细叶云南松林大径木择伐后的遗传多样性动态及小尺度空间遗传结构
2023-11-25黄春晖白天道李远发黄厚宸郑党斌蒋维昕
黄春晖,白天道,李远发,黄厚宸,郑党斌,蒋维昕
(1.广西大学 a.广西森林生态与保育重点实验室;b.林学院,广西 南宁 530004;2.广西壮族自治区国有雅长林场,广西 百色 533000)
空间遗传结构是指不同基因型个体在空间上的非随机分布[1],反映了物种/种群自身的生物学特性、生态适应性、生境变迁与自然选择效应[2]。植物种群的空间遗传结构主要取决于物种繁育机制、花粉和种子的散播方式、遗传漂移、微生境选择及植物非随机空间分布等因素[3]。通常来说,以风媒传粉和风力传播种子的松柏类植物,其个体基因型空间分布接近于随机分布[4],而当花粉、种子传播范围小于植物种群面积时,小尺度空间遗传结构十分明显[5]。对华木莲Sinomanglietia glauca[6]和水青树Tetracentronsinense[7]的小尺度空间遗传结构研究均发现,有限的基因流是居群形成空间遗传结构的主要原因。相反,种子具翅的植物,如鹅掌楸Liriodendronchinense[8],可借助风力传播种子,基因流较强,一定程度上降低了其空间遗传结构强度。小尺度空间遗传结构影响着植物种群、交配系统、有效种群大小、近交程度以及种群对微生境变化的适应能力,进而影响植物种群的动态[9]。通过分析种群空间遗传结构,可以有效地获得种群历史、生活史特征、遗传漂变、基因流和其他遗传过程的信息,有助于探讨各种进化因素间的作用,从而为科学制定种群的利用和保护策略奠定理论基础。
