石漠化区香樟不同叶龄光合特性及其主要环境因子的关系分析
2023-11-15刘亚珂苏春花
刘亚珂,姜 霞,苏春花*
(1.贵州民族大学生态环境工程学院, 贵州 贵阳 550025;2.贵州省林业科学研究院, 贵州 贵阳 550005)
化石能源作为一种不可再生资源仍然是我国当前能源消耗的重要抓手,据国家统计局2023年发布《2022年国民经济和社会发展统计公报》显示,我国一次能源生产总量产量巨大,相当于46.6亿t标准煤,增幅为9.2%,原煤产量45.5亿t,增幅为10.5%,原油产量2.05亿t,增幅约为3%,天然气产量2201.1亿kW·h,增长率为6.0%,其次采矿业也增长7.3%。这表明我国对化石能源的依赖仍在上升,也进一步会导致CO2浓度增加。党的二十大报告指出,中国式现代化是人与自然和谐共生的现代化,而低碳化是实现高质量发展的关键环节。国家十四五规划也提出,要加强降低碳排放强度,形成绿色生产生活方式,在2035年,使生态环境根本好转,美丽中国建设目标基本实现。为此,推动绿色低碳发展是建设中国式现代化的题中之义。花瑞祥等[1]研究发现,贵州省经济发展与碳排放呈较为稳定的弱脱钩关系,且能源结构是驱动碳排放脱钩的重要因素。生物质能源是一种低碳、环保、绿色的可再生能源。发展生物质能源不仅可以带来节能降碳的社会效益,而且具有经济效益,而生物质能源树种在山区的存储量巨大,是开发生物质能源的主场地[2]。贵州是世界上面积最大的喀斯特高原区,该地水土流失严重,肥力极差,导致石漠化现象日益严峻,该区生产力逐步降低,使农民与土地矛盾突出,“石漠化贫困”现象严重[3-5],且贵州人地分布不平衡,农民只能种庄稼“靠天吃饭”[6],但据2023年贵州省林业局发布《2022年贵州省林业年鉴》呈现,贵州省现有近1111万hm2森林面积,森林覆盖率达到62.81%,这为发展木本生物质能源树种提供了广阔的空间。
香樟Cinnamomumcamphora(L.)Presl属亚热带常绿阔叶乔木,是主要的生物质能源树种,也是贵州石漠化区生长良好的经济树种。据研究,香樟能提炼多种化学物质,如樟脑烯、柠檬烃、丁香油酚等,而这些物质能够净化有毒空气,起到趋避蚊蝇的作用。香樟适应环境能力强,对土壤要求不高,除寒性土壤外其均可生长[7]。由国家林业局2017年印发《林业生物质能源主要树种目录(第一批)》列出,包括香樟、阴香、天竺桂等在内的生物质能源樟科及其他能源科属树种共计102种,香樟果实含油率为32%,种子含油率在38%~44%,是生物柴油、生物化工的基础材料。当前国内有关香樟的研究报道主要集中于香樟的种子育苗、栽培以及病害威胁、单一环境胁迫下生物特性等方面。