缺氮胁迫下不同黑木相思基因型综合评价
2023-11-25陈天笑白小刚曾炳山陆钊华
陈天笑,白小刚,何 茜,胡 冰,曾炳山,陆钊华
(1.中国林业科学研究院热带林业研究所,广东 广州 510520;2.华南农业大学 林学与风景园林学院,广东 广州 510642)
在全球恶劣的气候变化下,非生物胁迫成为限制林木可持续生产的主要胁迫[1]。我国华南地区其土壤类型主要是红壤,土壤养分贫瘠,保肥能力差[2],再加上华南地区暴雨频发[3],土壤矿质营养淋失严重,使得林地土壤肥力退化,因此营养胁迫成为华南地区制约植物生长发育和林木生产力的主要因素之一。当今应对土壤养分匮乏的策略主要是化肥的使用和作物选育[4]。但由于林地生产者缺少科学的施肥技术和方法,常常出现化肥施用过量的情况,这不仅增加了生产成本,而且会对植物产生毒害作用,并且会对环境造成污染[5],所以通过营养学和遗传学的手段进行优良品种选育是一种行之有效的方法。
黑木相思Acaciamelanoxylon是含羞草科金合欢属的常绿大乔木,原产于澳大利亚东部和东南部。因其具有生长迅速、木材材性优良、适应能力强、改土性能好、能够调整树种结构、改善生态功能等特点,近年来随着其优良无性系的成功选育及在华南地区栽培面积逐年增加,是兼顾生态效益、社会效益和经济效益的多功能树种。但因其速生性,在生长发育过程中对矿质营养的需求量大,在实际推广过程中发现黑木相思表现出明显缺氮的症状,进而制约黑木相思人工林的生产力。由于不同黑木相思基因型的遗传特性存在差异,在应对氮缺乏生长形态表现不一,因此了解不同基因型应对缺氮胁迫的适应能力的差异,能够为黑木相思早期选育优良基因型提供理论基础。
氮素是植物生长发育过程中必需的大量元素之一,也是蛋白质、氨基酸、叶绿素和激素等物质的必需成分[6-8]。氮素在核酸和蛋白质合成、光合作用、呼吸作用以及氮代谢等生理生化过程中发挥重要的作用[7,9]。在氮素缺乏时,植株矮小、生长受到抑制,植株总生物量降低,植株会将更多资源分配给根系使得根系具有更强的养分吸收能力,根系生长受到促进,根冠比增加[10]。缺氮会降低光合作用、降低了电子传递速率、光合相关酶的活性和含量,降低羧化作用,增加光能向热能转化[11]。有研究表明,缺氮降低了咖啡幼苗的光合速率(Pn)、电子传递速率(ETR)、PSⅡ原初光能转化效率(Fm/Fv)和PSⅡ实际光化学效率(Fm′/Fv′),且不同咖啡品种对胁迫下敏感度不同[12]。在植物适应生长条件时,细胞器内的活性氧(ROS)水平较低,但在缺氮胁迫下,ROS水平升高,植物细胞内ROS产生和消除间的平衡受到破坏,为了保护膜脂不被氧化破坏,植物体会产生一些抗氧化酶抵御其伤害,抗氧化酶主要有超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD),因此抗氧化酶活性能够反映出植物的抗逆性程度[13-14]。丙二醛(MDA)是膜脂被氧化的最终产物,能够反映脂质膜脂破坏程度[15]。缺氮胁迫与抗氧化系统在很多植物都有研究,如米槁[16]、光皮桦[17]、杨树[18]、连翘[19]等。