含时滞和Ih流的神经元的同步放电行为*
2023-11-18关利南张新景申建伟
关利南 张新景 申建伟
(华北水利水电大学 数学与统计学院,郑州 450045)
引言
神经元是神经系统最基本的结构和功能单位,神经元相互连接形成复杂的神经元网络,从而在感觉,认知和运动控制中发挥重要作用.研究表明,有节律的行为,如运动、呼吸、进食,以及心跳等是由振荡神经元网络控制的[1].在许多生物中,游泳、进食和心跳等行为背后的节律性活动是由小型神经元网络控制的,这些神经元网络通常通过相互抑制的突触连接,一般包含成对或成群的神经元[2].其中控制水蛭心跳的神经元网络就属于这类小型网络[3].
在水蛭神经元中,除了包括普通的钠、钾离子电流外,还包括一种超极化激活的阳离子电流(hyperpolarization-activated cation current,Ih).由于其特殊的激活特性,自20世纪70年代末在窦房结细胞中首次被发现以来,它便引起了生理学家和生物医学学科研究人员的持续关注[4].到目前为止,在多种类型的神经元和细胞中都观察到了Ih流的存在,如海马锥体神经元、水蛭心脏中间神经元、运动神经元和丘脑神经元等[5-7].研究发现,Ih流在确定神经元的静息电位[8-10],树突整合[11],突触传递[12]等一些基本过程,以及在单神经元和神经元网络的共振和振荡活动中都发挥着重要作用[6].此外,研究还发现运动学习、工作记忆、睡眠和觉醒、空间参考和导航等重要功能都与Ih流相关[13].更重要的是越来越多的研究显示一些疾病,如癫痫、神经性疼痛障碍、心律失常等,也和Ih电流密切相关[14].
同步是自然界中广泛存在的现象.神经系统中的同步行为也受到了广泛关注和研究[15-17].现已发现,在多种神经系统中都存在同步行为,比如视觉皮层,躯体感觉皮层和初级运动皮层[18-20].由于神经活动是神经元的合作过程,因此同步在大脑的信息处理中起着至关重要的作用,例如,处理来自不同感觉系统的信息,形成对外部世界的连贯和统一的感知.但要注意,由于信号传输速度的有限性和神经递质释放的滞后,神经系统中信息的传递通常不是瞬时的,也即在神经网络中时滞普遍存在.此外,研究发现对于抑制性耦合神经元系统,时滞可以诱发其产生多种不同的同步放电模式[21].而神经系统中的群体同步和去同步等复杂放电行为通常与神经系统正常和病态功能密切相关[18, 19].因此神经元网络的同步转迁行为在神经系统至关重要.但Ih流对神经网络特别是抑制耦合神经元网络同步的影响还有待进一步研究.
本文在文献[21]的基础上考虑两个水蛭神经元的相互抑制耦合,构建包含时滞和Ih流的小型网络,用来探究Ih流对含时滞的抑制耦合神经元同步行为的影响.通过数值仿真发现时滞和Ih流都能诱发神经元产生多种不同的同步放电行为.
1 神经元模型和同步指标
1.1 神经元模型
水蛭神经元模型[21]如下,它主要被用来模拟水蛭神经元的节律性活动.
Hi]/τH
其中
f(x,y,z)=1/[1+exp(x(y+z))],
Γ(V)=1/[1+exp(-1000(V-Vsyn))],
i,j=1,2.
模型中参数C代表膜电容,Vi是水蛭神经元的膜电压,hi,mi,Hi分别是钠离子电流的失活门,钾离子电流的激活门和Ih流的激活门的开放概率.gNa,gk,gH,gL分别是钠离子电流,钾离子电流,Ih电流和漏电流的最大电导,ENa,EK,EH,EL是对应离子流的反转电位,τNa,τK2,τH分别是相应电流的弛豫时间,θH是变量Hi的激活电压,Ipol是神经元的极化电流.而gc代表神经元的耦合强度或者说突触电流的最大电导,Esyn和Vsyn分别代表突触电流的反转电位和阈值电压,τ是突触电流的时滞.
模型中的参数取值如下C=0.5nF,gNa=200nS,gK=30nS,gL=8nS,ENa=0.045V,EK=-0.07V,EH=-0.021V,EL=-0.046V,τH=0.1s,τNa=0.0405s,τK=0.9s,θH=0.04V,Ipol=-0.001nA,Esyn=-0.0625V,Vsyn=-0.04V.其中gH,gc,τ为调控参数.
