我国锈色棕榈象遗传多样性及遗传结构分析
2023-11-11肖雨倩黄梦伊
肖雨倩, 黄梦伊 , 阎 伟, 石 娟
1北京林业大学林学院,北京 100089; 2中国热带农业科学院椰子研究所,海南 文昌 571300
锈色棕榈象RhynchophorusferrugineusOliver隶属鞘翅目Coleoptera象虫科Cueculionidae棕榈象属Rhynchophorus(张润志等,2003),别名有红棕象甲、椰子隐喙象、椰子甲虫等(鞠瑞亭等,2006)。该虫在海南、广西、广东等中国南方地区一年可发生1~3代(陈义群等,2011; 欧善生等,2009),世代重叠严重。交配繁殖时,雌虫全天可进行交配,多次交配增加了雌虫排卵数,在一定程度上延长了雌雄成虫的寿命(纪田亮,2018),使得该虫在短时间内可迅速增加种群数量,加剧为害棕榈科植物。利用传统的检测技术并不能在早期发现植株内部的幼虫,导致受害植株很容易被运送到一个没有虫源的地方,直到侵害晚期该虫才被发现(Mankinetal.,2008),这是锈色棕榈象在世界范围内传播速度加快的主要原因之一。锈色棕榈象是南方林业检疫的主要害虫之一。1999年我国口岸检疫部门已有多次截获锈色棕榈象的报道(黄法余等,2000)。20世纪90年代后,棕榈科植物的贸易活动在南方多省份越来越频繁,锈色棕榈象的危害使得福建和广东的棕榈业受到重击。
SSR (simple sequence repeats)标记是以特异引物PCR为基础的分子标记技术,为“中性”标记,能客观反映生物之间的本质差异,很容易进行PCR扩增(Akagietal.,1997; Rassmannetal.,1991),在数量方面没有生物学上的限制,位点具有高度的多态性(Rassmannetal.,1991),呈孟德尔共显性遗传模式。利用PCR扩增出来的不同DNA条带模式可分为不同的基因型,提供了更多的遗传信息(Songetal.,2003)。此外,SSR标记在相关物种中还具有很高的可开发性,这些遗传标记物通常可用作比较遗传作图和进化研究的锚定标记(Varshneyetal.,2005)。基于以上优点,SSR被广泛应用于研究种间与种内遗传关系,在入侵昆虫种群来源鉴定、传播路径、杂交程度等方面的研究逐渐增多(刘佳妮等,2008)。
目前,微卫星分子标记SSR技术在我国各省(自治区)锈色棕榈象不同地理种群间的遗传关系和遗传结构还未有深入研究,仅有王桂花(2014)针对入侵历史较早的福建、海南、广西老疫区作的初步分析,并未涉及到2007年江西和2011年四川的新增疫区。鉴于此,本研究拟运用SSR技术,进一步扩大取样范围,增加近年才入侵的新地理种群(四川、江西)以及海南和云南边境地方的地理种群,对锈色棕榈象在全国范围内的种群进行更详细的遗传结构和分化研究,以期为今后控制锈色棕榈象的传播蔓延提供理论依据。
1 材料与方法
1.1 供试昆虫
2017年5月—2018年5月,在云南、四川、江西、广西、福建、海南等地,使用诱捕器诱捕以及对锈色棕榈象受害树木进行树干解析,共采集到177头供试锈色棕榈象成虫,全部用无水乙醇浸泡保存,并放置在-20 ℃冰箱中备用。