NUE表示氮素利用效率,表示植物吸收到体内单位质量的氮素能够形成干物质量的多少,Asif等[20]的研究表明不同氮素利用效率的棉花品种在不同硝酸盐浓度下的响应在地上部干质量、根系形态和氮代谢的酶活性等方面存在较大差异。国内开展过黑木相思缺氮胁迫研究[21],但未涉及抗逆性酶、荧光参数以及氮素利用效率和基于各项指标的综合评价。本研究以4个黑木相思优良无性系基因型幼苗(F1、SR3、SR14和SR17)为对象,利用砂培盆栽法人工模拟缺氮胁迫条件,研究缺氮对不同基因型生长和生理生化特性的影响,比较不同基因型之间响应策略的差异,并通过主成分分析结合隶属函数进行耐低氮综合评价,为林木氮素营养研究提供更多的数据支撑,也为林木抗逆性早期选育提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于中国广东省中国林业科学研究院热带林业研究所实验苗圃(23°11′34″N,113°22′40″E),属南亚热带季风气候。根据广州市气象台发布的数据,广州市2022年的年均气温23.2 ℃,7月平均气温为30.5 ℃,年平均降水量为1 891.9 mm,年均日照时数为1 780.9 h。
1.2 试验材料
供试材料为4种黑木相思基因型(F1、SR3、SR14和SR17)组培苗,由中国林业科学研究院热带林业研究所林木生物技术实验室提供。选用1~2 mm的纯净石英砂酸洗晾干后作为基质 ,试验采用规格15.7 cm×12.4 cm×16.5 cm(上口径×底部直径×高)的塑料花盆,每盆装2.6 kg石英砂,每盆种植1株,盆底下放置滤纸防止石英砂流失。营养液采用改良后的Hoagland营养液,设置CK (16 mmol·L-1N素)和-N (0 mmol·L-1N素)两个处理,pH调至5.8,所有药品均为分析纯,采用超纯水配置。
1.3 试验设计
2022年2月中旬将供试基因型从培养瓶中各取出300株种植在土壤中进行炼苗,2个月后选取长势基本一致(苗高为15±0.5 cm)、生长健康的试验材料进行缺氮胁迫试验,试验处理为4个基因型与2种氮素处理的组合,共8个处理,每处理组合30株,每株为1个重复。于2022年5月移栽到装有石英砂的塑料花盆中,缓苗2周,期间加盖80%遮阳网,并用1/4的全素营养液培养,待苗生长平稳并长出新叶后用超纯水培养,饥饿处理1周。2022年6月试验正式开始,前两周用1/2浓度的营养液,第3周后全剂量营养液进行培养。每隔3 d浇100 mL营养液,每天固定时间进行浇水,浇水量可视天气和苗木生长状况进行适当调整。每隔2周配施2‰的多菌灵或甲基托布津与超纯水一起灌入基质中预防霉菌并随机调整摆放位置以减少边缘效应。待12周幼苗出现明显的生长表型差异后,测定表型数据后采集样本进一步测定。
1.4 试验方法
1.4.1 苗高、地径
2022年9月对各处理全部植株进行苗高和地径测量,苗高采用直尺测量,地径采用游标卡尺测量。
1.4.2 生物量及根冠比
