日本脑炎病毒NS1蛋白致脑血管内皮细胞损伤研究
2023-11-26潘俊慧刘亚林梁真洁杨兴淼谢盛达曹瑞兵
潘俊慧,刘亚林,梁真洁,杨兴淼,谢盛达,曹瑞兵
(南京农业大学动物医学院,江苏 南京 210095)
日本脑炎病毒(Japanese encephalitis virus,JEV)是一种蚊媒传播的单股正链RNA病毒,属于黄病毒科黄病毒属[1]。水禽和猪是JEV重要的中间宿主和扩增宿主。JEV感染可导致人和马发生脑炎,种猪感染后可表现为母猪流产、公猪睾丸炎,对养殖业造成重大损失[2-3]。JEV基因组的开放阅读框(ORF)编码一个多聚蛋白,经病毒和宿主蛋白酶切割成3种结构蛋白(C、prM和E)和7种非结构蛋白(NS1、NS2A、NS2B、NS3、NS4A、NS4B和NS5)[4]。
JEV非结构蛋白1(NS1)由352个氨基酸组成,其相对分子质量为(40~50)×103,具体的大小取决于蛋白的糖基化状态。JEV NS1蛋白可形成单体、二聚体和六聚体[4],位于胞质中的单体和二聚体蛋白具有促进病毒组装的功能,而六聚体蛋白是一种分泌蛋白,会被感染的细胞分泌至宿主体液循环系统[5]。已有研究报道,JEV NS1可与异质核糖核蛋白K(hnRNP K)、波形蛋白(vimentin)相互作用进而调节病毒的复制[6]。JEV NS1蛋白免疫小鼠后可提供90%的保护率[7-10],为新型JEV疫苗的研制提供了科学思路。登革热病毒(Dengue virus,DENV)的分泌性NS1蛋白(sNS1)能够增强蚊子通过吸血方式对病毒的摄取,而针对NS1蛋白的特异性抗体能抑制DENV从动物传播到蚊子[1]。目前,JEV sNS1蛋白的生物学功能尚未阐明。
糖萼(endothelial glycocalyx,EGL)覆盖在血管腔内皮细胞表面,是血管屏障的重要成分。EGL主要通过蛋白聚糖(主要为糖胺聚糖)和糖蛋白(包含酸性寡糖和唾液酸寡糖,是寡糖链与多肽链共价相连所构成的复合糖)与内皮相连并形成一个网络结构。糖胺聚糖(glycosaminoglycans,GAG)是糖萼最重要的“主链”分子。在血管内皮上主要存在5种糖胺聚糖链:硫酸乙酰肝素(heparan sulfate,HS)、硫酸软骨素(chondroitin sulfate,CS)、硫酸皮肤素(dermatin sulfate,DS)、硫酸角质素(keratan sulfate,KS)和透明质酸(hyaluronic acid,HA)。EGL的结构对维持血管内皮的完整性至关重要。有报道称,DENV sNS1通过激活内皮细胞溶酶体中的唾液酸酶来水解EGL中的唾液酸修饰,sNS1蛋白能够激活组织蛋白酶L和乙酰肝素酶的级联反应导致HS从EGL脱离,总之,sNS1蛋白通过这2条途径实现对EGL的破坏,从而损伤血管内皮并造成血浆渗漏[11]。另外,其他黄病毒如寨卡病毒,也能利用这种机制破坏血管内皮[5],但JEV NS1蛋白是否具有损伤内皮细胞的生物学功能尚不清楚。
JEV NS1是一种糖基化修饰的蛋白,含Asn-130(N130)和Asn-207(N207)2个保守的N-糖基化位点,西尼罗病毒(WNV)的NS1蛋白除上述2个糖基化位点外还有第3个Asn-175(N175)糖基化位点[12]。鉴于JEV感染机体会导致脑炎的发生[13],为探究JEV sNS1蛋白的生物学功能,本研究拟通过昆虫细胞杆状病毒表达系统获得JEV NS1蛋白及其N207糖基化位点突变蛋白,再以人脑微血管内皮细胞(HBMEC)为模型,通过检测细胞表面的唾液酸修饰和胞内组织蛋白酶L的表达水平来评价NS1蛋白对内皮细胞的损伤情况,并利用小鼠试验进一步探究sNS1蛋白对血脑屏障的影响,也为优化JEV亚单位疫苗候选抗原奠定理论基础。
1 材料与方法