1.2 同步指标
本文用相关系数来表征两神经元的同步情况.两神经元膜电位的互相关系数ρ定义如下:
这里,< >代表N个Vi(i=1,2)的平均值,本文用到的N=1000000.ρ越大代表两神经元的膜电压相关性越高,也即同步程度越高.本文中ρ=1意味着实现了完全同步,而ρ<0意味着负相关.
2 结果
2.1 单神经元的放电特性
单个水蛭神经元模型的放电特性如图1所示,在不考虑Ih流时,神经元表现为周期-6簇放电,如图1(a)所示,也即每个簇内包括6个峰值,此时神经元的放电周期约等于2.9s.当Ih流存在时,比如gH=2nS,单个水蛭神经元模型展示周期-5簇放电,放电周期约为2.1s,如图1(b)所示.也就是说,增加Ih流后,簇放电周期降低,簇内放电个数减少.
2.2 时滞和耦合强度诱导的同步
当两个水蛭神经元相互抑制耦合,在合适的参数下,神经元对会产生同步放电行为,并且随着参数的改变还会出现同步转迁.本小节首先考虑时滞和耦合强度对神经元同步行为的影响,同时本文仅在单神经元的一个簇放电周期内考虑时滞τ的影响.


图2 无Ih流时不同时滞和耦合强度下的同步簇放电模式,黑色和红色线分别代表神经元1和神经元2的膜电压,蓝色和洋红色线分别代表两神经受到的抑制性电流Fig.2 Synchronization bursting patterns with different time delay and coupling strength without Ih current. The black and red curves represent the membrane voltage of neurons 1 and 2, respectively, and the blue and magenta curves represent the inhibitory current received by the two neurons, respectively

当系统中没有Ih流时,两神经元抑制耦合后,随着时滞和耦合强度的变化会产生不同的同步放电行为.例如,当耦合强度gc=1.1nS时,随着时滞τ的增大,神经元依次出现同步的周期-4簇放电,周期-5簇放电和周期-6簇放电,如图2所示.其中图2(a),图2(b)和图2(c)对应的时滞τ分别为0.36s,0.66s,1.3s.图中黑色实线和红色实线分别代表神经元1和神经元2的膜电位,蓝色实线和洋红色实线代表两神经元接收到的抑制性突触电流.此外,当τ=0.36s,gc=1.75nS时,两神经元表现出同步的周期-3簇放电模式.也即随着耦合强度gc和时滞τ的增大,神经元对可以产生多种不同的同步放电模式.
当Ih流存在时,例如gH=2nS,随着时滞τ的增大,神经元对依次产生同步的周期-3簇放电,周期-4簇放电和周期-5簇放电模式,如图3所示,其中图3(a),图3(b)和图3(c)对应的时滞τ分别为0.36s,0.7s,1.0s,三幅图对应的耦合强度都为gc=1.0nS.
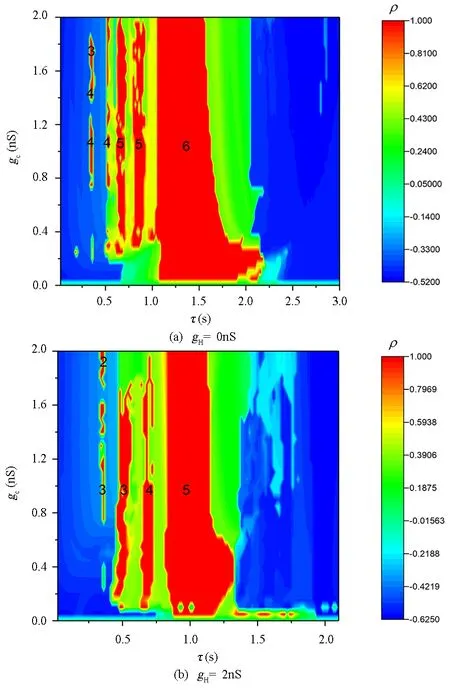
图4 时滞τ和耦合强度gc诱导的同步行为Fig.4 Synchronization behaviors induced by time-delayed τ and coupling strength gc
为了更进一步了解时滞和耦合强度对神经元同步放电行为的调控,我们计算了两耦合神经元膜电压的互相关系数在(τ,gc)双参数平面的分布情况,如图4所示.图4(a)和图4(b)展示的分别是gH=0nS和gH=2nS时的同步情况.其中颜色越红代表同步程度越高,红色代表完全同步.图4(a)和图4(b)都显示随着时滞τ的增大,可以产生多次同步放电.为了更直观的展示不同时滞τ下的同步放电中簇内含有的峰数,我们把不同时滞τ下同步簇放电的簇内峰数用数字标记在图中,数字是几就代表同步的簇内包含几个峰.如图所示,没有Ih流时,随着时滞τ的增大,系统依次产生同步的周期-4簇放电,周期-5簇放电,周期-6簇放电,并且产生同步的周期-6簇放电的τ的取值范围较宽.当Ih流存在时,随着时滞τ的增大,系统依次产生同步的周期-3簇放电,周期-4簇放电,周期-5簇放电,并且产生同步的周期-5簇放电的τ的取值范围较宽.同步簇放电的τ的取值范围与其所处相位的簇放电阈值有关[21].无论有无Ih流,神经元的同步放电都是随时滞τ的增大簇内放电峰数逐渐增大,随着耦合强度的增大,簇内放电峰数减小.但有Ih流与无Ih流相比,簇同步放电的簇内峰数整体降低,且同步的转迁次数变少.
