柑橘大实蝇精巢、精泵和精泵内骨骼的生长发育状况
2023-11-11汤建涛刘文茹桂连友刘梅柯周仁迪华登科姜振宇孙康林张国辉王裕雷蔡开源
汤建涛, 刘文茹, 桂连友*, 刘梅柯, 周仁迪,华登科,姜振宇, 孙康林, 张国辉, 王裕雷, 蔡开源
1长江大学农学院,湖北省农林病虫害预警与调控工程技术研究中心,湖北 荆州 434025;2湖北省农业科学院,农业质量标准与检测技术研究所,湖北 武汉 430064
柑橘大实蝇Bactroceraminax(Enderlein),属双翅目Diptera实蝇科Tephritidae (陈世骧和谢蕴贞,1955),是重要的经济害虫之一(Dongetal.,2014),国外主要分布于不丹、缅甸和印度西北部(Dorjietal.,2006),国内主要分布于湖南、湖北、四川、重庆、广西、贵州、云南等省(自治区、市)(李杖黎等,2012)。柑橘大实蝇寄主主要有柑橘属甜橙、金橘属金橘和枳属枸橘等(赵巍巍等,2020)。雌虫于幼果中产卵,幼虫在果实内部蛀食,在土壤中化蛹,一年发生一代,冬季以蛹越冬(Dongetal.,2013)。受害果实内部局部或全部腐烂并成为糊状,形成蛆柑,果实未熟先黄而脱落,完全丧失食用和经济价值(王小蕾和张润杰,2009)。
不同的环境温度和昆虫日龄都可能对昆虫生长发育产生影响,其中温度是影响昆虫生命活动的重要因素。昆虫作为变温动物,几乎完全无法调控自身体温,温度对昆虫的个体生长、发育、繁殖以及种群的数量和变化地理分布都会产生影响(白月亮等,2022)。温度过高或过低都会严重影响昆虫正常的生长发育和繁殖,极端温度下的昆虫可能会通过形态学、生活史及生理特征的改变来适应环境(陈兵和康乐,2005)。如桃蛀螟Conogethespunctiferalis(Guenée)在不同温度下卵巢发育速度有差异,卵巢发育速率不同,可为短期预测预报提供科学依据(张胜男等,2021);苜蓿斑蚜TherioaphistrifoliiMonell成虫寿命和生殖力在变温处理下比恒温处理明显增加(Bosch,1964)。将某一时间段的温度和种群数量及昆虫的发生规律结合,可以更加准确地预测预报昆虫发生(蔡普默等,2022)。除温度影响昆虫生殖器的大小外,日龄也与昆虫生殖器官的大小存在显著相关性。如亚洲玉米螟Ostriniafurnacalis(Guenée)雄蛾的日龄和精巢之间相关显著(许竟文,2020);蝗科的Chorthippusparallelus(Zetterstedt)在交配时,精巢大小直接影响射精量(Reinhardt,2007)。精巢的体积可以反映精子的数量,飞蝗LocustamigratoriaL.随着精原细胞的成熟、分裂,精子束的体积不断增大,精巢体积也随之增大(郝锡联等,2006)。精巢的大小也体现了雄性性成熟程度,一般来说,在达到性成熟后雄性的内生殖系统状态最佳,之后内生殖系统老化,精巢附腺等产生精液的器官会先开始萎缩(Pitnicketal.,1995),所以了解内生殖器官发育对于害虫发生量、发生期预测具有实践意义。
实蝇科雄性内生殖器结构主要由精巢(1对)、输精管(1对)、生殖附腺(若干对)、精泵(1个)、射精囊(1个)、输入射精管(1个)、输出射精管(1个)和后附腺(1个)组成(Dodson,1978; Drew,1969)。精巢均为一对黄色囊状物,基部具有漏斗状收缩,通过透明、细长的输精管与射精管相连,如泽兰实蝇ProcecidocharesutilisSpreng、昆士兰实蝇Bactroceratryoni(Froggate)、加勒比实蝇Anastrephasuspensa(Loew)、柑橘大实蝇Bactrocera(Tetradacus)minax(Enderleiu)(高熹等,2013; Dodson,1978; Drew,1969)等。精泵体呈淡黄色球体或稍方球体,由射精突演变而成,其体内存在近似扇形的骨片即精泵内骨骼,精泵内骨骼表面着生有强大的肌肉(Dodson,1978; Drew,1969)。欧阳倩等(2014)将瓜实蝇卵巢发育过程分为4期6个级别,刘颖等(2015)将柑橘大实蝇雌虫羽化后的卵巢发育进度分为5个等级,其分级可应用于该害虫预测预报实践中。
目前,有关其精巢、精泵以及精泵内骨骼生长发育状况研究甚少。本文基于光学显微镜测量恒温和室温条件下柑橘大实蝇雄虫的精巢、精泵以及精泵内骨骼的长度和宽度,并建立其长度、宽度和指数的函数模型,比较恒温和室温2种饲养条件下其雄虫器官发育状况差异,明确2种饲养条件下的精巢、精泵以及精泵内骨骼发育趋势及其差异,提供推断其雄虫日龄的方法,将有助于提高柑橘大实蝇人工繁殖效率,为柑橘大实蝇的预测预报及防治提供理论基础。