1.1 细胞与毒株
sf9昆虫细胞、High5细胞、人脑微血管内皮细胞系(HBMEC)、供体载体pFastBac1、感受态细胞DH5α、DH10Bac均由本实验室保存。
1.2 主要试剂
高糖DMEM细胞培养基、胎牛血清、细胞培养基Grace insect medium和SF-900Ⅱ SFM及Cell FectinⅡ昆虫细胞转染试剂均购自Invitrogen公司;蛋白胶配制试剂盒购自雅酶生物有限公司;细胞培养板和细胞培养瓶购自无锡耐思生命科技股份有限公司。
1.3 JEV NS1和NS1 N207Q基因的扩增
以真核表达质粒pVAX-NS1为模板[14],参考JEV的基因组序列(GenBank:GQ918133.2)并使用Primestar高保真扩增酶分别扩增包含有助于蛋白分泌的蜂肽基因序列[15](下划线)的JEVNS1和NS1N207Q的目的片段。引物序列通过Oligo 7软件设计,由南京擎科生物科技有限合成。引物对序列如下:
NS1 F/R:5′-CGCGGATCCGCCACCATGGATGAAATTCTTAGTCAACGTTGCCCTTGTTTTTATGGTCGTGTACATTTCTTACATCTATGCGACCGATCAATT-3′/5′-TTGTCGAGACTGCAGGCTCTAGACTAATGGTGATGGTGATGATGAGCATCAACCTGCGATCTG-3′;N207Q F/R:5′-GGATTGAGAGTCGCTACCAAGACACATGGAAACT TG-3′/5′-CAAGTTTCCATGTGTCTTGGTAGCGACTCTCAATCC-3′。
1.4 重组质粒pFast-NS1和pFast-NS1 N207Q的构建
1.4.1 重组质粒pFast-NS1的构建经鉴定成功的NS1目的片段和载体pFastbac1采用BamHⅠ和XbaⅠ双酶切,纯化后使用T4连接酶进行连接,再转化到感受态DH5α,从培养平板中挑取单一菌落进行鉴定。
1.4.2 重组质粒pFast-NS1N207Q的构建以pVAX-NS1为模板,通过PCR扩增获得NS1N207Q-1/NS1N207Q-2,与载体pFastbac1通过同源重组的方法进行连接(图1)。使用同源重组酶将线性化的载体片段和纯化的NS1N207Q片段进行反应。反应体系如下:pFastBac1酶切纯化片段110 ng,NS1N207Q-1纯化片段12 ng,NS1N207Q-2纯化片段11 ng,5×CE MultiS Buffer 4 μL,Exnase MultiS 2 μL,ddH2O补至 20 μL。使用PCR仪,37 ℃反应30 min。
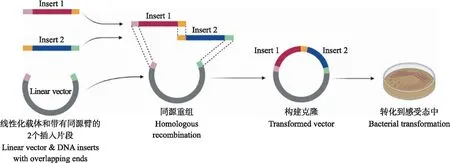
图1 同源重组克隆技术步骤Fig.1 Procedure of homologous recombination cloning technology
1.5 重组杆粒rBacmid-NS1和rBacmid-NS1 N207Q的构建
取鉴定正确的pFastBac1-NS1和pFastBac1-NS1N207Q重组质粒1 μL,加入新鲜制备的感受态DH10Bac/E.coli中,冰浴30 min;置于42 ℃水浴锅中热激90 s,迅速冰浴2 min;向其加入800 μL的液体LB培养基,置于37 ℃摇床,220 r·min-1培养4 h;在新的1.5 mL EP管中,用无抗性的液体LB培养基与培养好的菌液按照10倍比进行稀释,然后将菌液均匀涂在含卡那霉素、四环素和庆大霉素的3种抗生素的平板中,置于37 ℃培养48 h;在蓝白斑板上取单个白色菌落,重新划线得到纯化的白色菌落,经PCR鉴定成功后获得重组杆粒。
1.6 重组杆状病毒的制备