如李寿银等[8]对危害香樟的香樟齿喙象进行生长发育与酶活性研究;张泽锟等[9]则对不同梯度Cd胁迫下香樟生理变化进行探究;李小婷等[10]研究夜间单因素对香樟的生理影响。目前,对香樟不同叶龄光合因子与环境因子综合研究鲜有报道。因此,本研究以贵州石漠化区适生树种香樟为对象,分析各因子日变化特性,同时对香樟净光合速率与其他因子间的关系进行相关分析与通径分析,以期揭示香樟能源树种的生理特性,为贵州开发生物质能源树种进行节能降碳提供理论依据。
1 材料与方法
1.1 研究区自然概况
凯里市位于贵州省东南部、东接台江、南抵麻江,地处107°41′13″~108°12′19″E、26°24′13″~26°48′11″N,全市年平均气温在14.1~16.5℃,最热的是7月,平均气温23.9~27.2℃,最冷的是1月,平均气温是2.1~7.9℃。属亚热带温和湿润气候区,是典型的季风气候,植被是中亚热带常绿阔叶林。
试验区位于凯里市凯棠乡石漠化区,该区石漠化状况比较严重:坡度10°~25°,坡向为西南的造林地海拔(550~650 m)区,发育成的砂质土含量高,土层较浅薄,肥力较低[11]。外加人类不合理的活动,如过度开垦、放牧导致该地的植被灌木比较矮小,植株高度为50 cm左右,造成植物存活率低。
1.2 试验材料
选试验地香樟3株,取植株外围上部健康叶片,运用叶片养分速测仪 (TYS-3N)对香樟不同叶龄(6月、8月、10月)的生长指标(氮、叶绿素含量)进行测定,对长势相近的叶子测定3次,结果取均值作为叶片的叶绿素含量和相对氮含量(表1)。此外该叶也作为后期光合生理指标待测叶。

表1 香樟不同叶龄生长情况
1.3 测定方法与内容
1.3.1光合日变化 为反应不同叶龄光合特性,光合日变化试验于2017年晴朗少云的6月、8月和10月进行,选用LI-6400便携式光合测定仪对香樟叶片各项光合生理指标进行测定,其指标测定时间段为8:00~18:00,由于试验地坡度陡,山地多,登山困难等,故每隔2 h测量1次。
测量的指标主要包含环境影响因子:大气温度(Ta)、大气二氧化碳浓度(Ca)、空气相对湿度(RH)、光合有效辐射(PAR),以及光合特性因子:蒸腾速率(Tr)、净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)等,并且计算叶片的瞬时水分利用效率(WUE)、光能利用效率(LUE)及气孔限制值(Ls)。计算相关方法[12-13]:Ls=(Ca-Ci)/Ca;WUE=Pn/Tr;LUE=Pn/PAR。
