辣椒高多态性KASP标记的开发
2023-11-28常立春郭春贵张立国梁建丽王晓武
张 涛 常立春 郭春贵 张立国 武 剑 梁建丽 高 杰 王晓武*
(1 新疆农业大学园艺学院,新疆特色园艺作物种质资源与高效生产实验室,新疆乌鲁木齐 830052;2 中国农业科学院蔬菜花卉研究所,蔬菜生物育种全国重点试验室,北京 100081;3 赣州农业学校,江西赣州341000)
遗传多样性是生物体之间遗传物质的差异,并且这种差异可以遗传给后代。基因组学的飞速发展实现了在全基因组范围大规模鉴定DNA 序列变异(Batley & Edwards,2009),包括单核苷酸多态性(SNP)、插入缺失(InDel)以及更大的结构变异(SV)。其中,SNP 数量最多,分布最广泛,非常适合用于开发分子标记。分子标记在研究生物的进化途径、分析遗传多样性、开展基因图位克隆时都是必不可少的研究工具,特别是在作物育种中分子标记辅助选择(MAS)发挥了巨大的作用,在多基因聚合、目标基因快速导入、远缘杂交等育种过程中都能显著提高育种效率。
辣椒(Capsicumspp.)是茄科(Solanaceae)植物的一员(2n= 24),世界范围内辣椒品种繁多(Kim et al.,2014;Pereira-Dias et al.,2019)。中国市场的辣椒品种遗传背景较窄,对辣椒的育种改良不利(顾晓振,2016)。对中国市场的辣椒品种进行遗传多样性分析,将为辣椒遗传改良提供指导。前人利用分子标记开展了辣椒的遗传多样性分析。廖芳芳等(2023)利用果实性状与STR 标记检测相结合的方法对30 份辣椒种质资源进行遗传多样性研究,发现地理隔离并未对辣椒的遗传分化产生显著影响;张曼等(2020)基于20 个SSR 标记对112 份朝天椒种质资源进行遗传多样性分析,发现总体上材料的多样性与地理来源存在一定关联,但同时也存在地区间的互相渗透。
