基于线粒体COⅠ基因的三江源草原毛虫金小蜂分子鉴定与系统发育分析
2023-11-11王海贞
王海贞, 刘 昕
1吕梁学院生命科学系, 山西 吕梁 033000; 2中山大学生命科学学院, 广东 广州 510275
青藏高寒牧区草场的高寒草甸是青藏高原地区面积最大的生态系统,是国家生态安全屏障,也是牧民农牧业赖以生存的自然资源。据报道,青藏高寒牧区草场的草原毛虫GynaephoraqinghaiensisChou et Ying危害严重,虫害暴发时每平方米可达上千条,往往牧草刚返青就被啃食殆尽(范小建, 2011)。在草原毛虫危害严重的年份,采用化学农药大面积捕杀成为主要方法,严重破坏了青藏高寒牧区的生态平衡,且加剧了农药残留污染。因此,寻找适合的可对草原毛虫进行生物防治的寄生天敌昆虫,研究草原毛虫的生物防治方法,是实现绿色、环保、无公害、可持续控制草原毛虫虫害的有效途径,对保护青藏高寒牧区草甸生态环境、促进高寒牧区农牧业健康有序发展具有重要意义。
三江源草原毛虫金小蜂PteromalussanjiangyuanicusYang是王海贞(2020)在草原毛虫蛹期采集到的一种寄生蜂,后经过杨忠岐等(2020)从形态学水平鉴定为金小蜂科的一种新种。研究表明,三江源草原毛虫金小蜂对寄主草原毛虫蛹的自然寄生率较高,且出蜂量较大,是草原毛虫蛹期的优势寄生天敌昆虫,可作为扩繁的天敌昆虫对草原毛虫进行生物防治(王海贞,2020)。但是作为金小蜂科的一种新种,分子标记水平的物种鉴定还未完成,以及在进化过程中系统发育关系目前还不清楚,不利于这个新物种分类学及遗传结构的深入研究,制约了其对草原毛虫生物防治潜能的挖掘。
线粒体DNA (mitochondria DNA, mtDNA)是一种群体遗传学研究常用的分子标记(王玲等,2018; 殷玉生等,2012; 张桂芬等,2014; 张莉丽等,2016; Oleksiyetal.,2022),具有进化速度快、母系遗传、基因重组缺失以及易于扩增等特点(Djebbietal.,2018; Palrajuetal.,2018; Wangetal.,2021),是国际公认的广泛应用于昆虫种类鉴别的分子生物学技术之一(Janzenetal.,2017; Zhengetal.,2019)。目前,昆虫线粒体基因研究中结构与功能研究较为深入的是线粒体细胞色素氧化酶亚基Ⅰ(COⅠ)基因,在传粉小蜂Wiebesiapumilae(Hill)(吴文珊等,2013)、榕小蜂Eupristinasp.(陈友铃等,2013)、赤眼蜂Trichogrammasp.(李虹兵等,2018)等多种寄生蜂的遗传多样性及种群结构中广为应用。
本研究基于线粒体COⅠ基因,采用分子生物学手段对三江源草原毛虫金小蜂进行物种辅助鉴定,并通过构建系统发育树,探索三江源草原毛虫金小蜂的系统发育关系,为支持三江源草原毛虫金小蜂的形态学物种鉴定结果,以及深入研究三江源草原毛虫金小蜂的系统发育提供了科学依据,对草原毛虫的生物防治具有重要的意义。
1 材料与方法
1.1 样品采集与保存
本研究所用的三江源草原毛虫金小蜂样品采集自青海省玉树州治多县海拔4580 m高寒草甸的草原毛虫蛹内,并浸泡于非冻型DNA组织保存液(天恩泽)中,4 ℃保存,用于样品总DNA的提取。
1.2 总DNA提取与检测
三江源草原毛虫金小蜂总DNA的提取参照基因组DNA提取试剂盒说明书(Axygen)进行。采用Nanodrop 2000分光光度计 (Thermo Fisher Scientific)检测样品DNA的浓度和纯度。如果提取的样品DNA浓度过高,在进行PCR试验前需对样品DNA进行稀释。
1.3 COⅠ基因扩增反应
三江源草原毛虫金小蜂线粒体COⅠ基因扩增引物设计参照小蜂总科COⅠ基因通用引物(吴文珊等,2013),上游引物序列为5′-CAACATTTATTTTGATTTTTTGG-3′,下游引物为5′-TCCAATGCACTAATCTGCCATATTA-3′。PCR反应的总体积为50 μL,其中含有4 μL DNA模板(40 ng·μL-1),各2 μL双向引物(10 μmol·L-1),5 μL 10×rTaqBuffer (20 mmol·L-1Mg2+),4 μL dNTP Mixture(2.5 mmol·L-1),0.25 μL rTaqDNA 聚合酶(5 U·μL-1),32.75 μL ddH2O;PCR扩增条件为95 ℃预变性3 min, 94 ℃变性45 s,53 ℃退火1 min,72 ℃延伸 1 min,35 个热循环,最后在72 ℃延伸7 min。 扩增产物用1%琼脂糖电泳进行验证,并挑选扩增效果较好的样品,委托华大基因科技有限公司进行正反链双向测序。