1.3.2光响应测定 使用仪器红蓝光源,设置叶室温度25℃,Ca为380 μmol·mol-1,RH为70%左右。测定时将PAR由高到低设为2000、1500、1000、500、250、120、60、30、15、0 μmol·m-2·s-1,并以3 min作为时间间隔测其对应Pn变化,获得一系列数据点,将不同叶龄Pn与PAR数据导入光合助手软件(AQ Curve For Photosynthesis)中采用非直角双曲线模型进行拟合得到LSP、LCP、暗呼吸(Rd)等相关光合参数。
1.4 数据分析
运用EXCEL 2010软件、SPSS 23.0软件对采集数据进行相关分析和通径分析等,使用Graphpad prism 9.0软件进行绘图。
2 结果与分析
2.1 主要环境因子的动态变化
影响香樟光合作用的环境因子见图1。试验区午间光照强度较高,6月、10月趋势基本一致,呈单峰曲线,8月上下波动较大(图1a);6月、8月RH在10:00、14:00处差异波动较大(图1b);午前12:00 Ca呈下降趋势(图1c);6月Ta整体呈单峰曲线(图1d)。
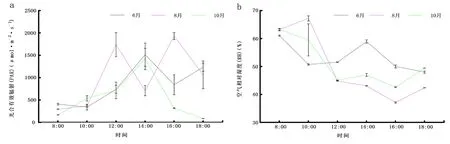
注:误差类型为标准误差,下同。
2.2 香樟不同叶龄光合日变化
2.2.1Pn日变化 由图2可知,香樟不同叶龄Pn均呈单峰型,6月份的Pn日变化有一个高峰在14:00出现,为6.52 μmol·m-2·s-1;而8月、10月峰值在12:00,分别为8.11、8.17 μmol·m-2·s-1,其峰值大小为10月>8月>6月。总体上,净光合速率在上午为递增趋势,在下午为递减趋势。从日均值看,香樟不同叶龄净光合速率表现为8月(6.314 μmol·m-2·s-1)>10月(5.269 μmol·m-2·s-1)>6月(4.415 μmol·m-2·s-1)。
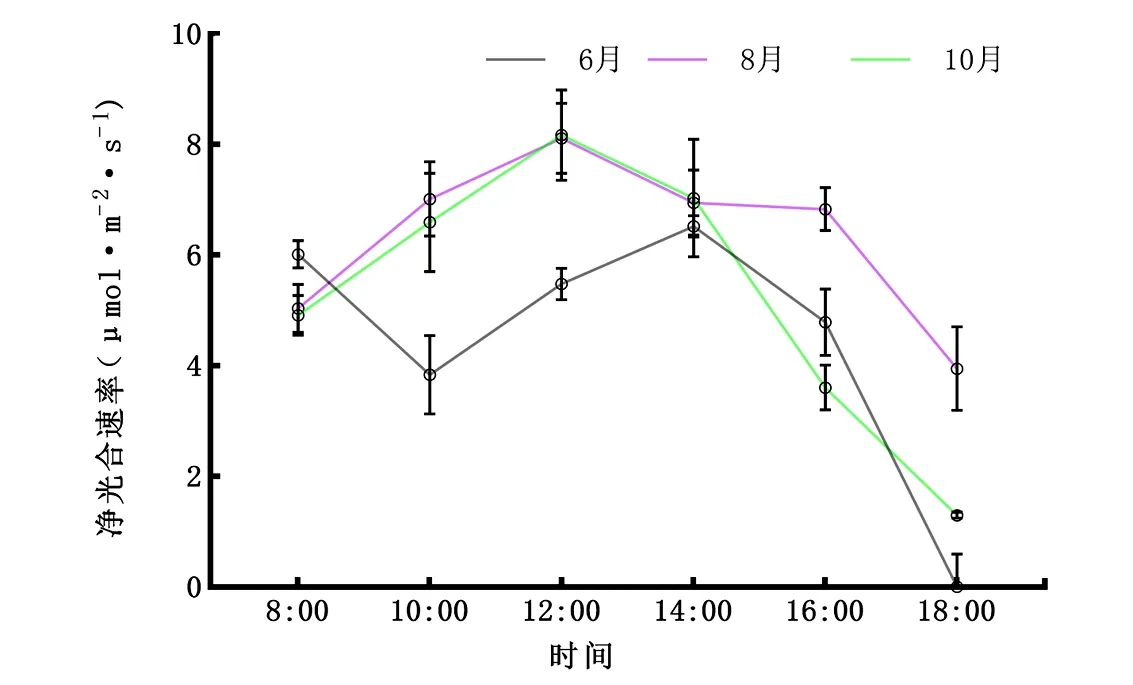
图2 6月、8月、10月净光合速率(Pn)日变化
2.2.2Gs日变化 由图3可知,6月、8月、10月不同叶龄Gs值除早上8:00外,其他时间段均低于0.2 mol·m-2·s-1,且变化趋势也相似,在10:00前下降,而后上升到中午,午后14:00又呈现下降变化,整体展现为先降后升再降的趋势。从日均值看,香樟不同叶龄Gs表现为8月(0.145 mol·m-2·s-1)>10月(0.132 mol·m-2·s-1)>6月(0.105 mol·m-2·s-1)。
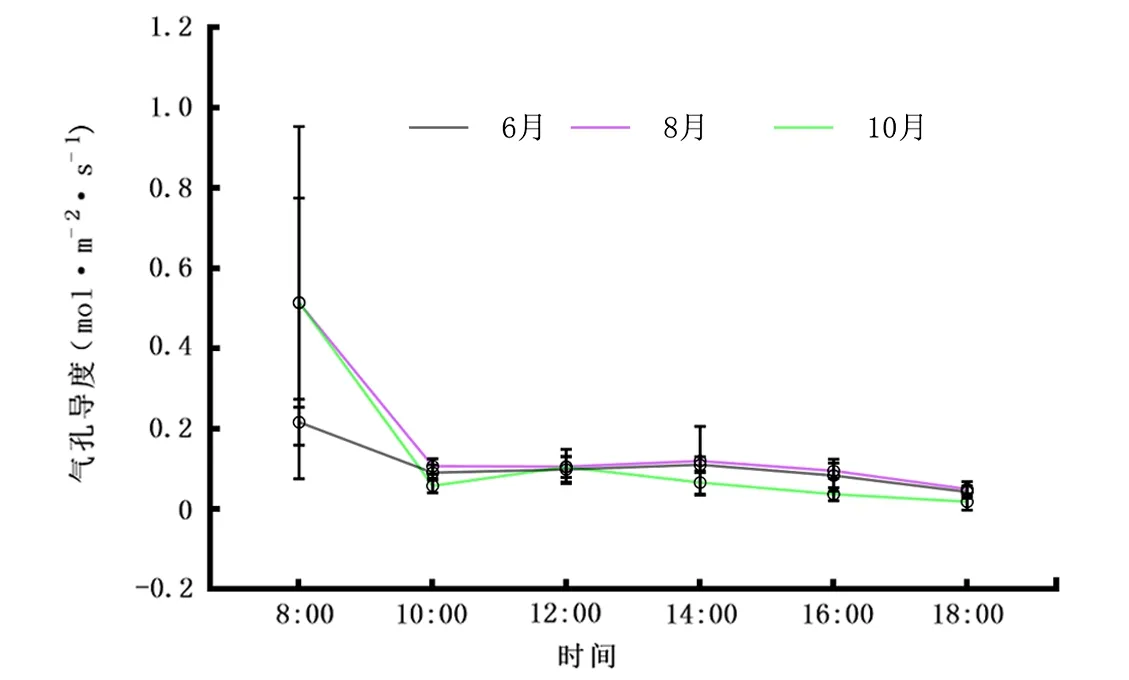
图3 6月、8月、10月气孔导度(Gs)日变化