将植株分为根、茎和叶三部分,再将各处理植物组织样品放入烘箱中105 ℃下杀青30 min,在70 ℃下烘干至恒质量,然后用天平分别称根、茎、叶的质量,即为各器官干质量。全株总干质量为根、茎和叶干质量之和,根冠比采用根干质量与地上部分干质量(叶干质量+茎干质量)的比值表示。
1.4.3 氮素积累量及利用效率
取测定完生物量的烘干植株,测定各处理下植株根、茎和叶的N含量。N元素含量测定采用凯氏定氮法(详见NY/T 2017—2011);各器官(根、茎、叶)元素N素积累量采用各器官(根、茎、叶)N素含量与各器官(根、茎、叶)生物量的乘积表示;全株N素积累量为根、茎和叶N素积累量之和;全株N素利用效率以全株生物量与全株N素积累量的比值表示。
1.4.4 气体交换参数测定
净光合速率(Pn)、气孔导度(Gs)和胞间CO2浓度(Ci)使用Li-6800便携式光合测定仪(Li-Cor Inc.USA)测定。在2022年9月选择晴天上午9:00—12:00,每个处理选择长势一致的3株,每株选取靠近顶芽倒数第5至第7片成熟功能叶进行测定,并做好标记,共测定9片叶。样品室CO2浓度设置为400 μmol·m-2·s-1,相对湿度设置为50%,流速设定为500 μmol·s-1,风扇转速10 000 rpm,采用LED红蓝光源(90%红光),光强设置为1 000 μmol·m-2·s-1,测定时待Pn趋于稳定后进行记录,每片叶子记录10次,取平均值[22]。
1.4.5 荧光参数测定
选择上述已标记的叶片,使用Li-6800便携式光合测定仪进行测定,测定时间、叶室参数设定与测定气体交换参数的设定相同,饱和脉冲光设为8 000 μmol·m-2·s-1(100 Hz,1 000 ms)。测定前用锡箔纸包裹叶片进行暗适应至少20 min,测定初始荧光(F0)和最大荧光(Fm),随后打开活化光(1 000 μmol·m-2·s-1),光适应30 min以上,测定出光适应下初始荧光(F0′)和最大荧光(Fm′)。计算PSⅡ原初光能转化效率(Fv/Fm)、PSⅡ有效光化学量子产量(Fv′/Fm′)[22]。
1.4.6 叶绿素含量测定
选取上述已标记的叶片带回实验室,将新鲜叶片剪碎混匀,称取0.1 g植物叶片组织和10 mL 80%的丙酮加入15 mL离心管,在低温、黑暗条件下浸泡24 h,期间摇晃2次,待植物组织完全变白后取上层清液测定波长在440、645和663 nm下的吸光度,按照公式计算出叶绿素a、叶绿素b及叶绿素a+b总量的含量[23]。
叶绿素a+b含量(Chla+b)=Chla+Chlb。式中:A440、A663和A645分别为表示在波长440、663和645 nm下测定的吸光度,V为各样品色素提取液的总体积,L;m为各样品的质量,g。
1.4.7 抗氧化酶及MDA测定
SOD活性、POD活性、CAT活性和MDA含量测定参考植物超氧化物歧化酶(SOD)酶联免疫分析试剂盒使用说明书、植物过氧化物酶(POD)酶联免疫分析试剂盒使用说明书、植物过氧化氢酶(CAT)酶联免疫分析试剂盒使用说明书和植物丙二醛(MDA)酶联免疫分析试剂盒使用说明书。
1.5 耐低氮能力综合评价
参考张雷[24]的方法,首先计算各个测定指标的耐低氮系数;然后对各指标的耐低氮系数进行主成分分析,得各综合指标系数及贡献率,求得综合指标值。