竞争性等位基因特异PCR(Kompetitive Allele Specific PCR)简称为KASP 标记,具有标记转化成功率高、分型准确、开发和检测费用低、检测通量灵活性强等特点,是目前开展高通量基因分型的主流标记类型(Semagn et al.,2014;王鹏 等,2021;范妍芹 等,2022)。KASP 标记是开展遗传多样性分析的有力工具。Yang 等(2020)在对4份大白菜重测序的基础上,鉴定筛选出SNP 位点,选择在染色体上遗传位置分布均匀的53 个KASP标记,对34 份大白菜材料进行基因分型和聚类分析,结果显示34 份大白菜可聚为3 簇,与其结球类型相一致;Shen 等(2021)利用23 份代表性青花菜材料全基因组重测序数据,鉴定并筛选了SNP位点,开发出100 个KASP 标记,对372 份青花菜材料进行遗传多样性分析,并从中选择了28 个KASP 标记对青花菜材料DNA 指纹图谱进行构建;Scholten 等(2016)利用转录组数据开发了葱属的洋葱与野葱和大葱间具有多态性的1 100 个KASP标记,用于构建种间的遗传连锁图谱;任海龙等(2023)从前人开发的200个芸薹种KASP 标记中,筛选出18 个核心KASP 标记,并构建了89 份菜薹品种的SNP 指纹图谱,证明了KASP 技术在菜薹品种鉴定中的可行性。最近,Zhang 等(2023)基于GWAS 分析的结果开发了165 个辣椒疫霉病抗性的KASP 标记,经验证,其中在抗/感材料之间有多态性的30 个KASP 标记被认为与辣椒疫霉病抗性相关联。此外,欧立军等(2019)开发了与辣椒素含量紧密连锁的KASP 标记。但是,迄今关于辣椒全基因组范围分布的、在资源材料或品种间具有高多态性、能够进行背景筛选的KASP 标记还未见报道。
基因组中SNP 广泛存在,其中绝大多数没有受到选择压力,使得它们在进化上是中性的,可以更完整地估计多样性水平(Edwards et al.,2007)。本研究旨在开发一套在辣椒基因组中均匀分布、在国内辣椒品种中具有高多态性、普适性强的KASP标记集,为开展辣椒资源多样性评估和育种改良提供技术支撑。