将sf9细胞用移液管吹下来进行计数,然后均匀铺到24孔板;待24孔板的sf9细胞生长密度为80%~90%时,开始转染。具体操作:取1 μg重组杆粒加到100 μL双无抗的Grace Medium;取Cell FectinⅡ 6 μL加入100 μL双无抗的Grace Medium中,混匀;将稀释好的杆粒与转染试剂混匀,室温孵育40 min。同时,用无菌PBS溶液清洗3次24孔细胞板里的细胞后弃洗液;将Grace Medium加入杆粒和转染试剂的混合液中,吹吸混匀,最后将混合物加入细胞孔中,27 ℃继续培养5 h后换液,24孔细胞板加入500 μL sf-900Ⅱ SFM,将细胞板置于27 ℃培养箱中培养,直到病变产生,收取病毒液。
1.7 杆状病毒的传代及扩增
将转染后收集的病毒液作为P1代病毒液,在4 ℃条件下4 000g离心15 min,然后将上清液进行过滤。收取P1代毒液,一部分储存在4 ℃冰箱备用,一部分储存在-80 ℃冰箱中长期保存,剩下的病毒液继续传代。将P1代病毒按照1∶10体积比感染sf9细胞,27 ℃恒温培养箱培养72 h,根据细胞病变情况收集病毒液,然后将其在4 ℃条件下4 000g离心15 min,收取P2代病毒液,置于4 ℃避光保存;将P2代病毒液按照上述相同的操作大量扩增获得P3代病毒液,按照20%的比例感染High5细胞并悬浮培养,72 h收集上清液,使用亲和层析的纯化方式获得目的蛋白。
1.8 NS1蛋白致人脑血管内皮细胞(HBMEC)损伤的细胞表面分子唾液酸修饰的检测
向细胞生长密度为80%的细胞培养皿中加入终浓度为5 μg·mL-1的NS1和NS1 N207Q蛋白,孵育 6 h 后通过检测WGA的荧光强度衡量HBMEC细胞表面的唾液酸修饰含量。WGA是一种小麦胚芽凝集素,能够选择结合N-乙酰唾液酸残基,其在溶液中以异二聚体的形式存在,相对分子质量约为38×103。避光操作下用PBS将WGA试剂稀释至5.0 μg·mL-1;取200 μL加入细胞培养皿中,置于5% CO2细胞培养箱中,37 ℃ 避光孵育30 min;经PBS清洗后用4%的多聚甲醛固定,经DAPI染核后在荧光显微镜下观察荧光强度。
1.9 NS1蛋白致HBMEC损伤的组织蛋白酶L含量变化检测
向细胞生长密度为80%的细胞培养皿中加入终浓度为5 μg·mL-1的NS1和NS1 N207Q蛋白,孵育6 h后将培养皿中的细胞固定,再经0.1% Triton-100破膜处理后以组织蛋白酶L特异性单抗为一抗进行孵育,共聚焦显微镜下检测细胞内的组织蛋白酶L含量的变化。
1.10 NS1蛋白致小鼠血脑屏障(BBB)损伤检测
为观察JEV NS1蛋白对HBMEC损伤致小鼠BBB的影响,参照文献[5]将4周龄Balb/c雌鼠分为3组,分别尾静脉注射NS1、NS1 N207Q和PBS。每只注射剂量为10 mg·kg-1。注射后每天记录小鼠的临床症状和精神状态。分别在注射24、48和72 h后,经尾静脉注射200 μL的1%伊文思蓝染料,1 h后用麻醉剂进行麻醉,并用30 mL生理盐水进行心脏灌注,剖取小鼠脑部,观察小鼠脑部的伊文思蓝染色情况。
2 结果与分析
2.1 表达JEV NS1蛋白的重组杆状病毒的鉴定
外源蛋白在昆虫细胞中分泌效率通常较低,在NS1的N端加入蜂肽序列可提高分泌效率,并在蛋白分泌之后切割蜂肽,这样不影响NS1蛋白的生物功能[15]。设计JEVNS1基因片段的上游扩增引物并引入蜂肽序列,经PCR扩增获得JEVNS1基因,经10 g·L-1核酸胶电泳检测,获得的1 099 bp目的条带与预期NS1大小一致(图2-A)。用限制性内切酶BamHⅠ和XbaⅠ进行载体线性化,将NS1片段克隆到杆状病毒表达体系的供体质粒pFastBac1。挑选阳性菌落,提取质粒进行BamHⅠ和XbaⅠ双酶切鉴定,条带大小与预期结果相符(图2-B),测序后确定已成功获得pFastBac1-NS1重组质粒。