2.3 时滞和Ih诱导的同步
本节,我们进一步考察Ih流对抑制耦合神经元同步放电的影响.与上节类似,我们计算了两耦合神经元膜电压的互相关系数在(τ,gH)双参数平
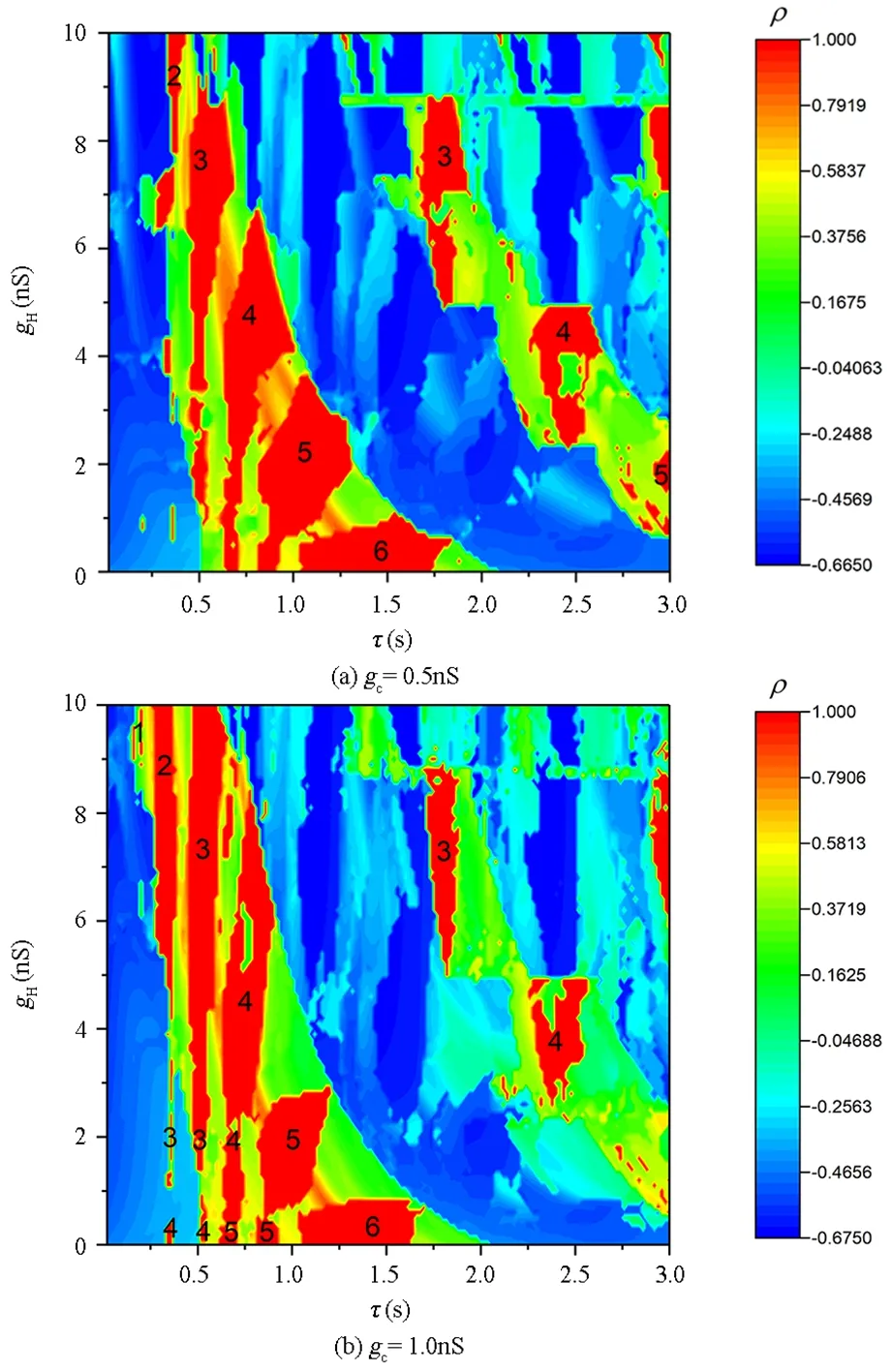
图5 时滞τ和gH诱导的同步行为Fig.5 Synchronization behaviors induced by time-delayed τ and gH
面的分布情况,如图5所示.同样的我们把不同时滞τ和gH下同步簇放电的簇内峰数用数字标记在图中.其中,图5(a)和图5(b)展示的分别是耦合强度gc=0.5nS和gc=1.0nS时的同步情况.从图中可以看出随着时滞τ的减小和gH的增大,神经元的同步放电模式依次为周期-6簇放电,周期-5簇放电,周期-4簇放电,周期-3簇放电,周期-2簇放电,当耦合强度足够大时,还可以转迁为周期-1簇放电行为,如图5(b)所示.即随着时滞τ的减小和gH的增大同步的簇放电模式中簇内峰数逐渐减少,簇放电周期逐渐变短,同时伴随着放电频率的增大.
