群落组成和结构差异以及携带松材线虫后气管肠道菌群的改变与松材线虫传播的关系。携带松材线虫的松墨天牛成虫肠道细菌群落组成与未携带松材线虫的樟子松墨天牛M.galloprovincialis的相似,肠道中变形菌门、厚壁菌门和蓝细菌门等为主要菌门,肠杆菌目、乳杆菌目等为主要菌目(图3)。变形菌门细菌在昆虫中比较常见(Kirketal., 2016)。Alves等(2016)研究发现樟子松墨天牛和松墨天牛细菌群落以变形菌门为主。上述细菌群可能在天牛的生命周期的不同阶段中发挥作用,影响松材线虫的传播。在松墨天牛成虫气管中,这3种菌门也是主要菌门,其细菌群落组成与肠道的相似,但变形菌门和肠杆菌目为携带松材线虫的松墨天牛成虫气管中主要菌门和菌目,而厚壁菌门和乳杆菌目为未携带松材线虫的松墨天牛成虫气管中的主要菌门和菌目(图3)。目前对于昆虫气管微生物的研究则较少,松墨天牛气管的微生物部分来源于自身的肠道,部分来源于蛹室。松材线虫的微生物群也遗传自松墨天牛(Alvesetal., 2018)。我们研究组发现, 松材线虫扩散型4龄幼虫携带的真菌细菌微生物的丰度和多样性均很低,几乎不会携带蛹室细菌进入松墨天牛气管,并因此实现对松墨天牛的保护,从而逃避松墨天牛免疫,提高松材线虫-松墨天牛复合体的稳定性(Zhangetal., 2021)。所以我们推测,松墨天牛气管细菌可能与其肠道相关。Alpha多样性和Beta多样性分析表明,在携带松材线虫后,松墨天牛成虫气管细菌多样性和丰度升高,其群落结构与未携带松材线虫的松墨天牛成虫气管中的也明显区分开来,推测松墨天牛成虫气管细菌丰度的增加与松材线虫-媒介天牛复合体有着比较紧密的联系(图4, 5)。
本研究进一步对松墨天牛成虫肠道和气管细菌之间的差异进行分析,发现在携带松材线虫后,松墨天牛成虫肠道和气管都出现了一些独有的共生细菌,沙雷氏菌属和肠杆菌属在携带松材线虫的松墨天牛成虫肠道和气管中均富集(图6)。这一结论与其他研究相似,如肠杆菌科在欧洲媒介樟子松墨天牛气管中丰度也较高(Alvesetal., 2016),沙雷氏菌属和芽孢杆菌属Bacillus等在葡萄牙的樟子松墨天牛胸部和腹部丰度较高(Vicenteetal., 2013b)。沙雷氏菌属在松墨天牛属的几个类群,包括松墨天牛、卡墨天牛M.carolinensis以及云杉花墨天牛M.saltuarius幼虫和成虫的肠道中也都可以被检测到。结合以上研究,说明沙雷氏菌属和肠杆菌属在不同地区及不同种的松墨天牛中普遍存在。沙雷氏菌属的细菌是松材线虫病害体系中的优势物种且分布广泛,在松材线虫及松树间的功能呈现多样性(Nascimentoetal., 2015; Proençaetal., 2017)。如SerratiaplymuthicaM24T3具有杀线虫活性并提高寄主压力耐受(Proençaetal., 2019)。S.marcescensstrain PWN146通过粘附在松材线虫体表提高对松树的毒力(Vicenteetal., 2016),而S.quinivoransBXF1对松材线虫无致死效应,可被携带并促进寄主松树根的生长(Nascimentoetal., 2016),S.marcescensAHPC29对线虫及其媒介都具有一定的毒性(田浩楷, 2022)。也有研究表明,S.marcescens是否致病与宿主的肠道微生物的稳态相关(Raymannetal., 2017; Heuetal., 2021)。然而沙雷氏菌在松材线虫与松墨天牛间的功能报道较少。有研究发现,沙雷氏菌属的部分种类有助于松材线虫在高活性氧压力的寄主松树中存活(Vicenteetal., 2013a),在宿主血淋巴中的粘质沙雷氏菌会逃避宿主免疫并致死宿主。在携带松材线虫后,松墨天牛气管中沙雷氏菌属增加,可能与松材线虫携带的沙雷氏菌属有关。之前的研究表明,沙雷氏菌属量大时可以使得松墨天牛致死,如果加大沙雷氏菌属的量,将成为杀灭松墨天牛的生防因子。总之,基于沙雷氏菌属在松墨天牛肠道和气管中有较强的定殖能力,以及对于线虫及其媒介均有致死效果,沙雷氏菌属可以作为生物防治松材线虫病害的非常有潜力的候选靶标。同时,其在松墨天牛气管中的富集及功能还需进一步研究。
松材线虫病在我国快速传播蔓延,且逐年加剧,迄今成为我国最为严重的林业外来有害生物之一,对其进行防控刻不容缓。综上所述,松材线虫的携带对松墨天牛成虫气管和肠道细菌都会产生影响,研究松材线虫侵染后松墨天牛成虫肠道和气管菌群组成和群落的变化以及优势种的变化,对探究松材线虫-松墨天牛复合体与细菌间的关系十分重要。本研究发现松材线虫可增加松墨天牛成虫气管细菌丰度,沙雷氏菌属Serratia在松墨天牛成虫气管中的富集与松材线虫-媒介天牛复合体有着比较紧密的联系。目前已有控制病媒传播疾病的策略,利用微生物组的功能,包括使用益生菌作为植物防御促进剂,或通过宿主微生物组的操纵来破坏昆虫媒介的生命周期(Riccietal., 2012; Lebeis, 2014; Laceyetal., 2015)。本研究为松墨天牛成虫肠道和气管微生物多样性研究提供了理论基础,将为松材线虫的防控技术提供重要的理论依据。