式(5)中:CI(m)为综合指标值,Bi为各测定指标进行规范化的值,Prin(m)i为综合指标的系数,之后对各无性系的综合指标进行隶属函数分析;
式(6)中:Xi表示第i个综合指标,Xmin表示第i个指标的最小值,Xmax表示第i个综合指标的最大值。根据综合指标的贡献度进行各综合指标权重计算;
式(7):Wi表示第i个综合指标在所有综合指标中的重要程度,Pi表示第i个综合指标的贡献率,最后结合隶属函数值和权重计算各无性系的综合耐低氮能力;
式(8):D为各无性系在缺氮胁迫下的耐受性综合评价值,D值越大,代表耐低氮能力越强。
1.6 数据处理与分析
本试验所有数据采用Excel 2016软件进行平均数和标准差计算,采用SPSS 22.0软件进行数据处理分析,使用单因素方差分析(One-way ANOVA)和Ducan’s多重比较法检测样本间的显著性差异,N积累量分配率比较采用独立样本T检验,采用Origin 2023软件进行制图。
2 结果与分析
2.1 缺氮对不同黑木相思基因型幼苗苗高和地径的影响
由图1可知,苗高和地径受基因型差异和缺氮处理影响极显著(P<0.01),地径受基因型差异和缺氮处理交互作用影响显著(P<0.05)。F1、SR3、SR14和SR17的苗高在缺氮条件下较对照(CK)均显著降低,降幅分别为20.7%、25.5%、19.1%和13.6%。SR3、SR14和SR17的地径在缺氮下较CK显著降低(P<0.05),降幅分别为17.7%、12.6%和19.2%,而F1变化不显著。

图1 缺氮对不同黑木相思基因型的苗高和地径的影响Fig.1 Effects of nitrogen deficiency on seedling height and ground diameter in different genotypes of A.melanoxylon
2.2 缺氮对不同黑木相思基因型幼苗生物量和根冠比的影响
由图2可知,全株干质量和根冠比受基因型差异和缺氮处理影响显著(P<0.05),根冠比受基因型差异、缺氮处理和两者交互作用影响显著(P<0.05)。与CK相比,4个黑木相思基因型的地上部干质量和全株干质量在缺氮均呈下降趋势。在缺氮下,F1、SR3、SR14和SR17的地上部干质量分别较CK降低了30.9%、43.6%、44.9%和52.7%,全株干质量分别较CK降低了27.3%、40.3%、42.7%和48.4%,缺氮下4个黑木相思基因型的根冠比均显著升高(P<0.05),而根干质量均无显著变化。
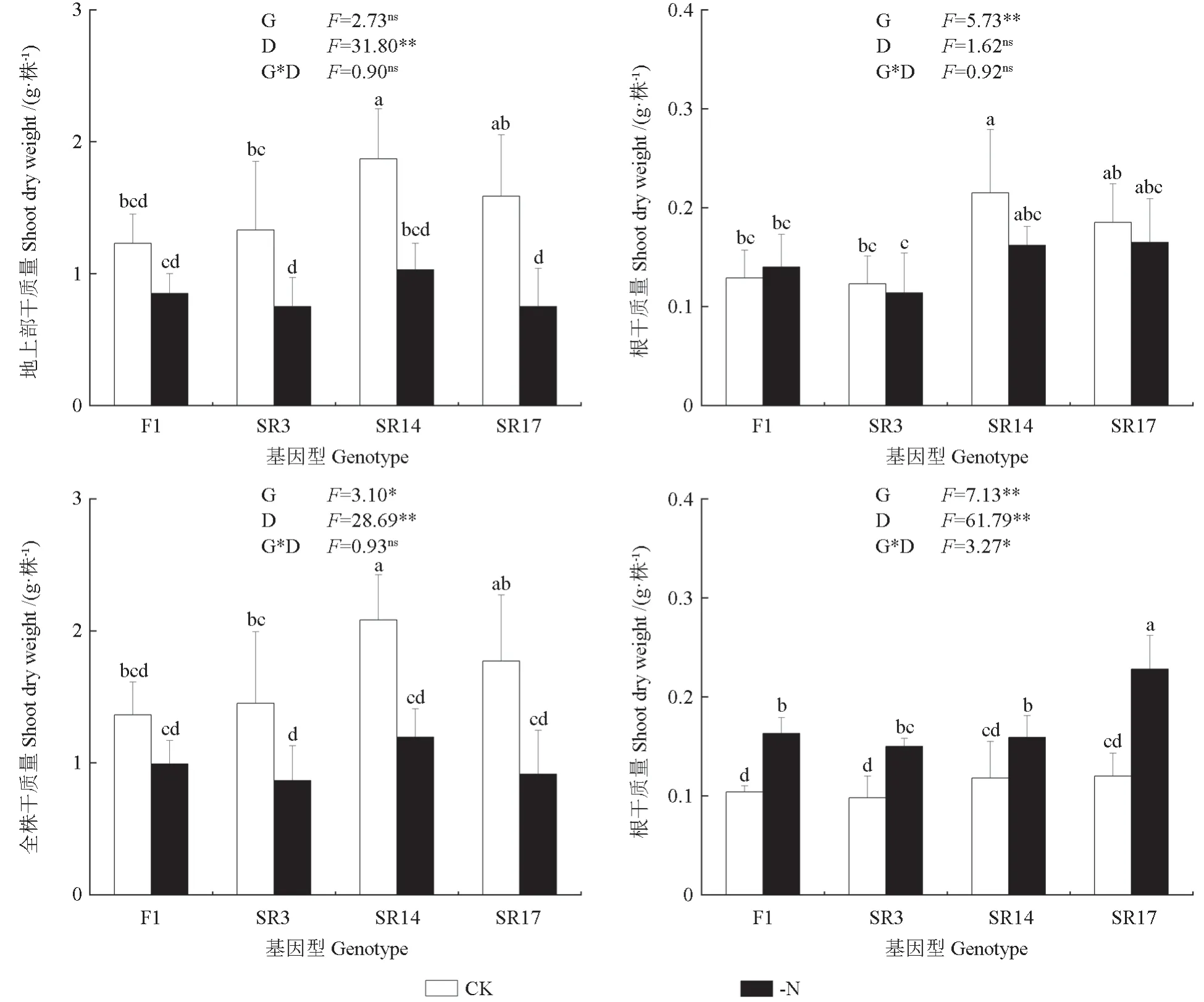
图2 缺氮对不同黑木相思基因型的生物量和根冠比的影响Fig.2 Effects of nitrogen deficiency on the biomass and root-to-shoot ratio in different genotypes of A.melanoxylon
2.3 缺氮对不同黑木相思基因型幼苗光合色素含量的影响