2.2.3Ci日变化 由图4可知,6月、8月、10月叶片Ci在早上8:00达白天最大值。8:00~14:00,除10月曲线在10:00~12:00有小幅度提升外,其他时间段均与6月、8月一致为递减趋势。总体上Ci日变化呈“V”字型,即早、晚高,午间低。
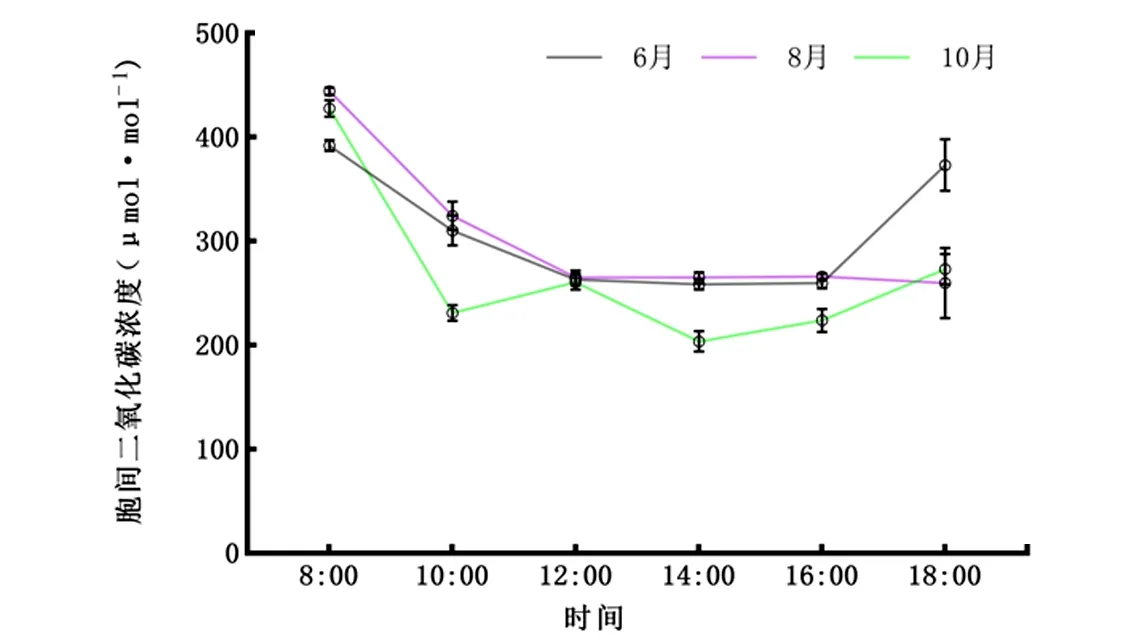
图4 6月、8月、10月胞间二氧化碳浓度(Ci)日变化
2.2.4Tr日变化 从图5可知,6月、10月蒸腾速率Tr整体走势保持一致,均在10:00下降而后上升到12:00,此后一直降低到傍晚,峰值在12:00出现,蒸腾“午休”不明显,但8月出现双峰,蒸腾“午休”较明显。由图1~5可知,从早上10:00开始,香樟Tr、Pn和Gs随着光合有效辐射增强及大气温度的升高而逐渐上升。蒸腾速率与净光合速率日变化趋势相近,说明Tr是影响Pn的重要原因之一。从日均值看,香樟不同叶龄Tr值表现为8月(2.635 μmol·m-2·s-1)>6月(2.082 μmol·m-2·s-1)>10月(1.901 μmol·m-2·s-1)。
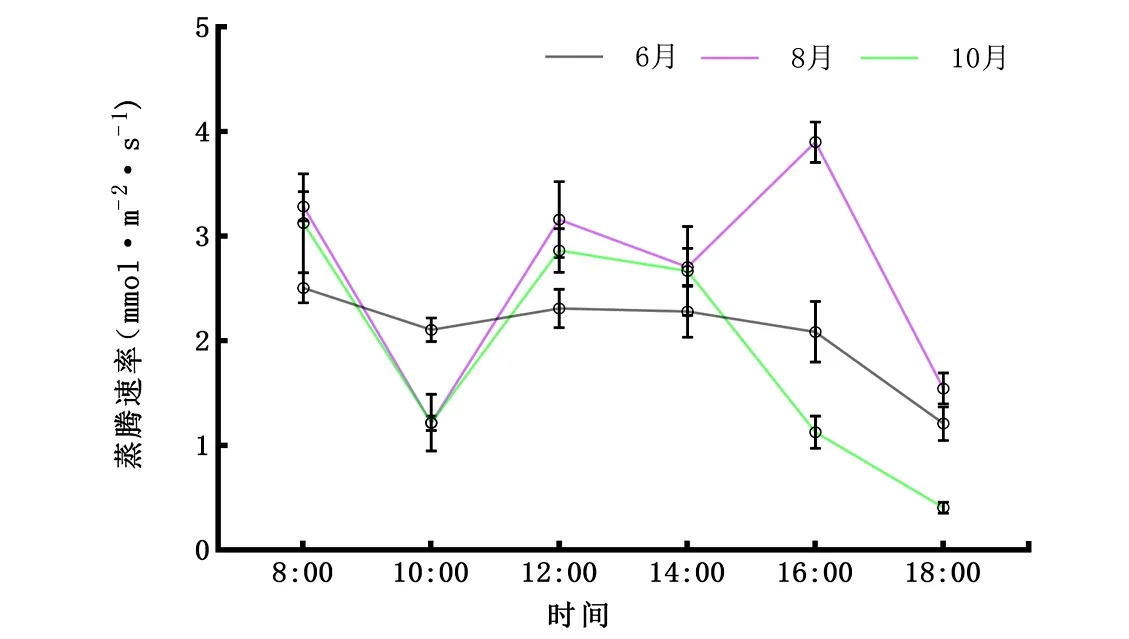
图5 6月、8月、10月蒸腾速率(Tr)日变化
2.2.5WUE日变化 由图6可知,早上8:00,6月叶片WUE为2.44 μmol·mol-1之后开始下降,到10:00为1.85 μmol·mol-1,在10:00到14:00这段时间内其开始上升,随后出现下降情况,而8月和10月WUE曲线变化均为先增后降再增的趋势。从日均值观察,香樟不同叶龄瞬时水分利用率表现为10月(3.273 μmol·mol-1)>8月(2.839 μmol·mol-1)>6月(1.881 μmol·mol-1)。总体上香樟8月WUE值低于10月。此外,香樟不同叶龄WUE均值表现为上午值高于下午值。
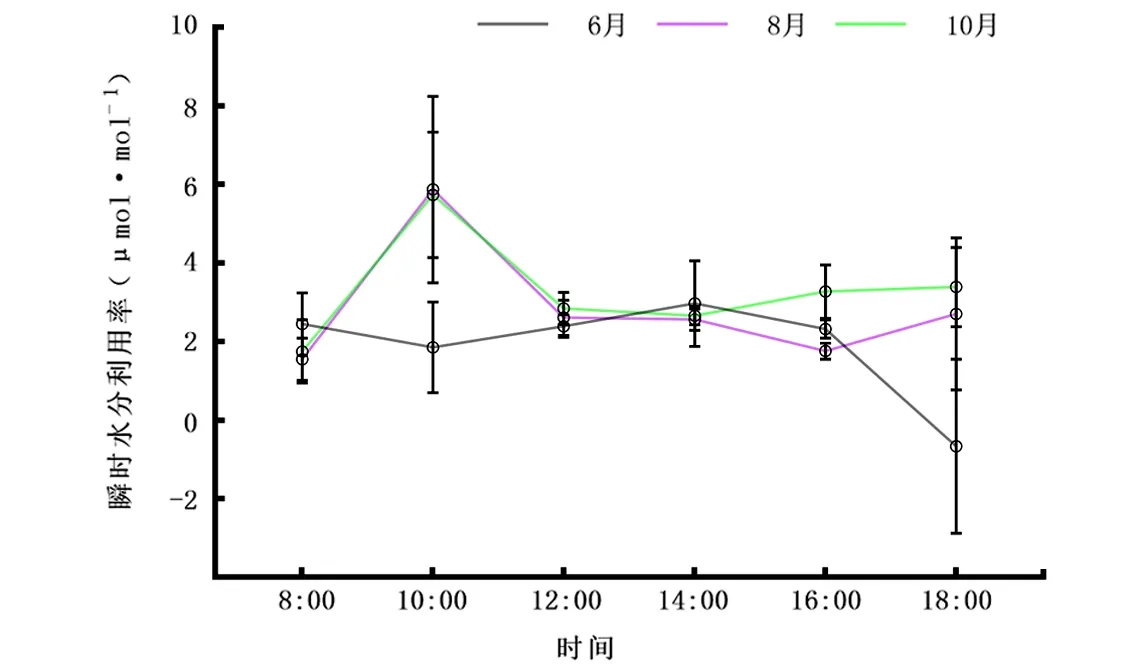
图6 6月、8月、10月瞬时水分利用率(WUE)日变化
2.2.6LUE日变化 由图7可知,6月LUE整体呈下降趋势;从早上8:00开始达最大值0.01 μmol·mol-1;随着大气二氧化碳浓度在8:00~14:00的降低,LUE也在急速下降,到14:00降至0.005 μmol·mol-1;表现出Ca具有影响LUE趋势作用。10月LUE曲线趋势与6月不同,整体表现为每两小时先递减再递增周期性规律;除早上8:00最大值0.03 μmol·mol-1及下午14:00最小值0.005 μmol·mol-1外,其余时间段均稳定在0.01 μmol·mol-1。而8月LUE变化曲线恰好与10月相反,整体呈先递增后递减周期性规律,均在0.01 μmol·mol-1左右浮动。8月、10月光能利用曲线均表现在两小时变化规律上。