择伐是实现森林可持续经营的重要措施,在林业生产中已持续超过半个世纪[10]。研究表明,择伐通过移除部分成年树木改变了林分的竞争环境,调节了林木间的资源分配,对林地环境、林木生长和林下更新有显著影响[11-15]。择伐通过剔除部分基因型,很可能会对林分的基因构成及空间遗传结构产生影响。有研究认为,择伐会降低后续更新群体的遗传多样性[16],也有研究发现择伐和未择伐群体在遗传多样性和空间结构上无显著差别[17-18],但择伐后保留个体有等位基因频率减少和丢失的趋势[19]。许玉兰等[20]对不同择伐条件下的云南松Pinusyunnanensis遗传多样化研究发现,不同的择伐方式对云南松群体遗传多样性影响并不显著;周祺[21]对色木槭Acermono天然种群的研究发现,人为抚育或干扰后保留适当的种群个体数则物种遗传多样性不会受到较大影响,同时有利于调控种群的空间遗传结构。尽管这些研究从人为经营对林分遗传多样性影响的角度进行了有益的探索,但总体上择伐(尤其以收获为主的大径木择伐)后林分遗传多样性和空间遗传格局会发生何种变化,目前所知十分有限。
细叶云南松Pinusyunnanensisvar.tenuifolia被认为是云南松适应干热河谷气候的一个地理变种,仅分布于中国西南地区红水河、南盘江、北盘江下游地带,属于微域分布的植被类型[22]。在南盘江流域森林中,细叶云南松作为上层优势树种,在区域生态系统中占有重要地位[23]。近年来,该区域其他经济林(板栗Castaneamollissima、油茶Camelliaoleifera等)的迅速发展使得细叶云南松天然林受到大量蚕食和破坏,林分面积不断衰减[24]。目前,对细叶云南松林分的研究主要包括群落结构与动态[25]、空间分布[26]、种内和种间竞争[27]、木材特性[28]、遗传多样性[29-30]等方面。而择伐等人为扰动对细叶云南松林分遗传多样性及小尺度空间遗传结构是否造成影响目前尚未见研究报道。