1.4 序列分析与系统发育树构建
1.4.1 COⅠ基因序列拼接及碱基组成分析 利用Seqman软件对测序所得的三江源草原毛虫金小蜂线粒体COⅠ基因正反向序列进行拼接,删除头尾载体序列,通过观察测序峰图校正可疑位点,然后将拼接完成后的序列保存为Fasta格式的文件,并对拼接后的基因序列碱基组成进行统计分析。
1.4.2 基于COⅠ基因的物种分子鉴定与构建系统发育树物种选择 本研究将测序得到的COⅠ基因序列在NCBI中Nucleotide BLAST (basic local alignment search tool)进行序列比对,根据比对序列的相似度,进行物种分子鉴定。同时,挑选与三江源草原毛虫金小蜂线粒体COⅠ基因序列相似度较高且具有科、属代表性的膜翅Hymenoptera小蜂总科Chalcidoidea寄生蜂(共5科9属11种)用于遗传关系的分析及系统发育树的构建(表1)。选取的COⅠ基因序列通过登录号从GenBank中下载,并保存为Fasta格式文件,用于物种间基因序列比对和系统发育分析。

表1 GenBank中收录的11种寄生蜂COⅠ基因信息Table 1 Information of COⅠ gene of eleven parasitic wasps from GenBank
1.4.3 系统发育树构建 应用Mega 6.0软件,基于Kimura 2-paramter模型,计算物种间的遗传距离;通过邻接法(neighbor-joining)构建系统发育N-J树,采用自举法(bootstrap)重复抽样1000次检验系统发育树各分支的置信度(Tamuraetal.,2013)。
2 结果与分析
2.1 基因序列分析与物种分子鉴定
获得的三江源草原毛虫金小蜂线粒体COⅠ基因正向序列长度为847 bp,反向序列长度为845 bp。采用Seqman软件对正反向序列进行拼接组装,得到校正后的COⅠ基因序列长度为812 bp,A、T、C、G碱基含量分别为34.36%、42.61%、10.59%、12.44%,AT含量达76.97%,GC含量达23.03%,AT含量显著高于GC含量,具有明显的AT偏好性。NCBI数据库Blast结果显示,三江源草原毛虫金小蜂与蝶蛹金小蜂Pteromaluspuparum(L.) COⅠ基因相似度最高,为91.12%。
2.2 遗传距离分析
遗传距离分析结果如表2所示,三江源草原毛虫金小蜂与蝶蛹金小蜂遗传距离最小,为0.060,与Apocryptasp.遗传距离最大,为2.760,表明进化过程中三江源草原毛虫金小蜂与蝶蛹金小蜂亲缘关系最近,与Apocryptasp.亲缘关系最远。

表2 基于小蜂总科比对物种COⅠ基因遗传距离矩阵Table 2 The matrix of genetic distance for COⅠ gene of comparing Chalcidoidea species
2.3 系统发育分析
系统发育N-J树(图1)显示,Aphelinidae sp.PC124、Aphelinidae sp.、Apocryptasp. PC123这3个物种先后聚为一支,并从小蜂总科其他物种中分离出来,表明这3个物种与其他物种在进化上不属于一个分类阶层;三江源草原毛虫金小蜂先后与Eupelmussp.、Sycoscaptersp.、Idiomacromerussp.、Diaziellabizarre、Eurytomasp.聚为一支,且与Idiomacromerussp.+ (Diaziellabizarre+Eurytomasp.) + (Eupelmussp.+Sycoscaptersp.)形成姊妹群。
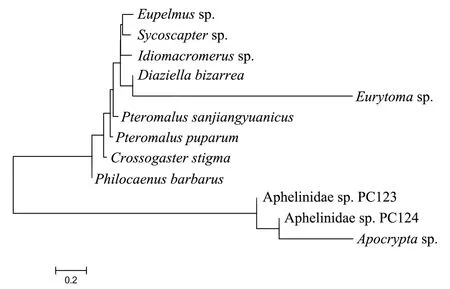