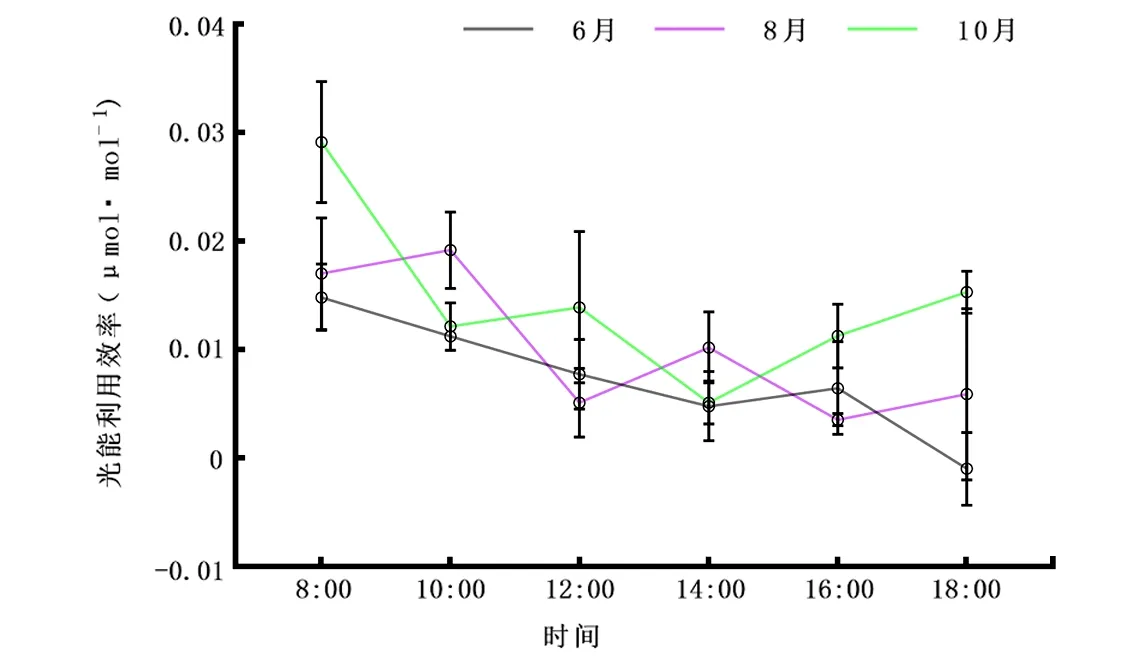
图7 6月、8月、10月光能利用效率(LUE)日变化
2.2.7Ls值日变化 由图8可知,10月气孔限制曲线呈“M”型,在上午10:00出现第1峰,而后下午14:00点出现第2峰。6月、8月气孔限制值整体趋势保持一致,8:00~14:00为上升趋势,6月14:00 Ls达最大值为0.31,随后出现下降趋势;但在18:00同时间段较8月差异大。
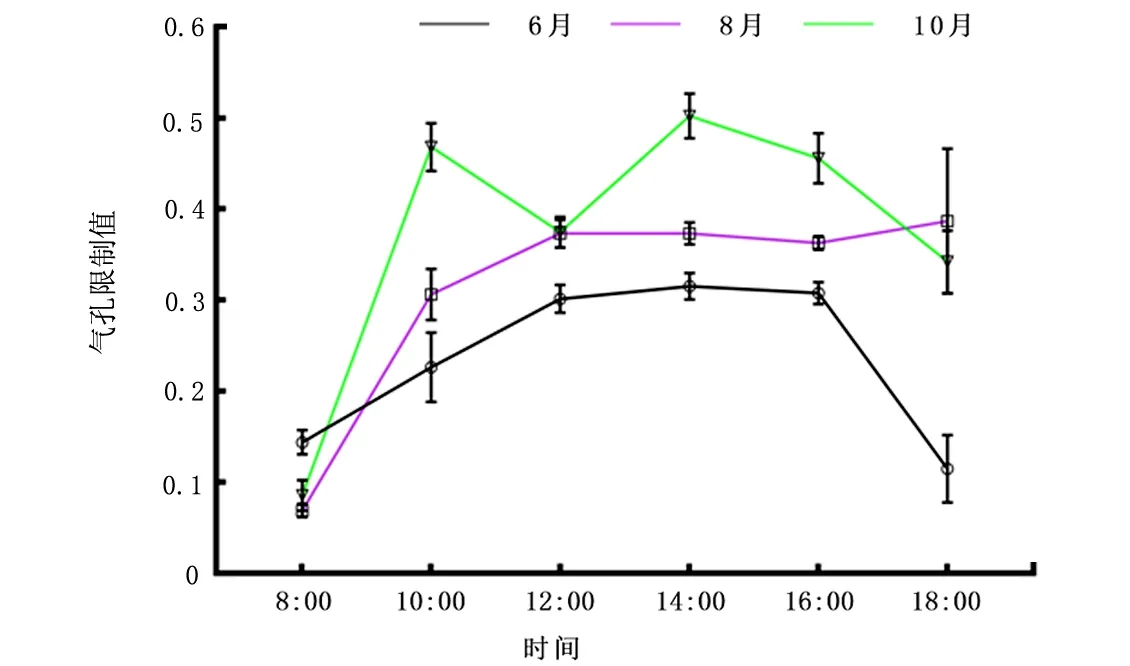
图8 6月、8月、10月气孔限制值(Ls)日变化
2.3 Pn-PAR响应特征动态变化
光响应曲线反映植物Pn随PAR增减而变化的规律。由图9可知,香樟不同叶龄光响应曲线拟合效果较好。当PAR﹤200 μmol·m-2·s-1时,3个月份的Pn随PAR增强呈直线上升,当PAR>500 μmol·m-2·s-1时,Pn上升变得缓慢,且当PAR>1000 μmol·m-2·s-1时Pn逐渐趋于平稳。各曲线对比发现,在相同PAR照射时,叶片净光合速率10月>8月>6月。通过比较不同月份的Pnmax,由表2可知,除6月最大净光合速率为3.95 μmol·m-2·s-1外其他月份Pnmax均在9 μmol·m-2·s-1左右。香樟不同叶龄LSP、LCP从大到小顺序分别为:6月>10月>8月、10月>6月>8月。Pnmax大小顺序为:10月>8月>6月;AQE大小顺序为,8月>10月>6月。
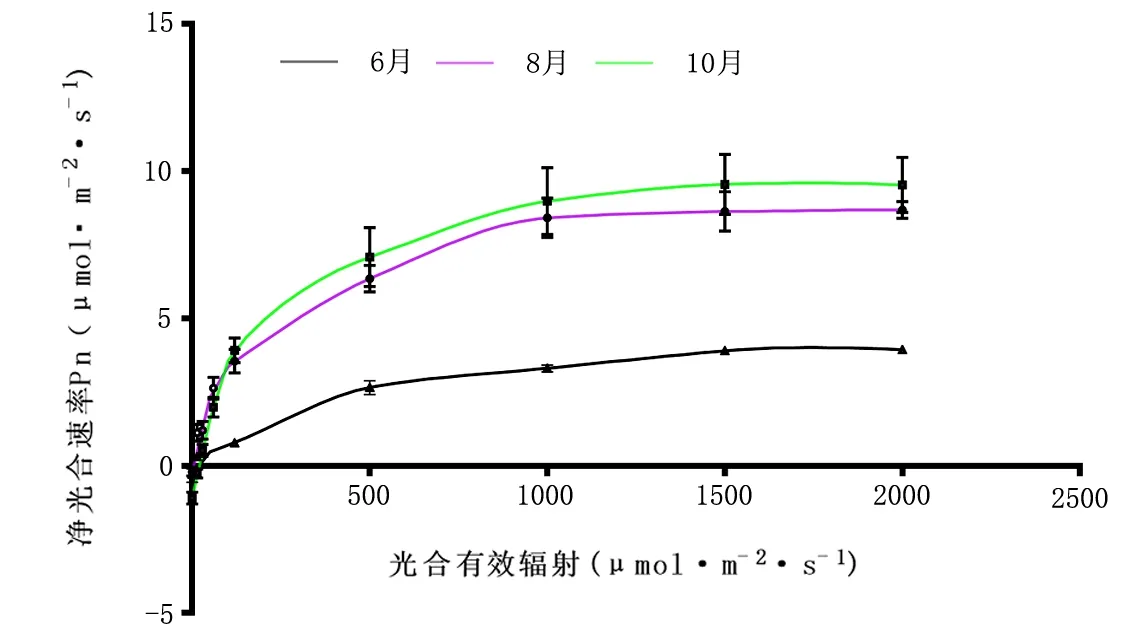
图9 6月、8月、10月光响应曲线变化

表2 香樟不同月份光响应曲线参数
2.4 光合特性因子与环境因子间的相关性分析
对香樟不同叶龄的光合特性因子与环境因子进行简单的相关性分析,见图10~12。
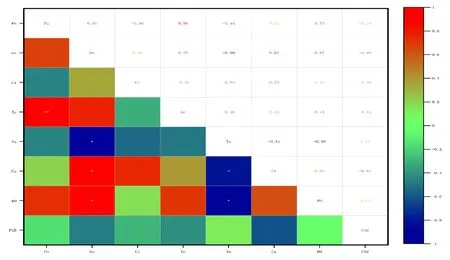
注:*表示在0.05水平上显著性差异,**表示在0.01水平上显著性差异,下同
由图10可知:6月香樟Pn与Tr呈极显著正相关,相关系数为0.96;Pn与Ca、RH、Gs均呈正相关,但在0.05水平上不显著;而Gs与Tr呈显著性正相关,相关系数为0.76,各因子对Pn的影响强弱为:PAR⟨Ca⟨Ta⟨Ci⟨Gs⟨RH⟨Tr。