本研究以细叶云南松天然次生种群为对象,基于nSSR标记探究该林分在早年(1970年)经大径木强度择伐(拔大毛)后,自然恢复至今(2019年)的林分遗传多样性变化及其小尺度空间遗传格局。试图从遗传角度揭示大径木择伐对林分恢复的影响,进而为细叶云南松天然林资源保育和人工林可持续经营提供理论基础。
1 材料与方法
1.1 供试材料及采样
样品采集于广西雅长兰科植物国家级自然保护区花坪样地(106°23′11″E,24°49′54″N,平均海拔约1 254 m)。据雅长林场记载,该林分于1970年进行了一次经营性采伐,即伐掉干形通直的成年大树(胸径≥20 cm),保留未达采伐标准的小树,其后林分自然恢复至今,未受到大规模人为干扰。2005年该林地被划为自然保护区,使该林分的保护和恢复得以进一步加强。2016年,因研究需要在该林分代表性区域建成了一块永久性标准固定样地(100 m×80 m),样地内上层优势树种为细叶云南松、铁坚油杉Keteleeriadavidiana、栓皮栎Quercusvariabilis以及西南桦Betulaalnoides等,样地详细情况参考李远发等[26]的相关报道。基于全站仪(南方测绘,NTS-372 R10)获取样地内每株细叶云南松相对坐标(x,y,z),总计46株,胸径均在10 cm以上[27]。2019年8月,采集样方内全部个体的当年生针叶,放入自封袋经硅胶干燥后保存、备用。同时利用围径尺测定每株林木的胸径(表1)。

表1 参试细叶云南松个体胸径Table 1 DBH of P.yunnanensis var.tenuifolia
1.2 DNA提取
采用CTAB法提取细叶云南松针叶总DNA。利用微量核酸蛋白检测仪(ScanDrop200,德国)进行DNA质量和浓度测定,并用1%琼脂糖凝胶电泳检测其完整性。
1.3 nSSR-PCR扩增