3 时滞,耦合强度和Ih流诱导同步转迁的动力学原因
从上述结果可以看出,时滞τ,耦合强度gc和Ih流的电导gH都能诱导神经元产生同步行为以及同步转迁.时滞τ和耦合强度gc诱导同步转迁行为的动力学原因可以参阅文献[21],本文不再赘述,下面仅给出Ih流诱导同步转迁的动力学原因.
这里我们借助快慢变量分离方法来解释Ih流诱导的同步转迁行为.图6(a)和图6(b)分别展示了gH=0nS和gH=2nS时单个水蛭神经元模型的快慢变量分离的分岔图.其中,红色实线、黑色虚线和黑色点线分别代表模型快子系统的稳定平衡点,鞍点和不稳定平衡点.绿色实线代表快子系统的膜电压对应的稳定极限环的最大和最小值,黑色实线是全系统的放电轨线.稳定平衡点与鞍点的碰撞点为鞍结分岔(标记为SN),稳定极限环与快子系统鞍点的碰撞点为鞍同宿轨分岔(标记为SH).神经元簇放电的相轨迹大致位于SN和SH点之间.神经元放电期间,在其快子系统的极限环上运行,当相轨迹越过SH点后,脱离极限环的吸引域,进入快子系统的稳定平衡点的吸引域,因此沿着快子系统的稳定平衡点运行,此时处于簇放电的休止期,当越过SN分岔点后又进入快子系统稳定极限环的吸引域从而开始放电,形成下一个簇.
当gH=0nS时,快子系统的鞍结分岔点对应的m约等于0.115063,鞍同宿轨分岔点对应的m约为0.352.当gH=2nS时,快子系统的鞍结分岔点对应的m约等于0.165618,鞍同宿轨分岔点对应的m约为0.352.两种情况下系统的SH点几乎不变,但gH=2nS时的SN点相对于gH=0nS时的SN点右移,如图6所示.也就是说,gH=2nS相对于gH=0nS时,SN与SH点之间对应的m的取值范围缩小,从而导致在SN与SH点之间能产生的峰数减少,近而使得簇放电的簇内峰数降低.为了更直观的观察gH对单神经元簇放电模式的影响,我们计算了单神经元的峰峰间期(ISI)关于gH的分岔图,如图7所示.随着gH的增大,神经元的簇内放电个数逐渐减少.此外,一般情况下,抑制耦合流不易诱发簇内放电个数增多,在抑制流的作用下,神经元簇内放电峰较易被抑制流压制下去,所以当抑制耦合流作用在神经元簇放电的合适相位就会诱发簇内峰数降低的同步簇放电模式.而随着gH的增大,单神经元的快子系统的SN点逐渐右移,从而使得簇内放电个数逐渐减少,最终导致随gH增大产生由簇内峰数较多的同步模式到簇内峰数较少的同步模式的转迁.