由图11可知,8月香樟Gs与Ci为极显著正相关,相关系数为0.93,胞间二氧化碳浓度(Ci)和大气二氧化碳浓度(Ca)、大气温度(Ta)、空气相对湿度(RH)均为显著性,其中与Ca呈极显著正相关,相关系数为0.94;与Ta和RH分别表现为显著性负相关、正相关。没发现Pn与Tr的显著性差异,但两者仍然是正相关,相关系数为0.32。各因子对香樟Pn的相关程度大小为:RH⟨Ca⟨Ta⟨Ci⟨GS⟨PAR≈Tr。
由图12知,10月Pn的相关性与8月类似,均表现出对Tr的正相关,但没有显著性差异,此外两月对PAR也是正相关。气孔导度(Gs)与Ci呈极显著性正相关,与Ca呈显著性正相关;同时,Ci与Ca也表现出显著相关性。各因子对香樟Pn的相关程度大小为:RH⟨Ca⟨Gs⟨Ci⟨Ta⟨PAR≈Tr。
2.5 光合特性因子与环境因子对香樟净光合速率的通径分析
相关性分析表明,影响香樟Pn的因子较多,但简单的相关性分析并不能精准的反映各因子间对Pn的关系,为此在相关性基础上采用通径分析方法,更深层次地分析了各因子对Pn的影响(表3)。由通径系数可知,6月,胞间二氧化碳浓度(Ci)对Pn的直接作用为负值(-0.748),大于其他因子的直接通径系数,同时也大于其他因子对Pn的间接通径系数,表明Ci对Pn的直接作用大于通过其他因子的间接作用;RH对香樟Pn的直接通径系数为正数,且大于其他因子的间接通径系数,说明RH对Pn具有较大的增进作用;Gs对Pn的直接作用为负值(-0.066),但在各因子综合影响下,Gs对Pn的相关系数却成为正值,说明光合特性因子和环境因子对Pn作用具有繁杂性。6月决策系数由大到小:R2(Tr)>R2(Ci)>R2(RH)>R2(Ta)>R2(Gs)>R2(Ca)>R2(PAR),且R2(Tr)>0。故Tr为主要决策变量,其原因是它通过Ci和RH对Pn的间接作用分别为0.267和0.113,协助Tr对Pn起促进作用。另外,PAR通过Ci对Pn的间接作用(0.118)抵消其对Pn的直接作用(-0.029),致使其决策系数为正值,表明PAR也是决策变量。
8月,Tr与RH直接通径系数均为较大值(分别是1.037、2.908),表明香樟通过自身对Pn的直接作用要强于通过其他因子的间接作用;Ta和Ca主要通过Ci对Pn发挥作用。按决策系数排序为:R2(RH)>R2(Ci)>R2(Ca)>R2(Tr)>R2(Ta)>R2(PAR)>R2(Gs)。由于PAR通过Ci间接作用(间接通径系数0.781)消除了其对Pn的直接作用(直接通径系数-0.036),使得相关系数发生变化,造成R2(PAR)<0;同理,Gs也是因为通过Ci对Pn的间接作用(-1.102)抵消其对Pn的直接作用(0.040),使其相关系数发生变化,导致R2(Gs)<0,表明PAR、Gs均是限制变量,但非主要限制变量,原因是二者均通过Ci对Pn起限制作用,除此之外多项因子也是主要通过Ci对Pn起限制或促进作用,且总体上Ci决策系数为较大值-0.647,说明Ci是主要限制变量之一。