参试标记来自马尾松P.massoniana基因组SSR[31]。经稳定性和多态性筛选后,最终选择15个标记用于群体扩增(表2)。PCR反应体系10 μL,其中包含DNA模板0.1 μL(50 μg·μL-1),10× Buffer 1 μL,Taq酶 0.1 μL,MgCl21 μL,dNTPs 1 μL,正反引物各 0.3 μL,ddH2O 6.2 μL[32]。扩增程序:94 ℃预热3 min,94 ℃变性30 s,50 ℃退火30s(表2),72 ℃延伸30 s,循环30次,最后72 ℃延伸10 min。PCR产物通过8%变性聚丙烯酰胺凝胶电泳后,利用运用GelAnalyzer 19.1[33]识别目的条带大小,以50 bp DNA ladder(Takara)为参照。
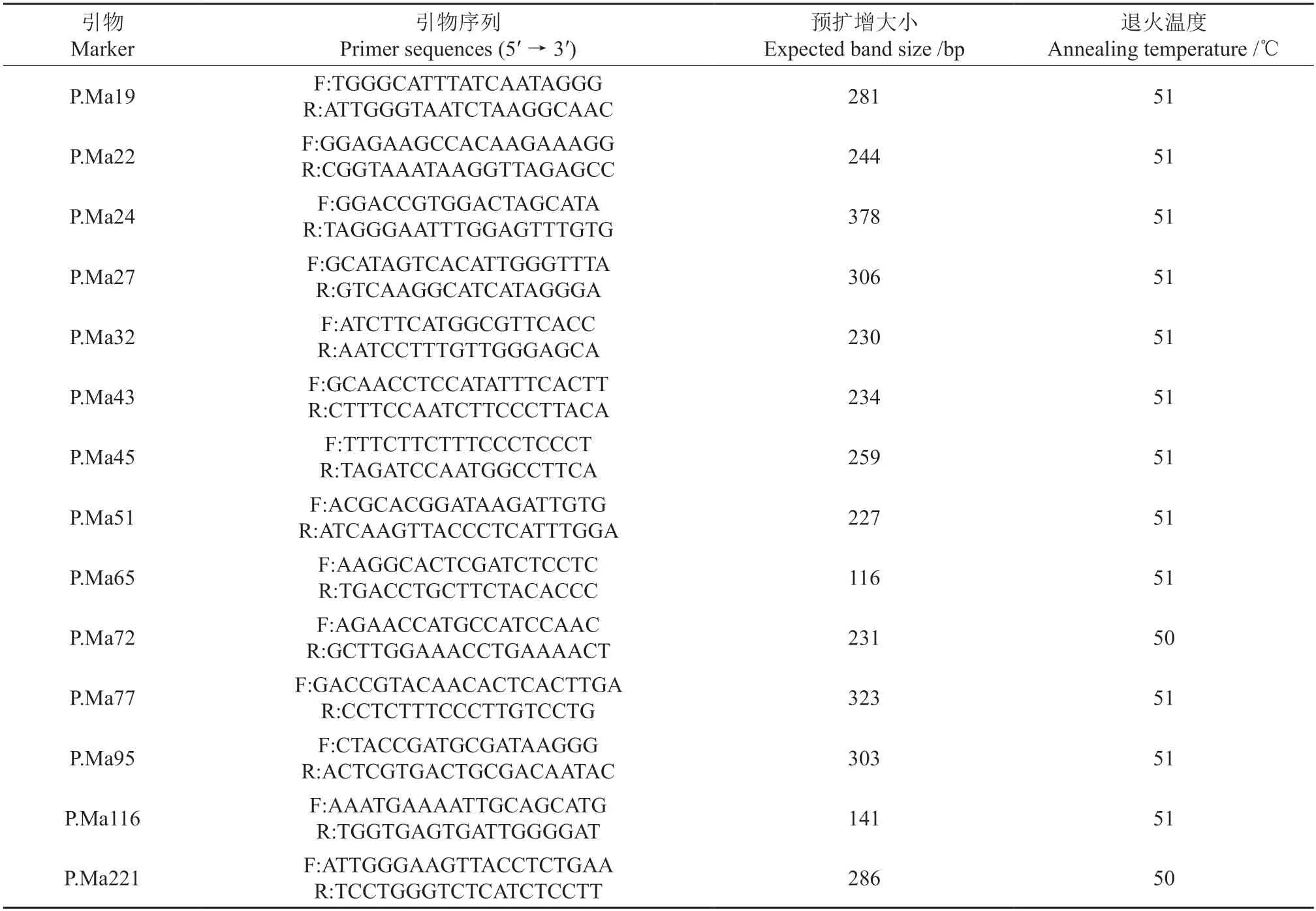
表2 nSSR位点序列及引物信息Table 2 Sequence and primer information of microsatellite loci
1.4 数据分析
数据经Excel 2016软件整理后,运用R 4.2[34]及其扩展包进行分析。并采用ggplot2[35]软件进行可视化。
1.4.1 大径木择伐后群体遗传多样性动态
为分析大径木择伐后49年内林分遗传多样性随时间的变化,基于直径大小与树龄的正相关关系,以及前人基于72年生细叶云南松解析木生长规律的研究报道[36],以其速生期(10~55 a)胸径平均年生长量(0.9 cm/a)为单位推算样方现有林木的更新时间,将样方内细叶云南松按胸径分为L(>45 cm,15个单株)、M(30~45 cm,19个单株)、S(<30 cm,12个单株)3组,其中L组代表伐前(树龄≥50 a)更新个体(0.9×50=45 cm),M组代表伐后15 a内(树龄34~49 a)更新个体(0.9×34=30.6 cm),S组代表伐后16 a至今(树龄≤33 a)的更新个体。