1 材料与方法
1.1 试验材料
供试辣椒材料包含安信沃椒的F2代以及来自全国14 个引种地区的207 份辣椒材料(表1),其中安信沃椒的F2代用于KASP 标记的多态性初步筛选,207 份辣椒材料用于多态性KASP 标记在辣椒品种资源中多态性验证。全部辣椒材料于2021年4 月播种至中国农业科学院蔬菜花卉研究所日光温室。

表1 207 份供试辣椒材料引种名称及来源
1.2 试验方法
1.2.1 DNA 提取 取辣椒幼嫩叶片,采用磁珠法提取DNA(张立国,2022)。使用NanoDrop1000(Thermo,USA)对DNA 进行质检。
1.2.2 KASP 标记设计 一套KASP 引物包括两条正向竞争性引物和一条反向通用引物。利用安信沃椒F1代重测序数据(中国农业科学院蔬菜花卉研究所分子遗传课题组前期已完成)与参考基因组CM334 进行比对,获得变异数据集,再根据辣椒的高密度遗传图谱(Zhang et al.,2019)与物理图谱的对应关系,确定SNP 位点对应的遗传位置,以20 cM 为间距从中筛选出234 个位点用于开发KASP 标记。利用DNAMAN(v6)与Primer 3(https://bioinfo.ut.ee/primer3-0.4.0/) 设计KASP引物。着丝粒区域只设计1 个标记(Zhang et al.,2019)。引物设计标准:引物长度20~26 bp,Tm值58~63℃,自身互补性小于5,3′端互补性小于3(Long et al.,2017;Wu et al.,2020;Yang et al.,2020)。
1.2.3 KASP 标记多态性筛选 利用Caliper(Life Sciences,USA) 和QuantStudio 12K Flex(Life Sciences,USA)进行引物多态性筛选。选取30株安信沃椒F2代植株,同时设置2 个空白对照(ddH2O)。