图2 表达JEV NS1蛋白的重组杆状病毒的鉴定Fig.2 Identification of recombinant baculovirus expressing JEV NS1 protein A.NS1目的片段的鉴定及扩增;B.重组质粒 pFast-NS1的酶切鉴定;C.重组杆粒rBacmid-NS1菌液的PCR鉴定;D. sf9细胞表达NS1蛋白的间接免疫荧光鉴定;E.Western blot和SDS-PAGE检测NS1蛋白(Mock:空白对照)。A. Detection and amplification of NS1;B.Identification of pFast-NS1 by double restriction enzyme digestions;C. Identification of rBacmid-NS1 by PCR;D. Indirect immunofluorescence identification the expression of NS1 in sf9 cells;E. The analysis of recombinant of NS1 in Western blot and SDS-PAGE(Mock:Black control).
将鉴定成功的重组穿梭质粒转座到感受态DH10Bac/E.coli,通过四环素、卡那霉素和庆大霉素的3种抗性和蓝白斑筛选得到白斑的rBacmid。使用M13上、下游引物进行菌液PCR鉴定,得到大小约3 350 bp的核酸条带,与编码NS1蛋白的基因片段的大小相符。由于目的片段较长,退火温度的设置和引物匹配灵敏度使用普通扩增酶进行扩增时出现非特异性结合,导致在约1 700 bp处出现非特异性条带(图2-C),以上结果表明rBacmid-NS1成功构建。
将rBacmid-NS1转染sf9细胞,72 h后收获杆状病毒P1代。将P1的杆状病毒进一步扩增获得P2代。P2代杆状病毒按照10%的体积比接种于sf9细胞,72 h后对重组病毒进行免疫荧光检测(IFA)和Western blot鉴定。固定sf9细胞,用NS1蛋白单克隆抗体作为IFA鉴定的一抗,在倒置荧光显微镜下观察,重组杆状病毒接种的细胞能够产生特异性的红色荧光,而对照孔的sf9细胞孔则无明显荧光(图2-D),表明拯救的杆状病毒能够有效表达NS1蛋白;Western blot能够检测到相对分子质量45×103左右的NS1条带(图2-E)。综上,成功制备了能表达NS1蛋白的重组杆状病毒。
2.2 NS1及其突变体NS1 N207Q蛋白的表达及纯化
将P3代NS1重组杆状病毒分别按照5%和10%的体积比接种High5细胞,通过对上清液样品进行鉴定,发现接毒量为10%且收样时间为72 h的上清培养基中表达的NS1蛋白含量最多,因此后续的NS1蛋白表达选择接毒量为10%的比例,收样时间为72 h(图3-A)。

图3 NS1和NS1 N207Q蛋白的表达与纯化Fig.3 Expression and purification of NS1 and NS1 N207Q proteins A. NS1重组杆状病毒感染High5最优表达鉴定;B. Western blot鉴定区分NS1和NS1 N207Q;C. 间接免疫荧光鉴定sf9细胞表达NS1 N207Q蛋白;D. SDS-PAGE鉴定纯化的NS1蛋白;E. SDS-PAGE鉴定纯化的NS1 N207Q蛋白;F. Western blot鉴定纯化的NS1蛋白;G. Western blot鉴定纯化的NS1 N207Q蛋白。