分别计算各组样本的遗传多样性参数,包括等位基因数目(Na)、Shannon’s指数(I)、观测杂合度(Ho)、期望杂合度(He)、固定指数(F)等采用R扩展包poppr 和hierfstat[37]进行计算;并基于置换检验分析组间差异。
1.4.2 小尺度空间遗传结构
利用R包LEA[38]计算个体间共祖度(遗传结构),并结合样本坐标采用克里格法(Kriging)进行空间插值,呈现样方内个体间的空间遗传模式;对样方高程数据采用反距离权重法(Inverse distance weighting,IDW)插值,描述样方地形;利用R包adegenet[39]进行空间主成分分析(Spatial principal component analysis,sPCA),并基于蒙特卡洛模拟检测空间结构显著性。
2 结果与分析
2.1 DNA提取及PCR结果
微量核酸蛋白检测仪检测结果显示,DNA纯度为1.61~2.03,纯度较高;DNA浓度为129.41~1 006.58 ng/μL。1%琼脂糖凝胶电泳显示提取DNA具有较好的完整性,无明显拖尾和弥散(图1)。PCR产物电泳条带稳定清晰(图2),满足下一步分析要求。
图1 部分样本DNA琼脂糖凝胶电泳图Fig.1 DNA agarose gel electrophoresis diagram of some samples
图2 部分个体在位点P.Ma77的扩增结果Fig.2 Amplification bands of partial samples at locus P.Ma77
2.2 遗传多样性动态