反应体系包括样本DNA 2.5 μL(浓度为30~50 ng · μL-1)、HiGeno 2 × Probe Mix A(北京嘉程生物科技有限公司)2.5 μL、SNP primer Mix(上海生工生物公司)0.07 μL(100 μmol · L-1,引物配比为A1∶A2∶C = 1∶1∶2.5,其中A1、A2表示两条正向竞争性引物,C 表示反向通用引物)。
KASP 标记的PCR 扩增程序:阶段1,95 ℃预变性10 min;阶段2,95 ℃变性20 s,61 ℃退火40 s(每个循环降低0.6 ℃,循环数为10);阶段3,95 ℃变性20 s,55 ℃退火40 s(循环数为34)。
1.2.4 KASP 标记体系转化 为实现更小的反应体系以及适应高通量基因分型,反应体系转化为烘干DNA 体系,文中出现的“体系转化”均指引物筛选体系转化为高通量分型体系。反应体系包括20~30 ng DNA,SNP primer Mix 0.028 μL,HiGeno 2 × Probe Mix A 1 μL。PCR 扩增程序阶段3 循环数变为39,其他条件与1.2.3 相同。
1.2.5 KASP 标记在辣椒品种资源中多态性验证 使用Microsoft Excel 的宏插件GenAlex(6.51 b2)对基因分型原始数据进行处理(Smouse et al.,2017),导出popgen 的输入文件。使用popgen32计算引物在引种材料中的多态性位点数及多态性位点百分率(Brown et al.,1980;Brown & Feldman,1981)。
2 结果与分析
2.1 辣椒KASP 标记开发
在辣椒全基因组范围共设计了234 个KASP 标记。所有设计引物的Tm 值在57.9~62.9 ℃之间,两条竞争性引物的Tm 值差值在0~2.3 ℃范围内。引物的序列长度20~26 bp,两条竞争性引物的序列长度差为0~3 bp。位点的变异类型有12 种,其中碱基互补配对氢键数量变化的有201 个,氢键数目未变化的有33 个。变异位点碱基互补配对氢键数目为三键不变时,每对引物的两条竞争性引物的长度差均为0;变异位点的碱基互补配对氢键数目为二键不变时,两条竞争性引物的长度差为0 的引物数量仅占43%;变异位点的氢键数目变化时,两条竞争性引物的长度差为0 的引物数量占58.7%。