图1 基于COⅠ基因构建的三江源草原毛虫金小蜂系统发育N-J树Fig.1 Phylogenetic N-J tree of P. sanjiangyuanicus based on COⅠ gene
3 讨论与结论
寄生天敌昆虫种类鉴定是害虫生物防控的前提,而建立在形态学特征观察基础上的物种鉴定,通常情况下会因为观察者的主观性而导致物种鉴定不准确,同时,传统的形态学鉴别技术需要由专业的高技术人员识别,而且受样本保存情况、样品发育阶段、细微差异的近缘种等因素的限制(卢西西等,2022)。随着DNA条形码技术在昆虫分类学中的应用,利用形态学观察与分子生物学相结合的技术逐渐取代了单一依靠形态学观察的物种鉴定方法,为物种鉴定提供了更加全面的证据,也为发现隐存种、新物种提供了新的依据(Ogden &Lindsay,2016)。本研究在前期已经对从草原毛虫蛹中采集到的寄生蜂进行了形态学物种鉴定,并认为是金小蜂科的新种——三江源草原毛虫金小蜂(杨忠岐等,2020),但未对其进行分子生物学物种辅助鉴定。为了使鉴定结果更加科学准确,本研究采用COⅠ基因分子标记技术,对采集到的寄生蜂样品再次进行物种辅助鉴定,NCBI数据库Blast结果显示,本研究采集到的寄生蜂与蝶蛹金小蜂COⅠ基因序列相似度最高,而蝶蛹金小蜂为金小蜂科寄生蜂,表明所采样品也可能为金小蜂科寄生蜂,但根据杨忠岐等(2020)的形态学鉴定结果,所采样品与金小蜂科其他寄生蜂种类在形态特征方面存在差异,因此,可以确定本研究从草原毛虫蛹内采集到的寄生蜂为金小蜂科的新种,再次支持了形态学物种鉴定结果。形态学与分子生物学鉴定相结合的方法为三江源草原毛虫分类学的深入研究提供了参考依据,丰富了草原毛虫生物防控寄生天敌资源库。
草原毛虫是分布在我国青藏高原高寒牧区的一种重要害虫,对高寒草甸植被破坏相当严重(王海贞和刘昕,2022)。青藏高原地理环境异常极端恶劣,因此,国内外对草原毛虫寄生天敌资源调查研究报道较少。1980年,沈南英等(1980)在草原毛虫蛹内发现了膜翅目金小蜂科寄生蜂草原毛虫金小蜂PteromalusqinghaiensisLiao。三江源草原毛虫金小蜂与草原毛虫金小蜂虽然在形态上都保留了金小蜂科的特征,但也有一些细微差异,因此被鉴定为2个不同的种(杨忠岐等,2020)。由于草原毛虫金小蜂没有分子方面的基因信息资源,因此无法与三江源草原毛虫做序列比对,进行深入的系统发育研究。故此,本研究只能选用小蜂总科中与三江源草原毛虫金小蜂COⅠ序列相似度较高的11种寄生蜂进行序列比对,以此来研究三江源草原毛虫金小蜂在小蜂总科中的进化地位。遗传距离是评估物种之间亲缘关系的最直接证据,三江源草原毛虫金小蜂与蝶蛹金小蜂的遗传距离最小,揭示在进化过程中三江源草原毛虫金小蜂与蝶蛹金小蜂亲缘关系最近。这在一定程度上再次支持了三江源草原毛虫金小蜂的形态学物种鉴定结果。系统发育树聚类结果显示,三江源草原毛虫金小蜂先后与Eupelmussp.、Sycoscaptersp.、Idiomacromerussp.、Diaziellabizarre、Eurytomasp.聚为一支,且与Idiomacromerussp.+(Diaziellabizarre+Eurytomasp.)+(Eupelmussp.+Sycoscaptersp.)形成姊妹群,表明三江源草原毛虫金小蜂在进化上可能与这5种寄生蜂处于同一分类阶层。
选择合适的分子标记,是用DNA序列进行准确分析的前提,不同的DNA序列在生物体中行使不同的功能,所承受的环境选择压力不同,因而在序列进化速度上存在差异(梁宏伟等,2018; 李爱梅等,2022; 许佳丹等,2019)。对生物不同分类阶元进行鉴定时,对所选择的分子标记的变异速度有不同的要求。在对亚种、种、属等较低的分类阶元进行研究时,要求所选择的分析标记要有较快的变异速度,这样才能确保在亲缘关系较近的不同个体之间存在足够的变异能够将它们区别开来(吴文珊等,2013)。在本研究中,与三江源草原毛虫金小蜂进行序列比对的11个寄生蜂物种均为小蜂总科昆虫,属于较低级的分类单元,因此选用进化速度相对较快的COⅠ序列进行系统发育研究。此外,很多学者认为,仅仅利用一个基因的分子标记进行物种鉴定时,可能存在一定的误差风险(吴文珊等,2013),因此,今后应利用线粒体COⅡ、Cytb基因及核糖体16S、28S基因对三江源草原毛虫金小蜂进行更加全面的分子生物学物种鉴定。