参试细叶云南松群体平均等位基因数(A)为4.7个,Shannon’s指数(I)为1.209,平均观测杂合度(Ho)为 0.487,期望杂合度(He)为0.641,该群体具有较丰富的遗传多样性(表3)。但其固定指数显著大于零(F=0.234),表明群体可能存在一定程度的近/自交。哈迪温伯格平衡(HWE)检验表明,大部分位点(除了P.Ma95)均显著偏离HWE(P<0.05),表明群体存在非随机交配。
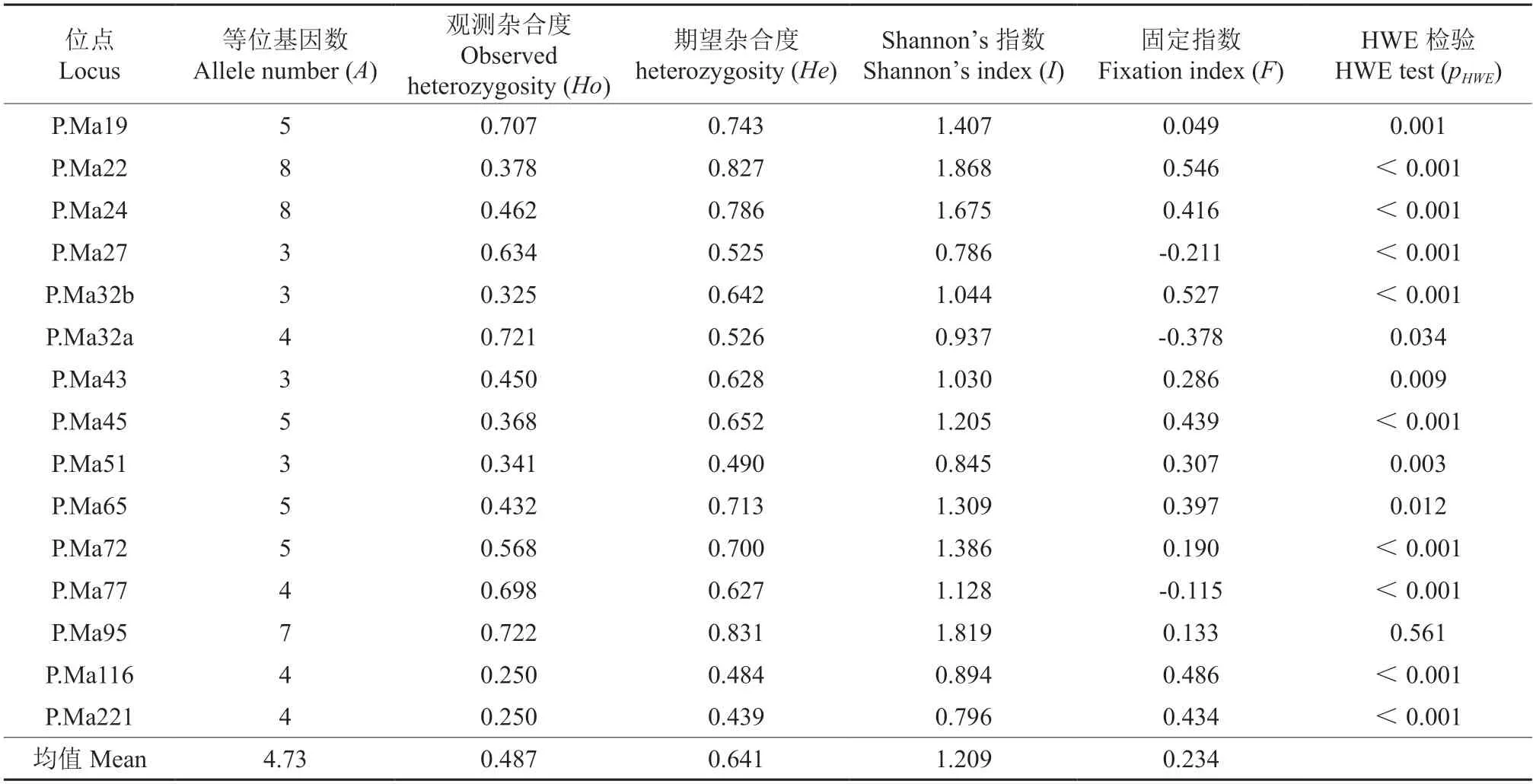
表3 参试细叶云南松群体在15个nSSR位点上的遗传多样性†Table 3 Genetic diversity of 15 microsatellite loci in P.yunnanensis var.tenuifolia
比较不同径级组的遗传多样性(图3),发现Ho有随径级减小而增加的趋势,L组与S组差异达到显著水平(P=0.02<0.05)。而He、I则表现为M组相对较小,且部分组间呈极显著差异(P<0.01)。尽管F值在组间未检测到显著差异,但其均值随径级减小呈下降趋势,表明该林分在大径木择伐后短期内遗传多样性有所下降,但随着时间的延长异交率和多样性逐步增加。
2.3 空间遗传结构
基于最小交叉熵值,参试个体nSSR基因型最佳分组数(K)为2(图4a)。nSSR共祖系数表明(图4b),群体内部分个体源于组间异交(含两种颜色),但也有部分个体可能源于近交或自交(呈现单一颜色)。
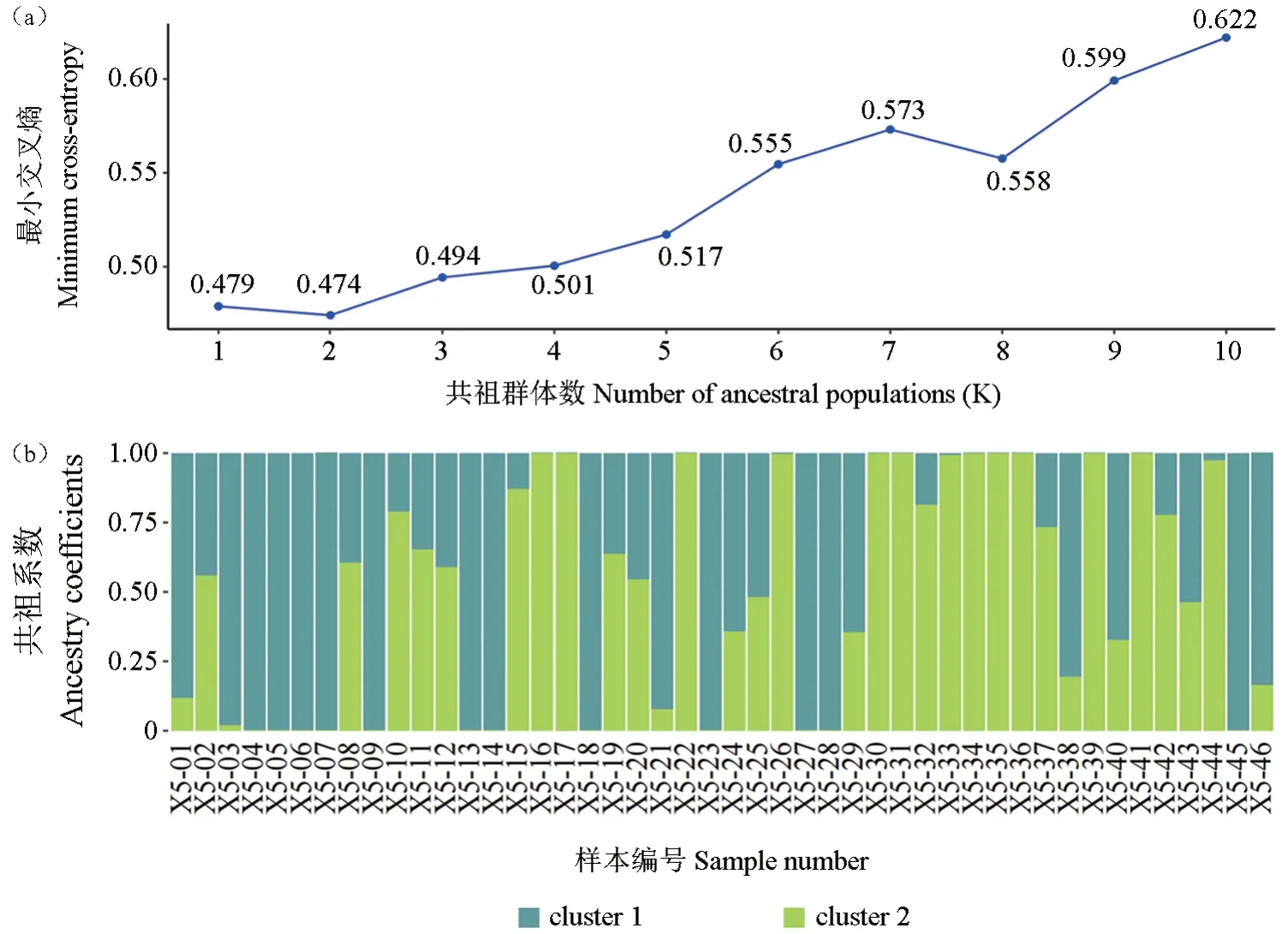
图4 参试细叶云南松样本的遗传结构Fig.4 The genetic structure of P.yunnanensis var.tenuifolia samples