由图3可知,叶绿素a(Chla)含量、叶绿素b(Chlb)含量、类胡萝卜素(Car)含量和叶绿素a+b(Chla+b)含量受基因型差异、缺氮处理以及两者交互作用影响极显著(P<0.01),表明缺氮对不同黑木相思基因型的光合色素含量表现不一。在缺氮下,4个黑木相思基因型幼苗的Chla含量、Chlb含量、Car含量和Chla+b含量均较CK均显著降低(P<0.05),其中这4个指标降幅最大的基因型是SR17,而F1的Chla含量和Car含量降幅最小,SR3的Chlb和Chla+b含量降幅最小。

图3 缺氮对不同黑木相思基因型的光合色素含量的影响Fig.3 Effects of nitrogen deficiency on the content of photosynthetic pigments in different genotypes of A.melanoxylon
2.4 缺氮对不同黑木相思基因型幼苗气体交换参数和叶绿素荧光参数的影响
由表1可知,净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、气孔限制值(LS)和Fv′/Fm′受缺氮处理影响极显著(P<0.01),Fv/Fm、净光合速率(Pn)、胞间CO2(Ci)和气孔限制值(LS)受基因型差异影响显著(P<0.05)。Fv/Fm和F0′/Fm′受基因型和缺氮处理两者交互作用影响显著(P<0.01)。在缺氮下,4个黑木相思基因型幼苗的Pn均显著降低(P<0.05),其中SR17降幅最大,降低了76.8%,其中F1降幅最小,降低了39.1%;缺氮下4个黑木相思基因型幼苗的Gs均呈下降趋势,其中SR17降幅最大,下降了33.3%,F1降幅最小,仅降低了2.8%;在缺氮条件下,4个黑木相思基因型幼苗的Ci较CK均显著升高(P<0.05),而LS均显著下降。在缺氮下,SR17的Fv/Fm显著降低(P<0.05),而对其他基因型无显著影响;F1、SR3和SR17的Fv′/Fm′较CK均显著降低,而对SR14号无显著影响。
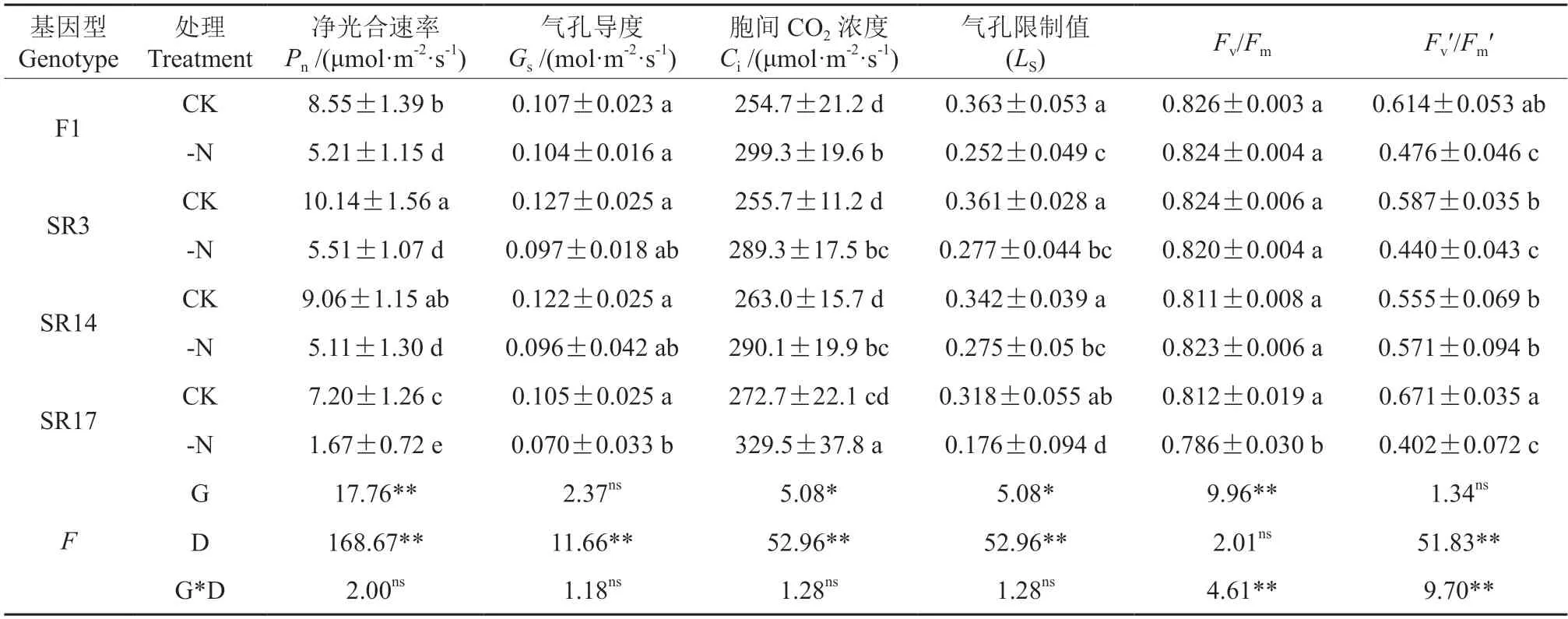
表1 缺氮对不同黑木相思基因型的气体交换参数和荧光参数的影响†Table 1 Effects of nitrogen deficiency on gas exchange parameters and fluorescence parameters in different genotypes of A.melanoxylon
2.5 缺氮对不同黑木相思基因型幼苗氮素积累量和利用效率的影响
由图4可知,全株氮素积累量和全株氮素利用效率受缺氮处理影响极显著(P<0.01),全株氮素积累量受基因型差异影响显著(P<0.05)。在缺氮下,4个黑木相思基因型幼苗的全株氮素积累量较CK均显著降低,其中SR17降幅最大,降低了58.2%,F1降幅最小,降低了39.0%;缺氮下4个黑木相思基因型幼苗的全株氮素利用效率均呈上升趋势,其中SR17增幅最大,增加了23.2%,而SR3增幅最小,仅增加了4.4%。由图5可知,缺氮下,4个基因型黑木相思幼苗的根系氮素积累量占比均呈上升趋势,其中F1、SR3和SR17分别显著增加了53.8%、40%和87.9%。