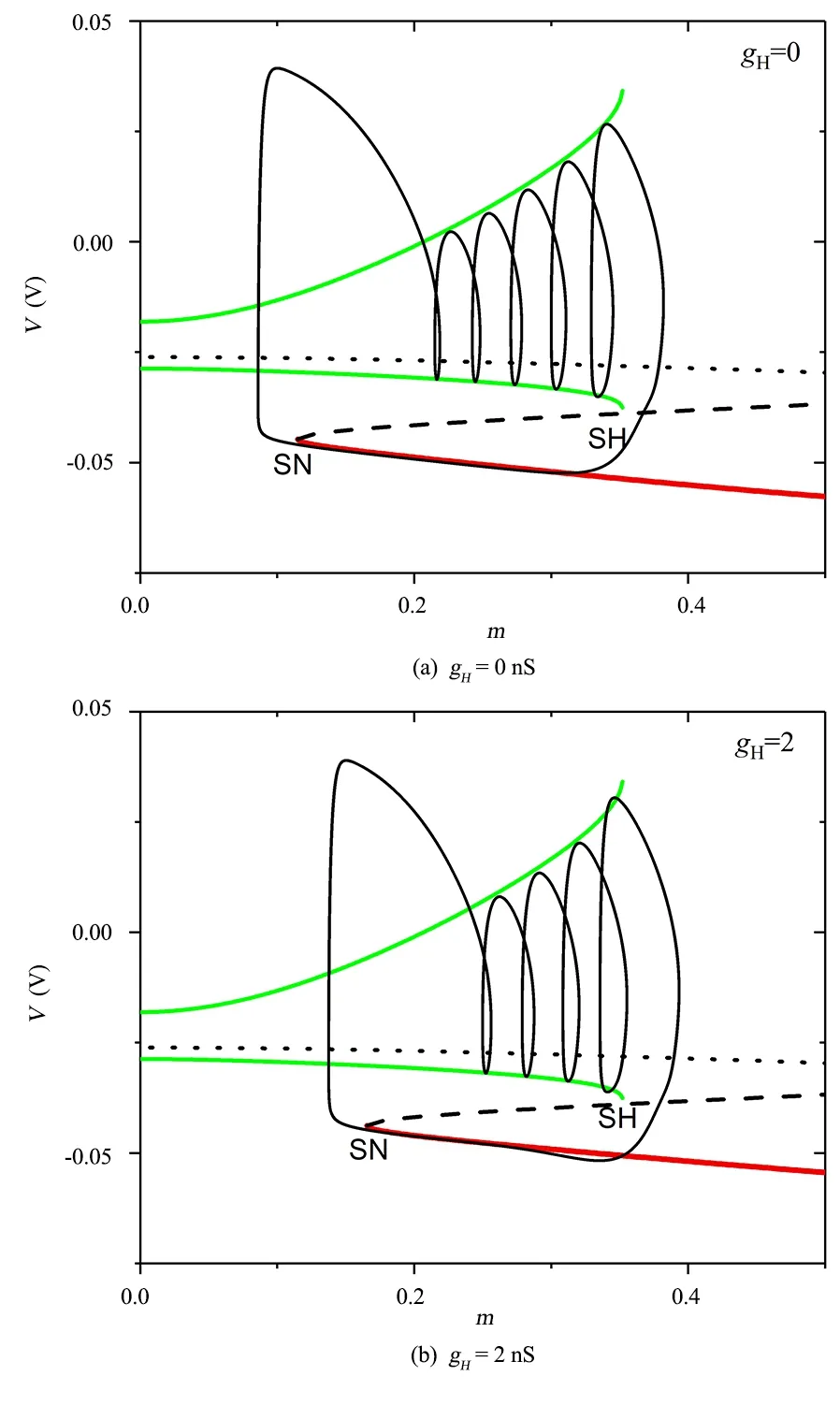
图6 单神经元模型的快慢变量分离图Fig.6 Fast-slow variable dissection diagram of single neuron model

图7 单神经元模型的ISI分岔图Fig.7 ISI bifurcation diagram of single neuron model
4 结论
本文研究了含时滞和Ih流的抑制耦合水蛭神经元系统的同步簇放电活动.通过数值仿真讨论了时滞,耦合强度和Ih流的最大电导对耦合系统同步行为的影响.根据快慢变量分离方法解释了Ih流诱导同步转迁的动力学原因.研究结果表明,无论有无Ih流,在合适的时滞和耦合强度下神经元都可以产生多种同步放电模式,并且无Ih流相对于有Ih流,由时滞诱发的同步放电的簇内峰数相对较多.此外,研究显示gH也能诱发多种同步行为.并且通过分岔分析得到,随着gH的增大,模型的快子系统的鞍结分岔点和鞍同宿轨分岔点之间的参数范围在缩小,近而使得簇内峰数减少,从而使得gH能诱导多种同步放电模式.总之,时滞和Ih流都是影响水蛭神经系统同步转迁行为的重要因素.