将个体间共祖度结合个体空间坐标进行空间插值(图5),发现nSSR基因型第二组(cluster2)除了极少数个体(n=3)分布在样方的左上角,大部分个体(n=21)主要聚集分布在样地右下角,而第二组(cluster1)主要分布在样地中上部(图5a)。结合样方高程数据(图5b),nSSR大致为低坡位个体聚为一类,高坡位个体聚为另一类。基于nSSR基因型的空间主成分分析(sPCA)表明,第一主成分特征值占比较大,基于第一主成分的个体空间主成分得分分布模式与基于nSSR空间遗传分布模式类似(图5c)。
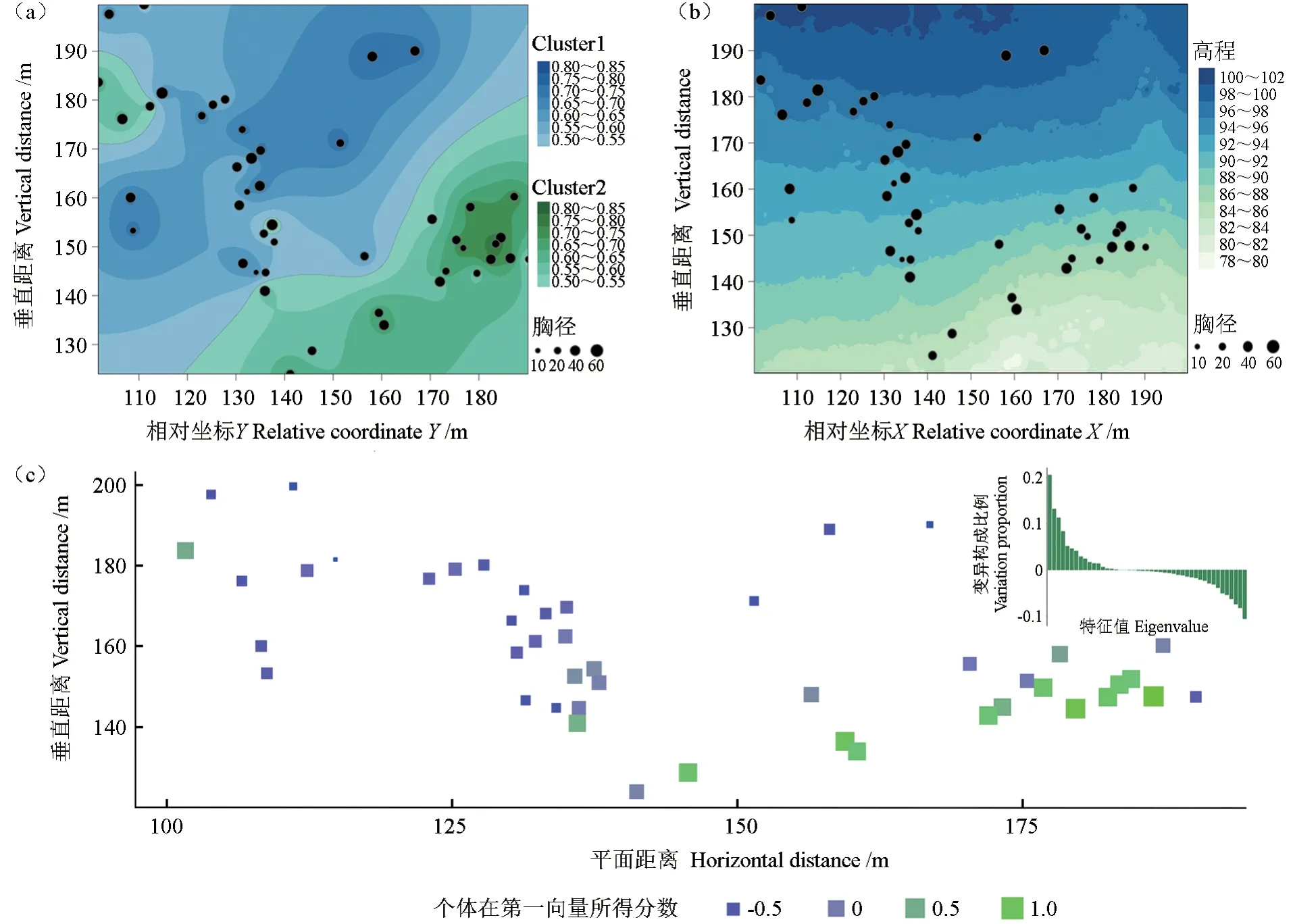
图5 基于个体遗传关系的空间插值及空间主成分分析Fig.5 Spatial genetic structure based on spatial interpolation and spatial principal component analysis (sPCA)
空间自相关分析表明,样地中细叶云南松在整体(图6a)和局部(图6b)尺度上均不存在显著的空间遗传结构(P>0.05)。这说明样方内个体尽管在遗传上似乎呈现一定的聚集分布,但个体间遗传距离与空间距离关联未达显著水平,表明林分现有遗传分布格局除了受到其花粉和种子自然传播的影响外,大径木择伐造成的母树空间格局改变可能是另一重要原因。

图6 细叶云南松样地在整体和局部尺度上的nSSR基因型空间自相关(基于1 000次蒙特卡罗模拟)Fig.6 nSSR genotype spatial autocorrelation analysis of P.yunnanensis var.tenuifolia on the global and partial scale in quadrat (based on 1 000 times Monte Carlo simulation).