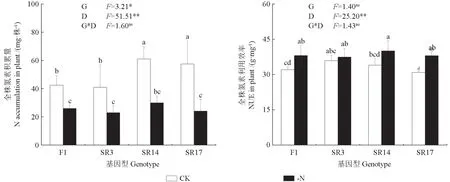
图4 缺氮对不同黑木相思基因型的氮素积累量和利用效率的影响Fig.4 Effects of nitrogen deficiency on nitrogen accumulation and utilization efficiency in different genotypes of A.melanoxylon
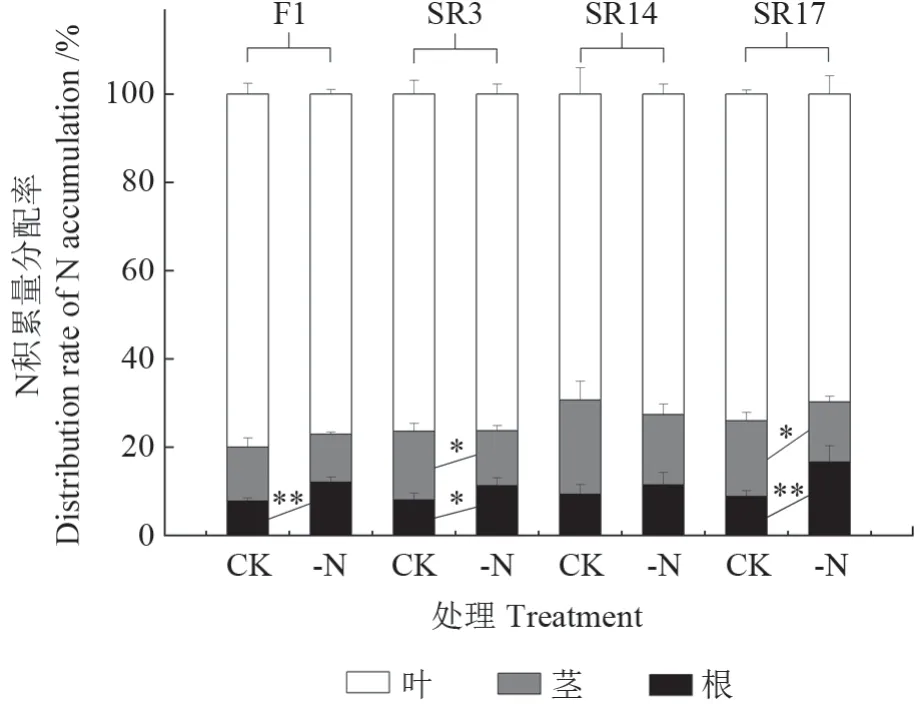
图5 缺氮对不同黑木相思基因型的氮素积累量分配占比的影响Fig.5 Effects of nitrogen deficiency on the distribution of nitrogen accumulation in different genotypes of A.melanoxylon
2.6 缺氮对不同黑木相思基因型幼苗叶片抗氧化酶和MDA的影响
由图6可知,SOD活性、POD活性和CAT活性受基因型差异影响极显著(P<0.01),SOD活性、POD活性、CAT活性和MDA含量受缺氮处理影响极显著(P<0.01),SOD活性、CAT活性和MDA含量受基因型差异进和缺氮处理两者交互作用影响极显著(P<0.01)。在缺氮下,4个黑木相思基因型幼苗叶片的SOD活性、POD活性、CAT活性和MDA含量均显著升高(P<0.05)。

图6 缺氮对不同黑木相思基因型的抗氧化酶和MDA的影响Fig.6 Effects of nitrogen deficiency on antioxidant enzymes and MDA in different genotypes of A.melanoxylon
2.7 缺氮下不同基因型黑木相思幼苗的综合评价
由表2可知,在缺氮条件下选出20个生长和生理生化指标(苗高、地径、地上部干质量、地下干质量、全株干质量、叶绿素a含量、叶绿素b含量、类胡萝卜素含量、叶绿素a+b含量、净光合速率、气孔导度、胞间CO2浓度、Fv/Fm、Fv′/Fm′、MDA含量、SOD活性、POD活性、CAT活性、全株氮素积累量和全株氮素利用效率)作为耐低氮能力评价指标,并对不同基因型耐低氮能力进行分析,计算出各评价指标的耐低氮系数,依据贡献度≥75%的标准选取了2个主成分(综合指标)进行分析。由表3可知,前2个综合指标解释了55.18%和23.78%的贡献度,总贡献度达78.96%,第一个主成分解释了地上干质量、全株干质量、叶绿素a含量、叶绿素a+b含量、类胡萝卜素含量、净光合速率、气孔导度、全株氮素积累量。第二个主成分解释了胞间CO2浓度和全株NUE。由表4可知,通过结合隶属函数算出各基因型的综合评价值并进行排名,评价值越大说明耐低氮能力越强,结果表明耐低氮能力从大到小依次是F1>SR3>SR14>SR17,本研究发现F1耐低氮能力最强。