引物的发夹结构分为引物自身互补性和3′末端互补性。在234 个标记中,引物存在发夹结构的有119 个,无发夹结构的有115 个;引物自身互补性为0~5,3′末端互补性为0~3。
2.2 辣椒KASP 标记多态性筛选
234 个KASP 标记中,159 个在F2群体中表现多态性,引物设计成功率为67.9%(图1)。在具有多态性的标记中,包括32 个相同基因型分簇集中的标记(图1-a),这类标记为最优标记;以及67个相同基因型分簇较分散的标记(图1-b),7 个杂合基因型和纯合基因型较近并且相同基因型分簇集中的标记(图1-c)和53 个杂合基因型和纯合基因型较近并且相同基因型分簇分散的标记(图1-d),这3 类标记后续进行种质资源或品种材料的遗传多样性分析时存在风险。不可使用的75 个标记的分型可分为3 类,无扩增的标记占比最大,为46 个(图1-e);4 个为相同基因型分簇分散的标记(图1-f),这类标记虽然可以将样本聚成3 种基因型,但其过于分散使3 种基因型的分型结果彼此之间距离太近,容易产生误判,故将其归为无法使用;25个为纯合基因型与杂合基因型无法区分的标记(图1-g)。
辣椒的12 条染色体中,每条染色体引物设计成功率不同,其中5 号染色体成功率最低,为56.0%;6 号染色体成功率最高,为100.0%(表2)。在115 个引物无发夹结构的KASP 标记中,引物设计的成功率为73%,与整体成功率基本相同,说明发夹结构并不会对多态性标记的引物设计造成大的干扰。
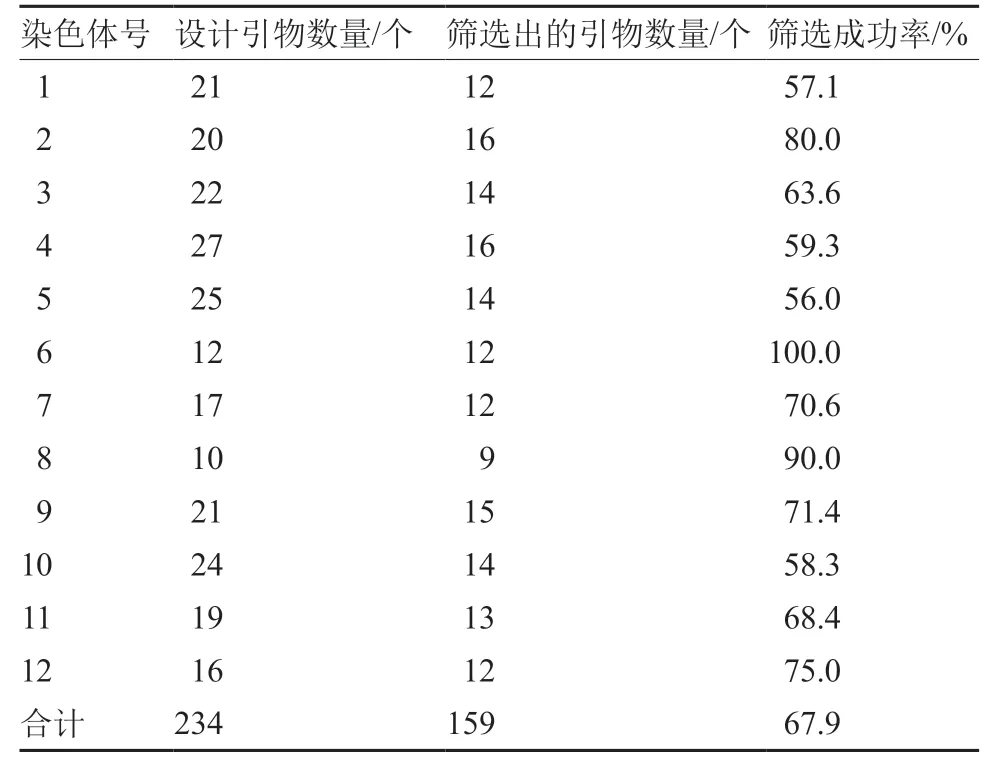
表2 辣椒多态性KASP 标记在12 条染色体上的分布
2.3 KASP 标记高通量反应体系转化