3 讨 论
3.1 择伐对林分遗传多样性的影响
遗传多样性是生物适应复杂生境、维持群体稳定的基础。对以异交为主的林木而言,群体减少可能导致其自交率增加,引起衰退直至濒危[40]。择伐作为森林经营的重要措施,无论是以森林抚育为主的间伐还是收获为主的大径木强度择伐,都会改变林分内的个体数量及空间距离,这很可能对林分的基因构成和交配模式造成影响[41]。以往的一些研究表明,强度择伐会降低林分等位基因及基因型数量,造成群体漂变,使伐前及伐后群体遗传距离增大,但在有效等位基因、期望及观察杂合度、固定指数上的影响较小[17,42,19]。也有研究表明,林分成年个体(相对于幼年个体)具有较高的等位基因丰富度和较低的固定指数[43-44],表明不同树种在择伐后的遗传多样性响应有所不同[42]。本研究结果表明,细叶云南松大径木强度择伐后,随着自我修复(更新)时间的延长,林分的观测杂合度有逐步增加的趋势,而期望杂合度和Shannon’s指数呈先下降后增加的趋势,总体上,林分的异交率呈增加趋势。推测由于大径木被伐除后,残留少量生长及形质较差的植株(一般认为是近/自交产生的后代)成为伐后一定时间内林分的主要繁殖母树,林分有效繁殖群体规模减小,使得林分更新子代在一定时间内自交率有所增加,遗传多样性下降。同时这些劣质母树与伐前更新(尚未进入繁殖期)个体一起构成L组(图2),在一定程度上降低了伐前更新群体的遗传多样性。随着时间的延长(10~15 a后),林分内伐前更新个体进入繁殖期,林分内有效繁殖群体增加,林分的异交率增大,林分遗传多样性得以恢复。从当前林分来看,参试细叶云南松群体略高于其他一些松属树种的遗传多样性,如红松(Na=2.08,I=0.38,Ho=0.15,He=0.24)[45]、樟子松(Na=2.53,I=0.291,Ho=0.051,He=0.083)[46]、马尾松(Na=3.31,I=0.64,Ho=0.35,He=0.36)[47]等,说明当前细叶云南松群体遗传多样性得到了很好的恢复。
3.2 林分空间结构及其影响因素
分析空间遗传结构能有效了解物种遗传变异与居群或个体所处位置生境状况的相关性,有助于了解物种遗传多样性形成的原因,为物种保护提供科学依据[48]。对植物而言,花粉流和种子流是影响空间遗传结构强度的主要因素[49]。有限的花粉流和种子流会形成显著的空间遗传结构[50]。花粉传播方面,风媒花比虫媒花扩散范围更大;种子传播方面,风力传播比重力或鸟类传播范围更大[51]。依靠风媒传粉和风力传播种子的松柏类植物,其个体基因型空间分布接近于随机分布[52]。与其他松树类似,本研究中,细叶云南松在100 m尺度范围内未检测到显著的空间结构。其作为风媒植物,花粉粒小,具气囊且不具黏性,易随风传播。另外,细叶云南松球果成熟后通常存留在树上并开裂,使种子散落,其种子具种翅,可借助风力传播,大大提高了种子往远处扩散的可能[29]。
将nSSR空间遗传分布与样方高程分布模式进行对比发现两者具有一定的关联,高海拔区(大部分)个体聚为一组,低海拔区个体聚另一组,表明样地地形可能直接或间接影响(如风向、土壤肥力等)了当前林分的空间遗传格局。值得注意的是,尽管样方内个体间无显著空间自相关,但基于共祖系数及遗传距离的空间聚类分析表明,细叶云南松在样方内仍存在一定的聚集分布。这可能归因于大径木择伐后原有林分的随机分布格局被破坏,林分内非随机分布的母树间距离变大,在林木就近交配原则和种子流共同作用下,相邻个体间表现出一定的遗传相似性。
需要指出的是,本研究仅基于nSSR标记探究了100 m×80 m样方范围内细叶云南松小尺度遗传格局,要想更为准确地探究其小尺度遗传结构,可能需要在多点设置更大的样方,并结合其父系遗传的叶绿体标记(如cpSSR)进行林分内花粉流分析,以进一步揭示其空间遗传结构。
4 结 论
本研究以大径木强度择伐后自然恢复49 a细叶云南松天然群体为材料,探究了大径木强度择伐对后续更新群体的遗传多样性及空间遗传格局的影响。结果表明,大径木择伐后一定时间内林分的遗传多样性有所下降,但随着林分自然更新和恢复其遗传多样性逐步增加。空间遗传结构分析表明该群体在样方内呈现轻度聚集分布。大径木强度择伐一定程度上影响了林分空间格局,但在后期花粉及种子长距离风力传播的影响下,林分的空间结构(非随机分布)得到缓解。细叶云南松林具有较好的遗传多样性自我修复能力。