1.2 锈色棕榈象基因组提取
将不同地理种群的锈色棕榈象成虫样本放置在滤纸上5 min,待样本上无水乙醇挥发(无酒精气味)后,取其成虫的头部和前胸背板相连部位的肌肉,置于1.5 mL的微型离心管中,按照AxyPrep基因组DNA小量试剂盒说明书操作步骤,进行提取。
1.3 PCR扩增及产物检测
参考Capdevielle-Dulacetal.(2011)合成13对微卫星引物,选取本研究前期已筛出的9对微卫星,委托北京擎科生物技术有限公司根据引物目的扩增质量和片段大小的不同,在各微卫星引物的上游引物5′端加上FAM、HEX、ROX 3种荧光染料中的一种进行荧光标记,并采用高效液相色谱法纯化合成上游及下游引物(表1)。
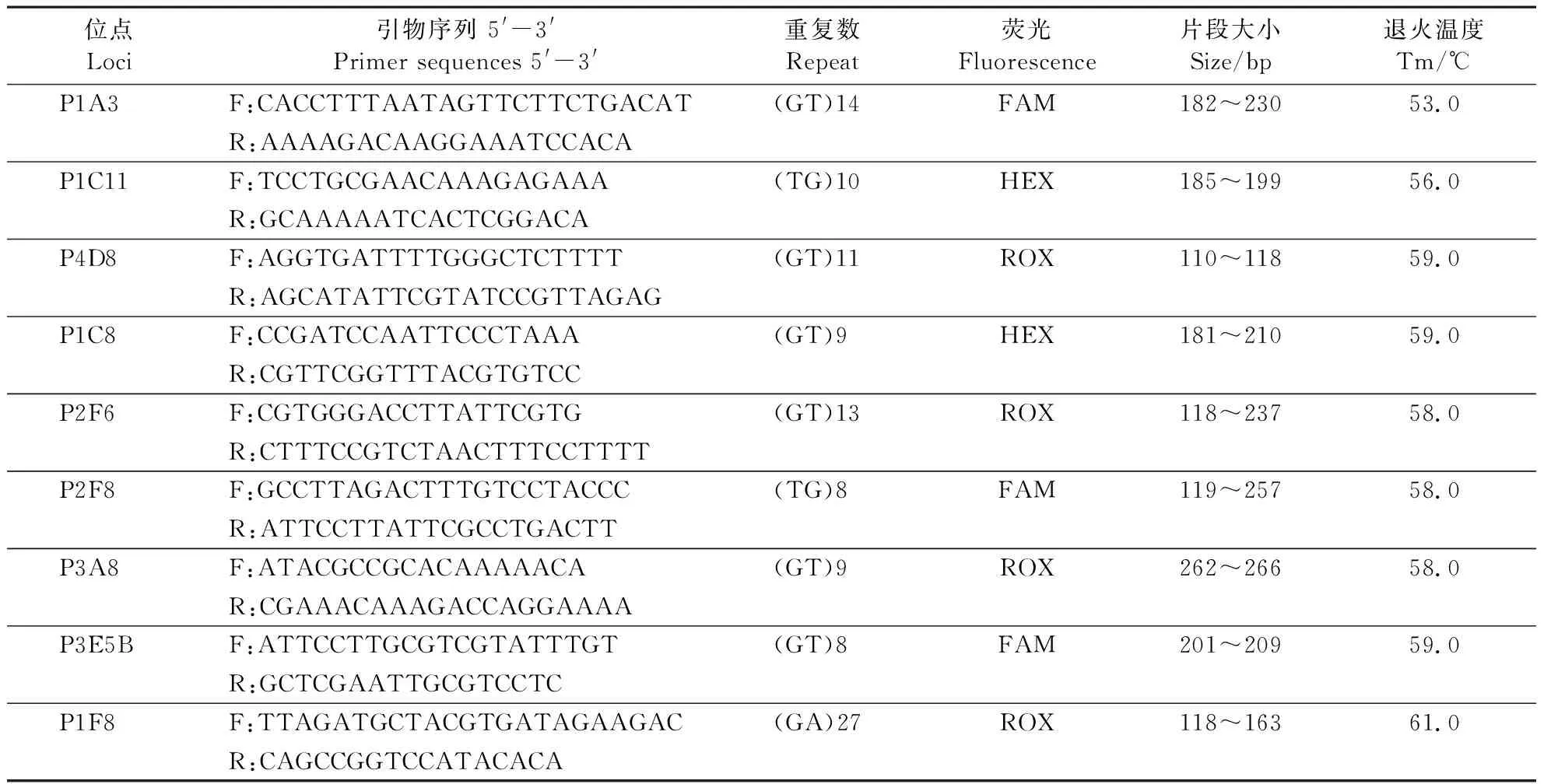
表1 锈色棕榈象9对微卫星引物的序列及相关信息Table 1 Primer sequences and character of 9 microsatellite loci of R. ferrugineus
对每个种群的样本进行微卫星目的片段扩增,进行微卫星荧光引物试验,采用25 μL的体系进行扩增。PCR扩增反应体系为:上下游引物各1 μL,DNA模板1 μL,2×T5 Super PCR Mix (擎科生物公司) 12.5 μL,补充无菌双蒸水至25 μL。反应程序如下:98 ℃预变性2 min;40个循环:98 ℃变性10 s,退火(设置53~61 ℃,具体每个微卫星引物的退火温度见表2)10 s,72 ℃延伸10 s,最后72 ℃延伸5 min,在4 ℃下保存。PCR扩增产物经1.5%的琼脂糖凝胶电泳后,置于凝胶成像仪中检测拍照。检测到有PCR产物后,将扩增相同个体的3个含有不同荧光标记的产物混合在一起,送至北京擎科生物公司,在ABI 3730测序仪上进行毛细管电泳检测。为了更客观地检测不同省(自治区)地理种群间的供试锈色棕榈象是否符合哈迪-温伯格平衡,本研究将12个地理种群以省(自治区)为单位划分为6组进行检测,分别为海南(三亚种群、昌江种群、万宁种群、五指山种群、三亚种群)、云南(蒙自种群、勐腊种群)、广西(玉林种群)、四川(崇州种群)、江西(新余种群)、福建(厦门种群、福清种群)。

表2 中国12个锈色棕榈象地理种群在9个微卫星位点的遗传多样性参数Table 2 The genetic diversity parameters of 12 geographic populations of R. ferrugineus in China
1.4 数据处理与分析
利用Gene Marker software version 2.2.0软件确定每个位点等位基因的大小。读取的数据以Excel表导出保存。利用Convert1.3.1软件将数据转换为后续分析软件需要的格式。