1~9/1′~8′:150 mmol·L-1咪唑洗脱液,10~13/10′~11′:300 mmol·L-1咪唑洗脱液。A. Identification of optimal expression of High 5 infected by recombinant baculovirus NS1;B. The molecular weight difference between NS1 and NS1 N207Q was determined by Western blot;C. Indirect immunofluorescence identification the expression of NS1 N207Q in sf9 cells D.The purified NS1 protein was identified by SDS-PAGE;E. The purified NS1 N207Q protein was identified by SDS-PAGE;F. The purified NS1 protein was identified by Western blot;G. The purified NS1 protein was identified by Western blot. 1-9/1′-8′:150 mmol·L-1 imidazole eluent,10-13/10′-11′:300 mmol·L-1 imidazole eluent.
本研究使用1 L的细胞摇瓶悬浮培养High5细胞200 mL,收集细胞培养上清液,经蛋白亲和柱纯化并通过SDS-PAGE和Western blot鉴定,发现JEV NS1蛋白在150 mmol·L-1咪唑洗脱下获得条带单一的NS1蛋白,而300 mmol·L-1咪唑洗脱下NS1蛋白的条带较粗,存在杂蛋白,说明NS1蛋白得到有效纯化(图3-B、C)。
为了后续探究JEV NS1蛋白特异性的生物学功能,在有效制备出JEV NS1蛋白的条件下,又通过点突变的方式突变其N207糖基化位点,经鉴定后成功拯救出NS1突变蛋白NS1 N207Q的重组杆状病毒,且突变蛋白的分子质量要略小于NS1蛋白的分子质量(图3-D、E)。NS1 N207Q重组杆状病毒感染High5细胞,收集上清液并纯化,再经SDS-PAGE和Western blot鉴定,可知NS1 N207Q蛋白得到有效纯化(图3-F、G)。
2.3 NS1蛋白致HBMEC细胞表面唾液酸和组织蛋白酶L含量变化的检测
为了探究JEV NS1蛋白的生物学功能,选用昆虫细胞-杆状病毒系统获得真核NS1蛋白及其突变蛋白。前期的探究中发现向HBMEC中添加NS1蛋白导致细胞间隙变大且细胞状态较差的现象,由于内皮细胞表面的保护屏障主要为糖萼(EGL),唾液酸是EGL的重要成分[5],因此,通过共聚焦的方式检测NS1蛋白对细胞表面唾液酸修饰的影响。将HBMEC接种培养皿,待细胞密度达到100%时,添加5 μg·mL-1NS1作用6 h,使用WGA荧光染料染色后固定细胞,在荧光显微镜下观察细胞表面分子唾液酸的修饰,我们发现NS1蛋白作用HBMEC细胞6 h后,与空白对照组相比,添加NS1蛋白组细胞表面的唾液酸含量明显减少(图4-A)。

图4 人脑微血管内皮细胞(HBMEC)表面唾液酸含量(A)和细胞内组织蛋白酶L含量(B)检测Fig.4 Detection of the content of sialic acid on the HBMEC surface(A)and the content of cathepsin L(B)in HEMECWGA:唾液酸指示剂 Sialic acid indicator;DAPI:细胞核染色 Nuclear staining.