表2 缺氮下不同黑木相思基因型各指标耐低氮系数†Table 2 Low-nitrogen tolerance coefficients of different A.melanoxylon genotypes under nitrogen deficiency

表3 缺氮下各综合指标的系数及贡献度Table 3 Coefficient and contribution of each comprehensive index under nitrogen deficiency

表4 缺氮下各基因型耐低氮能力综合评价Table 4 Comprehensive evaluation of low nitrogen tolerance of genotypes under nitrogen deficiency
3 讨 论
氮素是植物不可或缺的营养元素,缺氮会对植物生长发育及生产力产生不利影响[25],植物的生长指标能够直观反映植物的生长状况。本研究发现,缺氮条件对4个黑木相思基因型幼苗的苗高、地径、地上部干质量和全株干质量产生了显著的抑制作用,这是由于植物无法吸收足够的氮素以满足自身生长需求[18]。这与黄瓜[26]、槟榔[27]、构树[28]的缺氮处理试验结论相一致。缺氮条件下,F1的根系干质量呈上升趋势,而其他基因型呈下降趋势,同时F1基因型全株积累量在缺氮条件下降幅最小,这是由于F1基因型幼苗应对缺氮胁迫会促进根系生长以吸收更多的矿质营养。研究表明,缺氮胁迫下植物会吸收的氮素更多分配到根系从而使得根冠比增加[29],本试验中缺氮条件下4个基因型幼苗的根冠比均显著增加,根系中氮素积累量占全株比均较CK升高,这是植物应对氮缺乏胁迫的一个适应性策略,这与光皮桦[17]的研究结果相似。缺氮条件下,4个基因型的全株NUE均升高,说明低氮处理能够提高植物的全株NUE,这在前人研究中也得到了证实[30]。
叶绿素是植物光合作用的物质基础,其含量能够反映植物光合能力的强弱[31],叶绿素含量是判定植物光合作用器官是否在逆境胁迫下受到伤害的重要指标[32]。Chla和Chlb在光能的吸收和传递发挥重要作用,Car除具备天线色素功能之外,还可以直接猝灭单线态氧,减少活性氧的产生,以减轻胁迫下植物所受的膜脂过氧化损伤[22]。研究发现在缺氮条件下,4个基因型的Chla、Chlb、Car和Chla+b的含量均显著降低,一方面是由于缺氮条件下叶片氮含量下降,从而抑制了Chla、Chlb和Chla+b的合成;另一方面缺氮下叶片通过蛋白水解酶使得叶绿素分解为氨基酸、酰胺和NH4+以满足其他生理活动[6]。光合参数能够反映一个植物的生长潜力,Pn、Gs、Ci和LS能够反映植物光合生理应对胁迫的情况,是评价光合能力的重要指标[33]。判断Pn下降的主要原因是气孔因素还是非气孔因素要看Ci和LS,如果Ci降低而LS增大,才认为是气孔导度引起,即为气孔因素,如果Ci增加而LS降低,则认为是叶肉细胞羧化能力降低引起,即为非气孔因素[34]。本试验发现缺氮4个基因型幼苗的Pn和Gs均下降,Ci增加,LS降低,说明缺氮下Pn的下降是非气孔因素,由羧化能力降低所致,其原因可能是由于叶绿素含量降低、光合相关酶活降低等[35]。