利用PopGen32 (许佳丹等,2019; Yehetal.,2002),对锈色棕榈象各种群SSR数据进行期望杂合度(expected heterozygosity,He)、表观杂合度(observed heterozygosity,Ho)、等位基因数(number of allele,Na)、有效等位基因数(number of effective allele,Ne)、种群的F固定指数(F-statistics)、种群间分化系数(inbreeding coefficient among populations,Fst)、种群内近交系数(inbreeding coefficient in population,Fis)、总近交系数(totalinbreeding coefficient,Fit)、Nei氏标准遗传距离(Nei′s standard genetic distance, GD)(王志存,2008; 郑燕,2014)、基因流(gene flow,Nem)(Cushmanetal.,2006; Kreivietal.,2005)以及哈迪-温伯格平衡(Hardy-Weinberg equilibrium test)(Rodriguezetal.,2009; Wiggintonetal.,2005)的检测分析。
利用BOTTLENECK软件(Piryetal.,1999),基于无限等位基因模型(infinite allele model, IAM)、逐步突变模型(step-wise mutation model, SMM)和双相突变模型(two-phased model of mutation, TPM),使用符号检验(sign test)、标准化差异检验(standardized difference test)和Wilcoxon 检验(Wilcoxon test)3种检验方法对锈色棕榈象的不同地理种群进行瓶颈效应检验(许佳丹等,2019; Kimura &Crow,1964; Ohta &Kimura,1973)。
利用STRUCTURE进行遗传结构分析,对ΔK和lnPr(X|K)的结果进行比较分析,最后得出最佳K值(Evannoetal.,2005; Janesetal.,2017)。
利用GenAlex 6.502 (Rod &Peter,2012)计算种群的遗传距离计算和个体间遗传距离,根据计算结果,选择主坐标分析法(PcoA),查看不同坐标轴解释的种群间和个体间遗传变异所占百分比。
2 结果
2.1 遗传多样性
各个锈色棕榈象地理种群在9个微卫星位点上的观测等位基因数Na的范围为2.7778~5.7778;有效等位基因数Ne为1.8524~3.2160;Shannon′s信息指数I为0.6730~1.3511。各锈色棕榈象地理种群在基因座位上的表观杂合度Ho情况不一样,但均偏低,最高仅为0.4296,最低为0.2500;期望杂合度He在0.4831~0.6644;云南省勐腊锈色棕榈象种群拥有最高的基因多样性指数Nei′s,为0.6422,海南省昌江种群的基因多样性指数Nei′s最低,仅为0.3937 (表2)。
2.2 固定指数及基因流