根据分型结果,从159 个可使用的KASP 标记中筛选出141 个标记用于体系转化(表3)。筛选原则为:分簇集中优于分簇分散;分簇分散优于杂合纯合靠近;杂合纯合靠近的引物中,分簇集中优于分簇分散。
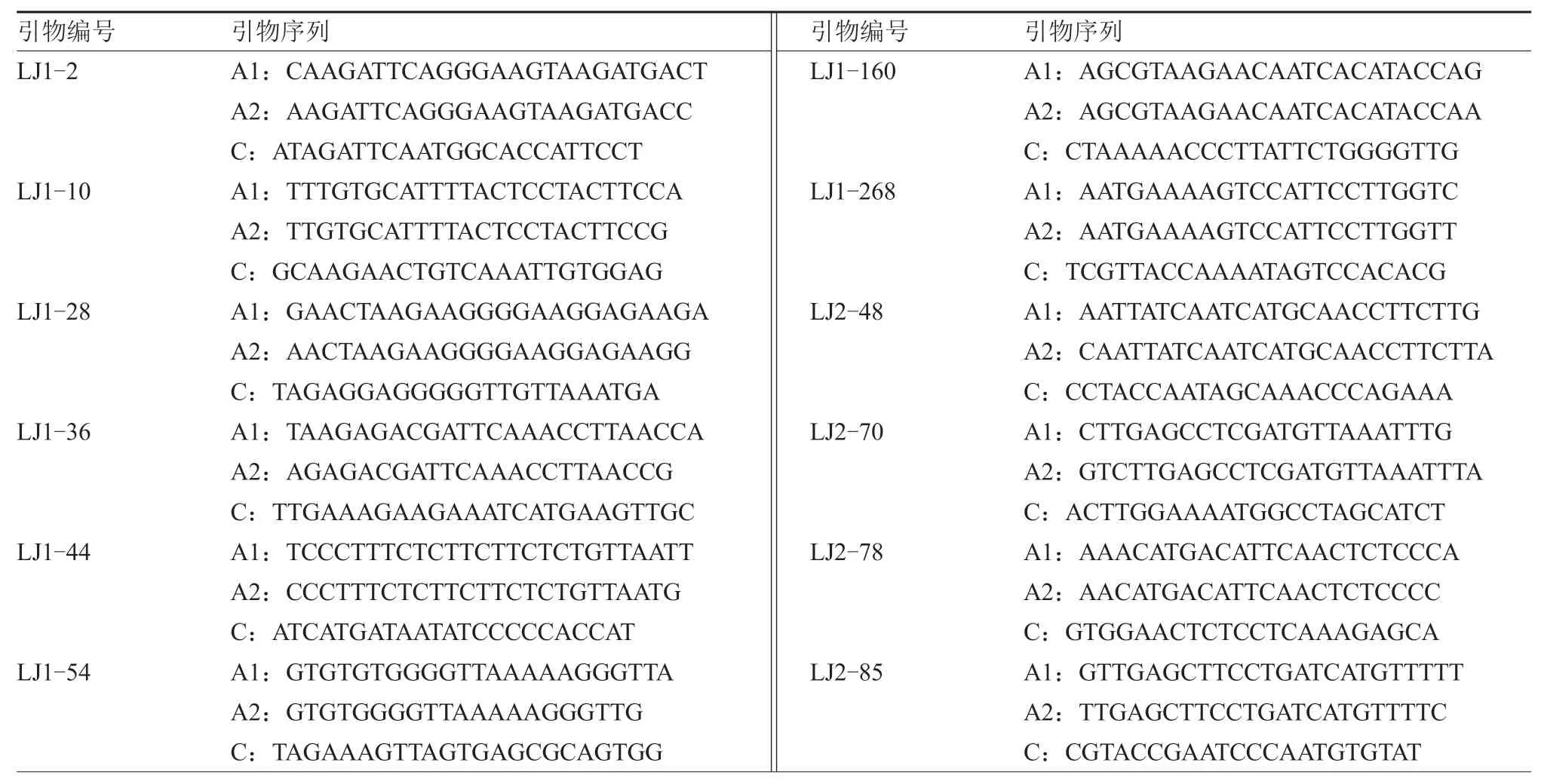
表3 KASP 分子标记集引物序列
141 个辣椒KASP 标记能够成功用于高通量反应体系的有130 个,成功率为92%,这些KASP 标记在辣椒染色体上基本均匀分布(图2)。分型结果中,有42 个标记与进行标记筛选时的分型结果不同,其中31 个标记表现为分簇效果提升,10 个标记表现为效果下降(表4)。
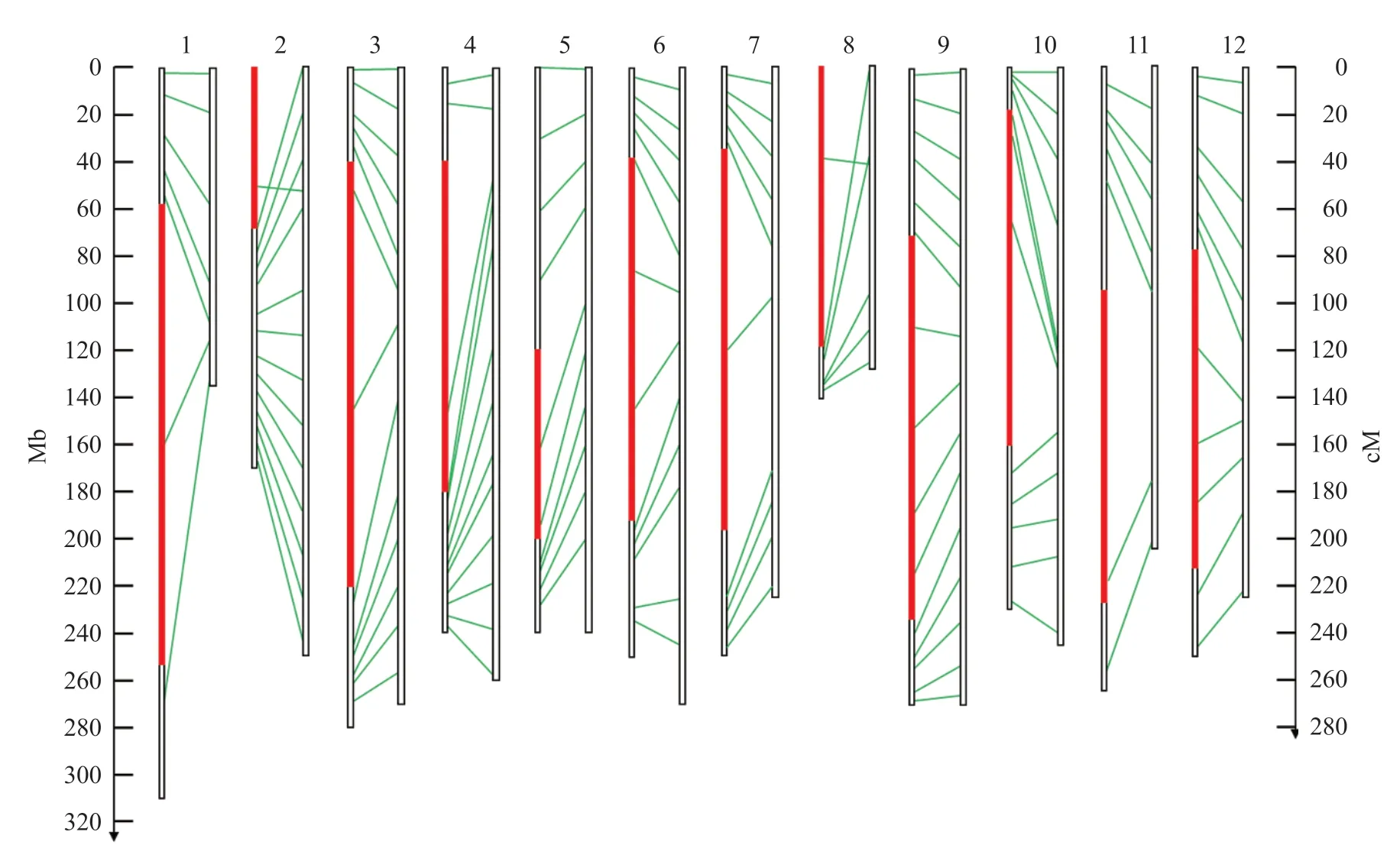
图2 KASP 标记在辣椒染色体上的物理位置与遗传位置的对应关系

表4 体系转化后分型效果与标记筛选时表现不同的KASP标记数量
2.4 多态性KASP 标记的普适性验证
按照引种来源地将207 份辣椒材料分为14 个群体,通过使用popgen 计算所有材料的多态性位点数及多态性位点百分率来验证设计引物的多态性和稳定性。遗传多样性分析结果表明,130 个KASP 标记在辣椒材料中多态性表现良好,多态性位点数为24~129 个,多态性位点百分率范围为18.46%~99.23%(表5)。
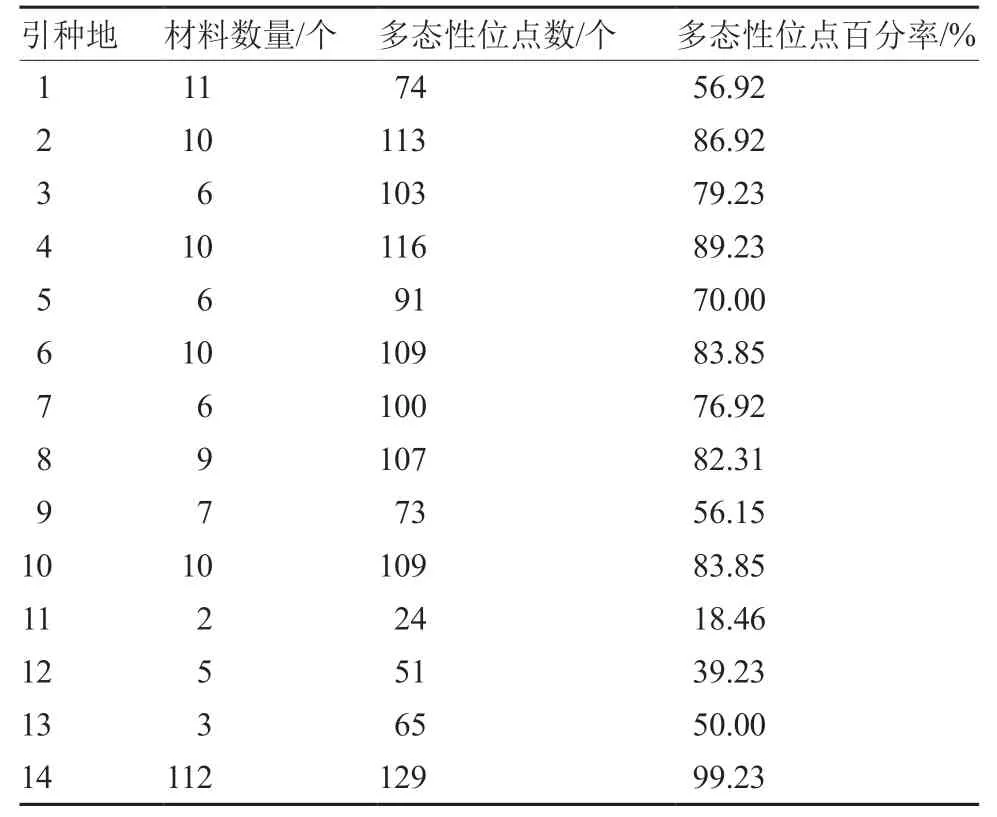
表5 130 个KASP 标记在207 份辣椒材料中的多态性位点数及百分率
3 讨论与结论
影响KASP 引物设计成功的主要因素有4 个:引物自身的Tm 值、序列长度、发夹结构和两条竞争性引物的扩增效率。引物的Tm 值决定了引物的特异性结合的效率,Tm 值越高引物特异性越强,KASP 引物特异性结合的PCR 反应过程是Touchdown PCR,设定温度是55~61 ℃,所以引物的Tm 值应尽量接近61 ℃。