由于EGL的另外一个重要成分为硫酸乙酰肝素蛋白聚糖,而组织蛋白酶L是降解硫酸乙酰肝素蛋白聚糖的重要前体蛋白酶[16]。为了进一步探究NS1蛋白对EGL的影响,我们又检测了NS1蛋白处理后的HBMEC中组织蛋白酶L含量。在共聚焦显微镜下观察发现,与空白对照组相比,添加NS1蛋白组细胞中组织蛋白酶L含量增多(图4-B)。
根据以上试验结果可得,JEV NS1蛋白具有下调脑内皮细胞表面唾液酸修饰的功能,并且NS1蛋白可诱导细胞内组织蛋白酶L的表达并通过激活硫酸乙酰肝素酶降解硫酸乙酰肝素最终造成EGL的损伤。为了探究该功能是否为NS1蛋白的特异性生物学功能,我们又将NS1突变蛋白NS1 N207Q以相同的方式作用HBMEC,6 h后检测内皮细胞表面的唾液酸修饰和细胞内组织蛋白酶L含量。检测结果表明,与对照组相比,添加NS1 N207Q蛋白组细胞的唾液酸修饰含量和组织蛋白酶L含量均无明显变化。因此,JEV NS1蛋白可降解HBMEC细胞表面的EGL并对内皮屏障造成损伤。
2.4 JEV NS1及NS1 N207Q蛋白对小鼠血脑屏障功能的影响
既然JEV NS1蛋白对于内皮细胞有明显损伤作用,且内皮细胞又是血脑屏障的重要组成成分,于是我们思考内皮细胞损伤是否对血脑屏障功能产生影响。参照已有文献DENV NS1蛋白尾静脉接种剂量为10 mg·kg-1[5]进行相关试验操作。对试验小鼠进行临床症状观察:NS1蛋白接种组和NS1 N207蛋白接种组小鼠,在24、48和72 h的观察期内,小鼠无明显异常,精神状态良好,食欲饮水正常,接种部位未出现发热、红肿等异常现象;PBS接种组在72 h的试验观察期内正常。各组小鼠均未出现明显的临床症状。
如图5所示:小鼠心脏灌注生理盐水后取出小鼠脑部,注射后24~72 h,NS1蛋白组均能观察到蓝色染料,而NS1 N207Q蛋白组和PBS组均未观察到明显的蓝色染料。此结果表明,NS1蛋白在注射后24和48 h即对小鼠的血脑屏障造成损伤,而72 h后对小鼠的血脑屏障的损伤减弱,而注射NS1 N207Q蛋白组小鼠血脑屏障无明显损伤,PBS组正常。

图5 NS1蛋白致小鼠血脑屏障损伤的染色观察Fig.5 Observation of brain staining on blood-brain barrier injury caused by NS1 protein in mice
3 讨论
本试验在探究JEV NS1蛋白生物学功能时发现,向HBMEC中添加NS1蛋白,细胞间隙变大且细胞状态较差。体外试验结果显示,JEV NS1蛋白能够下调HBMEC表面分子的唾液酸修饰,并且能够上调细胞内组织蛋白酶L的表达,表明NS1蛋白具有损伤脑内皮细胞糖萼的生物学功能。伊文思蓝属于一种常用的偶氮染料制剂,在血液中与血浆白蛋白有很高的亲和力,因此在科学研究中常被用于示踪剂观察血脑屏障(BBB)的完整性[17]。动物试验表明,注射NS1蛋白后小鼠的脑部观察到蓝色的伊文思蓝染料。由此判断JEV NS1蛋白具有损伤小鼠血脑屏障的功能。但在作用时间为72 h时,NS1蛋白损伤血脑屏障的作用减弱。其原因可能是DENV NS1蛋白作用内皮细胞是一个动态变化的过程[12],NS1蛋白作用的前期效果较为明显,后期其作用相对较弱;注射到小鼠体内的NS1蛋白随着时间的增加,也会在小鼠体内逐渐被降解。