由表3可知:中国的12个锈色棕榈象地理种群在各位点上Fit为0.1681~0.6952,平均值为0.3778;Fis为0.2717~0.8067,平均值为0.4879。Fst值在位点P1C8最大,为0.3659;在位点P1C11最小,为0.1058,平均值为0.1769。说明锈色棕榈象主要的遗传变异发生在种群内部,种群内发生的遗传变异(82.31%)显著高于在种群间发生的遗传变异(17.69%)。9个微卫星位点中仅有P1C8、P4D8的基因流Nem值小于1,分别为0.4332和0.7919,其他位点的基因流Nem值均大于1。

表3 各位点固定指数及基因流Table 3 Fixation index and gene flow of 9 microsatellite loci
2.3 遗传分化
中国12个锈色棕榈象地理种群两两之间的遗传相似度为0.4183~0.9702 (表4),锈色棕榈象福清种群与来自海南省(昌江、琼海、五指山、万宁、三亚)和云南省(蒙自、勐腊)的种群遗传相似度均小于0.6,种群之间的交流较少。其他种群两两之间的遗传相似度大于0.6,表明这些种群之间基因交流较为频繁。锈色棕榈象三亚种群和福清种群之间的遗传距离最远,为0.8715,万宁种群和五指山种群之间的遗传距离最近,为0.0208。

表4 中国12个锈色棕榈象地理种群间遗传相似系数(右上角)和Nei′s标准遗传距离(左下角)Table 4 Genetic similarity index (above diagonal) and Nei′s standard genetic distance (below diagonal) among 12 geographic populations of R. ferrugineus in China
2.4 哈迪-温伯格平衡
在哈迪-温伯格平衡检测中(表5),锈色棕榈象昌江种群和蒙自种群分别在P1C8位点和P2F8位点上缺失了杂合度。对不同省(自治区)供试种群在9个微卫星位点上的检测发现:四川种群在所有位点上都偏离哈迪-温伯格平衡;海南和福建种群仅分别在位点P2F8和P3A8上符合哈迪-温伯格平衡;云南种群只在P1A3、P1C11上符合哈迪-温伯格平衡;广西种群在P2F6、P2F8、P1C11、P4D8、P3E5B上符合哈迪-温伯格平衡;江西种群在P2F6、P1C11、P4D8、P3A8符合哈迪-温伯格平衡。在这些种群中,广西和江西种群几乎在一半位点上都符合哈迪-温伯格平衡。

表5 不同种群的哈迪-温伯格平衡的P值Table 5 P value of Hardy-Weinberg equilibrium exact test for different populations
2.5 瓶颈效应检测
利用BOTTLENECK软件检验发现(表6),被常用来作检测分析的TPM模型中,云南蒙自的锈色棕榈象种群在3种检测方法中的P值均小于0.05,说明该种群在近期可能经历过瓶颈效应。而其他种群在TPM的3种检测中均未表现出近期有瓶颈效应经历(P>0.05)。IAM模型中3种检测方法得到的结果与TPM模型的结果相似,除了蒙自种群经历过瓶颈效应以外,其他种群都未经历过该效应。在SMM模型中,虽然某些检测显示有一小部分种群经历过瓶颈效应,但大部分种群均没有检测出该效应经历。