1 材料与方法
1.1 供试虫源
2020年10月在湖北宜昌长阳县(30°58′N,111°99′E)柑橘园内采集幼虫,置于长江大学昆虫生态学实验室内以橘子饲养。3 d后转移到盛有河沙(厚度5~7 cm)的塑料盆(直径17 cm,高11 cm)内,土壤湿度保持20%左右。
蛹羽化后,连续3 d收集雄成虫(第1天500头,第2天900头,第3天400头),共1800头,置入养虫笼(长×宽×高:35 cm×35 cm×35 cm)中,标注羽化日期,每个养虫笼雄成虫数不超过100头,每天收集雄成虫的一半置于养虫室(长4 m×宽3 m×高3 m),在自然条件下饲养(以下简称室温),另外一半置入同样大小养虫室,在温度(25±2) ℃、相对湿度(65±5)%、自然光条件下饲养(以下简称恒温),雄成虫均饲以酵母菌浸粉和蔗糖(质量比1∶3)混匀的饲料。饲料每3 d更换一次。用喷水壶将清水喷在养虫笼顶部纱网外侧的双层湿润餐巾纸(长20 cm,宽10 cm)上,每天早晚各供一次水。
当天羽化出来的雄成虫记为0日龄,第2天开始收集(1日龄),每天从各养虫室随机收集10头雄虫进行解剖(为防止解剖出现破损,每次挑选12~13头),连续收集35 d共35个日龄的雄成虫,总计700头。
1.2 雄虫生殖器官的解剖和测量
精巢和精泵解剖:用解剖镊子夹住柑橘大实蝇雄虫的胸部并固定,用剪刀剪去头部和胸部,取腹部置于盛有生理盐水的培养皿内,在体视显微镜下,首先将成虫背面朝上,用镊子从断口处夹住两侧体壁,沿腹部末端逐步撕开腹部体壁,然后清除腹腔内脂肪粒和其他组织器官,找到精巢和精泵并分离其周围多余支气管,最后将精巢和精泵置于生理盐水中漂洗2次后供测量和观察使用。
精泵内骨骼解剖:精泵测量完成后,将其置于盛有生理盐水的培养皿内,用解剖镊子分离精泵上的肌肉组织,得到精泵内骨骼供测量和观察使用。
在徕卡体视显微镜(Leica M205A)下观察解剖到的精巢、精泵、精泵内骨骼形态结构,使用自带的软件测量其长度和宽度(图1)。精巢长度测量为沿中间线分割线位置分段测量,然后累加,宽度则测量最宽处(图1A);精泵和精泵内骨骼长度和宽度均是测量其最长处和其最宽处(图1B、C)。各龄期(1~35龄)精巢的长度和宽度、精泵的长度和宽度各测10头(次),各龄期(1~35龄)精泵内骨骼的长度和宽度各测5头(次)。

图1 柑橘大实蝇精巢、精泵、精泵内骨骼的测量位置Fig.1 Measurement sites of testis, ejaculatory pump and ejaculatory pump endoskeleton of male adult, B. minaxA:精巢;B:精泵;C:精泵内骨骼;a:长度;b:宽度。A: Testis; B: Ejaculatory pump; C: Ejaculatory pump endoskeleton; a: Length; b: Width.
1.3 室内的温度和湿度的测量
饲养期间的温度和湿度采用小型气象站(Kestrel 4500, Pocket Weather Tracker, Nielsen-Kellerman, USA)测量。经测定,室内最高平均温度为26.66 ℃,最低平均温度为19.35 ℃,平均温度23.01 ℃,温度范围13.5~34.5 ℃。室内最高平均相对湿度100%,最低平均相对湿度61.34%,相对湿度范围30%~100%。
1.4 数据分析
精巢指数=精巢长度×精巢宽度。精泵指数=精泵长度×精泵宽度。精泵内骨骼指数=精泵内骨骼长度×精泵内骨骼宽度。
所有数据利用DPS软件进行分析(唐启义和冯明光,2002)。在室温和恒温条件下的柑橘大实蝇雄虫的日龄分别与其精巢长度、精巢宽度、精巢指数、精泵长度、精泵宽度、精泵指数、精泵内骨骼长度、精泵内骨骼宽度、精泵内骨骼指数的相关性采用幂函数模型(麦夸特法拟合)分析。相同饲养条件下1~35日龄之间的雄虫的精巢长度、精巢宽度、精巢指数、精泵长度、精泵宽度、精泵指数、精泵内骨骼长度、精泵内骨骼宽度、精泵内骨骼指数的差异比较,采用单因素方差分析(one-way ANOVA),其平均数差异性均采用Tukey法多重比较。恒温和室温饲养条件下柑橘大实蝇雄虫的精巢长度、精巢宽度、精巢指数、精泵长度、精泵宽度、精泵指数、精泵内骨骼长度、精泵内骨骼宽度、精泵内骨骼指数的平均值差异比较,采用配对两处理t检验。