有研究指出,蛋白的糖基化对于其生物学功能有重要影响[5]。为了探究JEV NS1蛋白损伤内皮功能的特异性,我们将NS1蛋白的N207糖基化位点进行突变并获得突变蛋白NS1 N207Q。试验结果显示,JEV NS1 N207Q处理后HBMEC表面的唾液酸修饰和组织蛋白酶L含量均无明显变化,说明NS1 N207Q蛋白处理对小鼠的血脑屏障无明显损伤。由此可得,JEV NS1蛋白N207位点对其损伤脑内皮的生物学功能具有重要作用。
JEV NS1是一个多功能的非结构蛋白,对病毒的增殖有重要作用,其主要参与病毒复制复合体的形成,具有促进病毒复制的功能[18-19]。此外,NS1蛋白还参与机体的免疫相关因子的互作[20]。JEV感染细胞后产生的NS1蛋白以含有脂质物的桶状六聚体形式分泌,并进入到血液循环中,因此其可以用作诊断抗原和病毒感染严重程度的生物标志物[21]。有研究提出NS1基因作为核酸疫苗免疫小鼠,可以保护90%小鼠抵抗致死剂量的JEV感染[22]。然而,目前针对JEV分泌的NS1蛋白的生物学功能研究较少。有研究报道,DENV NS1蛋白直接与血管内皮细胞相互作用,导致血管内皮糖萼受到破坏,而内皮糖萼层(EGL)维持内皮屏障完整性至关重要[23]。因此,本试验制备了分泌性JEV NS1蛋白并通过体外和体内实验结果证明JEV NS1蛋白可损伤HBMEC并影响小鼠的血脑屏障,而其突变蛋白NS1 N207Q损伤HBMEC的功能丧失。
研究报道,登革热病毒(DENV)和寨卡病毒(ZIKV)的NS1蛋白可以促进蚊子感染病毒,而NS1蛋白抗体可通过Fc依赖性的方式结合体液中及细胞膜上的NS1蛋白清除被病毒感染的细胞[24-25]。因此,病毒的NS1蛋白被认为是新型疫苗的潜在候选抗原。研究表明,用关键片段缺失的NS1蛋白免疫小鼠既可以降低损伤宿主血管的副作用,也能保护小鼠对抗DENV的致死性感染;同时也能减少病毒对蚊子的感染,从而减弱DENV的自然传播链,这可能为控制黄病毒在自然界中的传播提供新途径[1]。因此,JEV NS1蛋白可作为一种既可以提供免疫保护又可以阻断病毒的跨宿主传播的双效抗原。
虽然JEV NS1具有一定免疫原性[9,26-27],但是NS1蛋白本身会诱发内皮细胞损伤,无法直接作为候选亚单位疫苗的储备蛋白,而NS1 N207Q蛋白则无明显损伤作用,可以作为候选免疫原进行进一步的探究。
本试验根据氨基酸序列的分析,将207糖基化位点进行突变,制备出对内皮细胞无明显损伤作用的NS1 N207Q蛋白。由于分泌到上清液中的重组NS1 N207Q蛋白的纯化效率较低,后续研究需要进一步对蛋白序列进行优化或选择添加双标签以及使用亲和层析与分子筛组等纯化模式以提高纯化蛋白的效率[13-14],也可通过缺失具有损伤功能的目的片段而获得安全有效的缺失NS1蛋白,可结合E蛋白制备既产生中和抗体又产生NS1抗体的二联苗。
综上所述,本研究通过人脑内皮细胞损伤试验和小鼠血脑屏障通透性试验,结果表明JEV NS1蛋白能够引起脑内皮的损伤,而NS1 N207Q突变蛋白对内皮的损伤作用显著减弱。本研究为JEV新型亚单位疫苗的研制提供新思路和物质基础。