10月,香樟Tr、Ci的直接通径系数均为较大值,且大于通过其他因子的间接通径系数,表明Tr、Ci对香樟Pn的直接影响要大于其他因子的间接影响。另外,RH是通过Ta和Ca对Pn进行作用的。10月决策系数从大到小为:R2(Gs)>R2(PAR)>R2(Ci)>R2(Tr)>R2(Ta)>R2(Ca)>R2(RH),各决策系数均是负数,说明都起到抑制作用,其中决策系数较强的PAR、Gs表现形式与8月类似,均通过间接作用来抵消直接作用,从而导致相关系数发生变化。
3 讨论与结论
3.1 香樟光合特征与叶龄的相关性
植物叶片光合作用常用来表达其生长和适应各种环境因子胁迫的能力[14-15]。净光合速率(Pn)不仅决定着植物对物质累积能力的高低,而且也决定着植物生长的快慢[16]。通过对贵州省凯里市凯棠乡石漠化区香樟不同叶龄日变化各种参数的测定,发现不同叶龄净光合速率表现为8月>10月>6月,说明8月是香樟生长旺季,净光合速率较大,植物生长能力强,有利于香樟对有机物的积累。对于6月,其净光合速率较低,物质积累少,生长则缓慢。植物光合作用的强弱受自身内部光合因子及外部环境因子的共同影响。在本试验区,虽然8月Tr出现了双峰值,但不同叶龄Pn曲线均呈单峰型,且6月、10月蒸腾速率也是单峰型,光合‘午休’现象不明显,这与冯大兰[17]等对桑树研究一致。说明在6月、10月叶龄下,香樟也不惧高光高温环境。此外,6月上午10:00的净光合速率下降可能是云彩遮阳所致。
叶片Pn、Tr和Gs折线在10:00~14:00变化中趋势基本一致且均达到较大值,而在午后,Pn减小,Gs也在减小这与He等[18]研究类似。表明香樟对午间强光的适应性高于其他时间段。殷亦佳等[19]认为植物通过控制蒸腾作用失水来调节自身稳态,使植物本身失水量最小。本研究WUE、Pn、Tr数值均集中于12:00~14:00,Pn与Tr波动小,导致WUE较稳定。表明蒸腾作用通过调控水分利用,来保持Pn的动态平衡。植物的WUE、LUE是衡量植物生长发育的重要指标之一。WUE由Tr与Pn所决定可表征植物的抗逆性[20-21]。本文中6月WUE出现先降后升再降的趋势,而8月、10月瞬时水分利用率与其相反。这与刘旻霞等[22]研究的“同季节WUE呈先增后减的趋势”不太一致。这可能由于Pn与Tr变化幅度不一样导致,使水分利用率较低。整体上看:8月、10月WUE大于6月,说明在抵抗石漠化胁迫过程中,8月叶吸收水分要强于6月叶,也进一步表明8月香樟对抵抗石漠化胁迫能力较强,这与曹镥镔等[23]干旱胁迫马尾松研究类似。另外,香樟不同叶龄LUE在午间变化幅度小,表明其对光能利用率在午间较稳定。