表6 检测锈色棕榈象种群瓶颈效应的P值Table 6 P value of BOTTLENECK test on populations of R. ferrugineus
2.6 STRUCTURE的遗传结构分析
对STRUCTURE输出的结果进行ΔK和lnPr(X|K)计算,确定12个锈色棕榈象地理种群聚类的最佳K值为3。从STRUCTURE推断出的三大分支在12个种群中的具体分布情况可知(图1):3个分支在各个地理种群中都有混合现象,说明不同种群间存在不同程度的基因渗透。其中,云南勐腊和云南蒙自种群以及广西玉林种群中第一分支(红色部分)所占的比例较大。在海南的5个种群中,都是第三分支(紫色部分)所占的比例最大,5个种群划分在一组,江西新余和四川崇州种群中第二分支(黄色部分)的占比较大,但是第一和第三分支也有一定的占比,2个种群的分支混合结果较为相似。福建厦门与福建福清种群中第二分支(黄色部分)的占比最高。但是相比福建福清种群,福建厦门种群中第三分支和第一分支的比例要更高一些,与江西新余和四川崇州的种群有一定相似性。
2.7 基于主坐标分析法(PcoA)的分析
利用GenAlex 6.502构建了空间排序二维图,通过二维方式展现不同坐标轴。在种群主坐标(图2)中,第一主坐标解释了总变异的56.71%,第二主坐标解释了总变异的19.33%,由于前2个主坐标解释的总变异大于70%,本研究主要通过第一和第二主坐标分析不同种群和个体间的关系。锈色棕榈象12个地理种群中,海南的5个种群聚为一个类群,这与STRUCTURE分析的聚类结果一致,福清种群与其他所有地理种群都有一定差异,保持最远的距离,与它较接近的种群有厦门和崇州种群,新余种群则与崇州和玉林的种群距离都较相似,但结合种群(图2)和个体(图3)的主坐标分析,新余与崇州距离关系更近,因此,将福清、厦门、崇州、新余的种群划分在一起,STRUCTURE分析结果也显示这3个种群与福清种群有一定的相似性。而云南的2个种群在主坐标分析中也聚在一起,与2个种群关系较近的是玉林种群,将其划为一支。总体来说,主坐标构建的分析结果与STRUCTURE保持一致。
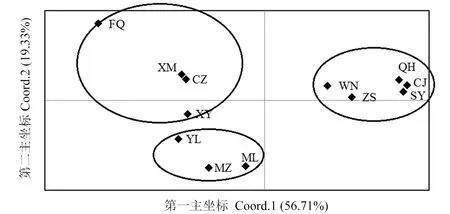
图2 锈色棕榈象12个地理种群的主坐标(PcoA)分析Fig.2 Principal coordinate analysis (PcoA) of R. ferrugineus of 12 geographic populationsCJ:昌江;SY:三亚;QH:琼海;ZS:五指山;WN:万宁;MZ:蒙自;ML:勐腊;CZ:崇州;XY:新余;XM:厦门;YL:玉林;FQ:福清。CJ: Changjiang; SY: Sanya; QH: Qionghai; ZS: Wuzhishan; WN: Wanning; MZ: Mengzi; ML: Mengla; CZ: Chongzhou;XY: Xinyu; XM: Xiamen; YL: Yulin; FQ: Fuqing.