两条引物的扩增效率主要被两条竞争性引物的Tm 值差值及序列长度差值影响,SNP 位点变异类型会影响两条竞争性引物的序列长度差。两条竞争性引物的Tm 值差值的调节可通过增减某一条竞争性引物5′端碱基的数量实现,但长度差要尽量小,此时可尽量选择SNP位点变异类型为G、C互变的类型。
本试验在引物筛选时出现的一类特殊的引物分型效果,即杂合基因型与纯合基因型混在一起,此类分型效果出现的可能性有二:一是由于引物筛选时F2代样本数量只有30 个,样本中并未出现第3 种基因型;二是这类标记本身是共显性标记,不能区分杂合子。由于本试验旨在找到高多态性的共显性标记,所以在多态性筛选时将这类标记归为不可使用。
本试验中,烘干DNA 体系反应体系更小、通量显著提高(由筛选引物的Quant Studio 12 k flex的1 板384 孔板提高为20 板384 孔板),而且从分型结果来看与引物筛选反应体系效果基本相同,甚至很大比例的标记表现出更好的分型效果,所以干体系的转化十分必要。其中干体系的荧光信号产生PCR 反应过程的循环数确定需要进行预实验,从34 个循环试起,每次增加3 个循环,观察分型结果,使分型结果尽量集中,但要保证NTC 在原点附近。
对于干体系的分型结果,标记筛选时表现为分簇分散的在干体系中,有43.7%的标记分簇效果显著提升;标记筛选时表现理想的、分簇分散的、杂合纯合较近且聚类集中的标记,其分簇情况基本不变;而引物筛选时表现为杂合纯合较近且分簇分散的引物在干体系中分型失败率较高,由此说明本试验在挑选用于高通量反应体系转化的标记时,所采用的分簇集中优于分簇分散,分簇分散优于杂合纯合靠近,杂合纯合靠近的引物中分簇集中优于分簇分散的选择原则是正确的。
通过由207 个辣椒品种组成的群体的基因分型结果,证明本试验设计的130 个辣椒的KASP 标记具有高多态性、强普适性,并且适用于高通量基因分型平台。
本试验开发了一组包含130 对辣椒KASP 引物且均匀分布在辣椒12 条染色体上的分子标记集,其多态性高,普适性强,且适用于高通量基因分型。