叶绿素荧光参数是反映PSⅡ光化学效率的重要指标[36],对分析植物光能利用具有重要作用,植物正常生长情况下,Fv/Fm的值一般在0.80~0.83范围内[37],仅有SR17基因型幼苗在缺氮条件下Fv/Fm显著降低,并低于0.8,而其他都在0.8以上,说明在缺氮条件下,SR17的黑木相思幼苗PSII中心受损程度较大,这可能是导致SR17的Pn降幅最大的主要原因,而其他基因型不受影响。Fv′/Fm′表明开放时PSⅡ中心原初光能捕获效率,本研究中F1、SR3和SR17均显著降低,说明其PSⅡ反应中心开放程度下降,但SR14无显著差异,这可能是由于SR14本身植物特性及试验环境有关。
植物在受到胁迫时,细胞内活性氧的产生增加,过量的活性氧会使植物的膜脂发生过氧化[38],植物会提高POD、CAT和SOD等抗氧化酶以保护细胞膜脂免受活性氧的伤害。本试验发现在缺氮条件下4个基因型的POD、CAT和SOD活性均显著提高,这是植物应对氮缺乏的一种策略,这与前人在小麦[39]、玉米[40]和青杨[18]的研究结果相似。MDA是植物细胞膜过氧化反应的产物之一,其含量能够反映细胞内膜脂受损程度[41]。本试验中在缺氮条件下4个基因型幼苗的MDA含量均显著升高,尽管抗氧化酶活性维持在一个较高的水平,但在缺氮胁迫下细胞产生的活性氧仍对植物膜脂造成损害。
植物在非生物胁迫下的适应程度受多因素控制,不同评价指标在不同基因型存在差异[42]。本研究采用主成分分析和结合隶属函数分析各个指标并采用耐受系数,第一可以消除无性系之间的固有差异,第二可以消除单一指标异常变化对评价造成的影响,该方法在耐旱性[42]、耐热性[44]、耐盐性[45]等研究中被广泛使用。本试验参考前人研究[17,46-47]并结合指标测定结果,选取了20个具有代表性的耐低氮指标,苗高、地径、地上部干质量、地下干质量、全株干质量、Chla、Chlb、Car、Chla+b、净光合速率、气孔导度、胞间CO2浓度、Fv/Fm、Fv′/Fm′、MDA含量、SOD活性、POD活性、CAT活性、全株氮素积累量和全株氮素利用效率。结果表明,4个无性系的耐低氮能力从大到小依次是F1>SR3>SR14>SR17,F1的耐低氮能力最强,说明缺氮对F1的影响较小,耐胁迫能力强。
本研究没有针对氮浓度做梯度试验,只反映了在缺氮和全氮条件下不同黑木相思基因型的表现差异,且试验是在人工模拟缺素环境下进行的,是否在大田中有相同的结论等待进一步验证。后续将开展不同氮梯度下的试验,并结合代谢组和基因组等手段挖掘耐低氮基因,为黑木相思高效培育提供理论参考。
4 结 论
缺氮胁迫下,4个黑木相思基因型幼苗的生长发育受到抑制,全株氮素积累量显著降低,光合色素含量降低、光合作用受到抑制;缺氮会提高抗氧化相关酶,但过量的活性氧仍给膜脂带来损害;不同基因型黑木相思对缺氮胁迫的响应存在显著差异,通过主成分分析并结合隶属函数综合评价可知,4个基因型的耐低氮能力由大到小依次为F1>SR3>SR14>SR17。F1耐低氮能力最强,表现在胁迫对其影响较小,而SR17耐低氮能力最弱。