图3 锈色棕榈象不同个体间的主坐标(PcoA)分析Fig.3 Analysis of principal coordinates (PcoA) between different individuals of R. ferrugineusCJ:昌江;SY:三亚;QH:琼海;ZS:五指山;WN:万宁;MZ:蒙自;ML:勐腊;CZ:崇州;XY:新余;XM:厦门;YL:玉林;FQ:福清。CJ: Changjiang; SY: Sanya; QH: Qionghai; ZS: Wuzhishan; WN: Wanning; MZ: Mengzi; ML: Mengla; CZ: Chongzhou;XY: Xinyu; XM: Xiamen; YL: Yulin; FQ: Fuqing.
3 讨论
除了广西和江西的种群在大部分位点符合哈迪-温伯格平衡以外,其他种群在所研究的9个微卫星位点基本都偏离该平衡,尤以入侵较早的海南、福建、云南3个省份的锈色棕榈象偏离最显著。表观杂合度比期望杂合度低是种群偏离哈迪-温伯格平衡的典型表现。观察锈色棕榈象的生活习性可以发现,该害虫在中国多个省(自治区)一年可繁殖2~3代,世代重叠严重。锈色棕榈象各虫期基本都在同一棕榈树中生活取食直至寄主树死亡,由于该害虫迁飞能力较弱,只能短距离扩散,近亲交配繁殖的概率明显大于与异源种群进行交配繁殖,所以在这些入侵历史较早的省(自治区)中,锈色棕榈象之间仍然会存在较高的近交机会。本文得到的较高近交系数Fit(最高达0.6952,平均值0.3778)和总近交系数Fis(最高达0.8067,平均值0.4879)也反映出锈色棕榈象存在一定的近交现象。频繁的近交必然导致种群个体间随机交配的概率下降,从而导致等位基因缺少杂合度,海南昌江和云南蒙自的种群分别在P1C8位点和P2F8位点上缺失杂合度,以及海南、福建、云南这3个省份种群多样性指数中表观杂合度大部分低于期望杂合度的情况也证明了这一点。
阿利效应(Allee effects)指出:当物种种群密度较低时,过分稀疏的种群数量会阻止个体的生长,并且增加了个体找到合适配偶的难度(王瑶等,2007)。入侵物种到达新的入侵地时,一般数量较小,为了避免阿利效应,会先选择一个适合生存的地方进行群聚,停止在新入侵地扩大扩散范围,避免天敌和其他不利因素影响,使种群在新入侵地灭绝(Drake &Lodge,2006; Taylor &Hastings,2005)。待种群密度较高时,再入侵其他地方。四川崇州种群是近年才入侵的种群,在入侵初期,种群可入侵的路径较少,种群数量小。对于新建立的种群,为避免阿利效应,锈色棕榈象会在新入侵地很小范围内繁殖(扩散范围基本没有很明显变化),个体在种群中会因没有合适的配偶而更容易近亲繁殖。新分布地区的限制、环境的适应性以及该害虫在寄主中钻蛀取食的习性,这些因素都可能加重其在新入侵地四川崇州的近交现象,该种群在所有位点的表观杂合度都明显低于预期杂合度的结果也证明了近交现象的存在。因此,对于海南、云南、四川新余、福建的种群出现哈迪-温伯格平衡偏离的原因可以认为主要是种群内频繁近交。