相关性分析表明,6月,虽然气孔导度对Pn影响能力强,但对其影响最大是Tr,而Ta、PAR影响能力较弱。8月,此时Gs对Pn的影响仍较强,但最强的依然是Tr,不同之处是PAR在该月也表现出对Pn的强相关性。10月影响Pn最强的因子依旧是PAR、Tr,其次是Ta。香樟不同叶龄Tr对Pn均是正相关,且8月、10月Tr、PAR、Ta及Gs对净光合速率的相关性一致,除Gs是负相关外,其他均是正相关。这与冯晶红等[24]对15种消落带适生植物研究结果Pn与Tr、PAR为正相关类似,表明Tr、PAR对Pn的影响较强,但其Gs为正相关与本文不同,很可能是因为本文特殊的喀斯特石漠化区域所致。
3.2 植物叶片Pn-PAR响应曲线参数差异比较
LSP、LCP是反映植物适应强光、弱光的重要指标[25]。香樟不同叶龄的LSP表现为8月<10月<6月,LCP表现为8月<6月<10月。在8月,香樟的LCP较低,反映出香樟在8月对弱光的适应能力强。姜霞等[26]研究发现高饱和点的树种喜光,对强光利用能力强,本文3个月LSP均大于1000 μmol·m-2·s-1表明香樟为喜光植物。AQE是反映植物对光利用效率的指标,香樟不同叶龄AQE表现为8月>10月>6月,以夏季最高,表明其在夏季对光能的利用效率高,此外最大净光合速率Pnmax是8月≈10月>6月,说明夏季的光合作用能力强,也进一步表明,在8月香樟由于高效的光能利用率有利于其对有机物的累积。
3.3 香樟不同叶龄通径分析
根据各因子对净光合速率的通径分析知,6月Tr对香樟Pn的直接通径系数最大;8月Tr与RH直接通径系数均为较大值(分别是1.037、2.908),表明Tr、RH通过本身对Pn的直接影响要大于通过其他因子的间接影响;10月香樟Tr、Ci的直接通径系数也均为较大值,且大于通过其他因子的间接通径系数,说明Tr与Ci对Pn的影响大于通过其他因子的间接影响,这与王冉等[27]对草原植物通径分析类似。此外,整体上香樟不同叶龄Gs与PAR是通过其他因子的间接作用来发挥其对Pn的影响。
综上表明,香樟不同叶龄Pn的变化主要受蒸腾速率、胞间二氧化碳浓度与大气湿度的影响,但影响净光合速率的Tr、Ci和RH不存在简单的线性关系,且还受到其他光合特性因子和环境因子的综合影响。另外,虽然PAR、Gs对Pn表现出简单的强相关性,但二者的通径下表明,PAR与Gs是通过其他因子的间接作用来实现对其Pn的影响。香樟捕捉低光能力较强,能够有效利用林下弱光,有助于其自然生长更新。另外,在8月夏季,香樟的光合能力较强,有利于其对有机物的积累。