所有检测的种群中只有蒙自种群显示经历了瓶颈效应,但是在对蒙自种群的遗传多样性进行分析时,发现该种群的遗传多样性还是较丰富的,并未因为瓶颈效应而急剧降低种群的遗传多样性。虽然种群有经历瓶颈效应,但是蒙自与一些东南亚国家的地理距离较近,目前,东南亚被认为是锈色棕榈象的起源地之一(Ferry &Gomez,2002),也是发生记录较早的地区,这些国家的锈色棕榈象种群保留着丰富的遗传多样性,且云南省与广西邻近,推断蒙自种群在经历瓶颈效应前,与这些国家和地方之间进行频繁的基因交流,使得其遗传多样性极为丰富。此外,基因流检测结果(蒙自种群与中国其他10个种群有频繁的交流;STRUCTURE中显示该种群有不同分支来源的混合)也都暗示了这点。虽然蒙自种群在近期遭遇了瓶颈效应,但是由于前期拥有极为丰富的遗传多样性基础,即使种群数量减少而丢失了部分遗传多样性,该种群也能保持较高水平的遗传多样性。
Fst值变化越大,表明种群分化程度就越高,锈色棕榈象12个地理种群在各位点的Fst值大部分都小于0.18,平均值为0.1769,说明锈色棕榈象主要的遗传变异发生在种群内部,种群间遗传变异程度低于种群内遗传变异程度。锈色棕榈象种群间的基因流结果显示:大部分种群间存在频繁的基因交流,但福建福清种群与海南和云南的7个地理种群(昌江、琼海、五指山、三亚、万宁、勐腊、蒙自)之间并没有出现高水平的基因流,考虑到福建福清种群与海南和云南个种群的遗传距离较远(Nei′s>0.5),此结果有可能预示着中国锈色棕榈象种群存在多个入侵源和入侵路径。
STRUCTURE和主坐标分析(PcoA)结果结合遗传距离和遗传相似性结果,福清种群与厦门、新余、崇州的种群都有一定的基因交流,这4个种群聚为一支。新入侵地江西新余和四川崇州的种群都表现出与福建福清和厦门种群特征相似的遗传结构。从STRUCTURE结果中发现,虽然福建2个地理种群在这2个新入侵地的遗传结构中占比最大,但来自云南省和海南省的种群也对这2个入侵地有一定程度的基因渗透。所以对于四川崇州和江西新余的种群来说,入侵源不是单一的,主要来自福建。虽然四川省和云南省接壤,与福建省相距较远,但是地理距离不是主要影响因素。推测长期稳定的棕榈植物的贸易往来会导致长时间的入侵,以至于远距离的福建种群的入侵程度大于接壤省份云南省的种群的入侵程度。
STRUCTURE和主坐标分析(PcoA)将云南的2个种群及广西玉林种群聚为一支。云南的勐腊和蒙自种群都表现出与其他种群不同的遗传结构,这2个种群都处于云南省的边境,可能与相邻东南亚国家的锈色棕榈象种群在遗传结构上会更相似,这与Wangetal.(2015)认为中国南部锈色棕榈象最可能的来源地是印度、菲律宾和越南的结论相符。
海南的5个种群单独聚为一支,它们与其他种群的遗传结构有一定区别,这与Wangetal.(2017)对海南和福建种群的遗传结构分析结果相符。海南的地理位置四面环海,气候更为湿润,与其他种群地理位置和气候有着明显区别。此外,Wangetal.(2017)研究显示,来自广东的种群与海南和广西的种群在遗传结构上较相似,并与这2个省(自治区)的种群有很强的基因交流。虽然海南与广东之间有海峡的地理障碍,但是地理障碍不是阻止锈色棕榈象在这些地区分布的主要因素。而人类活动却大大影响害虫的扩散,棕榈植物的运输广泛影响了锈色棕榈象的分布,以此推断海南种群可能从广东通过苗木运输传入。而广西玉林种群在本研究中与云南种群的遗传结构相似,推测广东种群和云南种群在遗传结构上有一定相似性。针对本研究的结果可以推断:中国锈色棕榈象的入侵源并不是单一的,而是由多个来源地在不同时间通过不同或相同的路径传入中国。对锈色棕榈象入侵路径的推断,本文虽然在前人的基础上,增加了更多的供试样品进行了探讨,但是鉴于入侵种入侵方式和路径的复杂性,还需要采集国外如东南亚的种群进行对比分析,才能得到更为精准的结论。