2 结果和分析
2.1 柑橘大实蝇雄虫精巢在成虫期内的生长发育
恒温条件下饲养的雄成虫的日龄(x)分别与其精巢长度(y=2.47547x0.154374,F=506.8,df=1,33,P<0.01)(图2A)、宽度(y=0.678849x0.082041,F=244.4,df=1,33,P<0.01)(图2B)和精巢指数(y= 1.65625x0.24169,F=620.9,df=1,33,P<0.01)(图2C)的相关性均可用幂函数模型(麦夸特法)拟合,他们之间均存在极显著相关。随着雄成虫日龄的增加,其精巢的长度、宽度和指数均呈现逐渐增加趋势。在低日龄雄成虫之间,随着日龄的增加,其精巢长度、宽度、指数呈现逐渐增加趋势,日龄较高后增加趋于平缓。1~35日龄的精巢长度(F=12.3,df=34,315,P<0.01)(图2A)、宽度(F=8.6,df=34,315,P<0.01)(图2B)和指数(F=17.3,df=34,315,P<0.01)(图2C)均存在极显著差异。1日龄时的精巢长度、宽度和指数最低,而精巢长度、宽度和指数最高值分别在33、35和33日龄,与1日龄相比分别增长了70%、35%和125%。
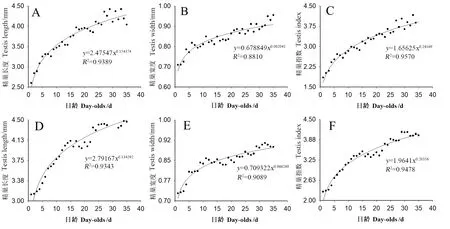
图2 柑橘大实蝇雄虫精巢发育趋势Fig.2 The growth and development trend of testis of male adult, B. minax. A、B和C为恒温饲养;D、E和F为室温饲养。A, B and C were kept in constant temperature rearing; D, E and F were kept in room temperature rearing.
室温条件下饲养的雄成虫的日龄(x)分别与其精巢长度(y=2.79167x0.134292,F=469.5,df=1,33,P<0.01)(图2D)、宽度(y=0.709322x0.066269,F= 329.2,df=1,33,P<0.01)(图2E)和精巢指数(y=1.9641x0.20356,F=642.2,df=1,33,P<0.01)(图2F)的相关性均可用幂函数模型(麦夸特法)拟合,他们之间均存在极显著相关。随着雄成虫日龄的增加,其精巢的长度、宽度和指数均呈现逐渐增加趋势。在低日龄雄成虫之间,随着日龄的增加,其精巢长度、宽度、指数呈现逐渐增加趋势,日龄较高后增加趋于平缓。1~35日龄的精巢长度(F=8.7,df=34,315,P<0.01)(图2D)、宽度(F=5.5,df=34,315,P<0.01)(图2E)和指数(F=12.7,df=34,315,P<0.01)(图2F)均存在极显著差异。1日龄时的精巢长度、宽度、指数最低,而精巢长度、宽度和指数最高值分别在30、32和29日龄,与其1日龄相比分别增长了46.8%、24.7%和81.0%。
室温条件下饲养的雄成虫的精巢长度[(4.00±0.14) mm]显著高于恒温条件下饲养的雄成虫的精巢长度[(3.75±0.13) mm](t=9.97,df=34,P<0.05)。在恒温[(0.85±0.02) mm]和室温[(0.85±0.02) mm]条件下饲养的雄成虫的精巢宽度之间无显著差异(t=0.32,df=34,P>0.05)。室温条件下饲养的雄成虫的精巢指数显著高于恒温条件下饲养的雄成虫的精巢指数(t=7.30,df=34,P<0.05)。
2.2 柑橘大实蝇雄虫精泵在成虫期内的生长发育
恒温条件下饲养的雄成虫的日龄(x)分别与其精巢长度(y=1.1764x0.1325,F=336.8,df=1,33,P<0.01)(图3A)、宽度(y=0.7095x0.206,F=219.3,df=1,33,P<0.01)(图3B)和精泵指数(y=0.8363x0.3381,F=246.1,df=1,33,P<0.01)(图3C)的相关性均可用幂函数模型(麦夸特法)拟合,他们之间均存在极显著相关。随着雄成虫日龄的增加,其精泵的长度、宽度和指数均呈逐渐增加趋势。雄成虫在1~5日龄,随着日龄的增加,精泵长度呈现逐渐增加趋势,在6~35日龄,精泵指数呈现平缓增加趋势。在1~35日龄,精泵长度(F=56.7,df=34,315,P<0.01)(图3A)、宽度(F=49.7,df=34,315,P<0.01)(图3B)和指数(F=60.8,df=34,315,P<0.01)(图3C)均存在极显著差异。在1日龄,精泵长度、宽度、指数最低,而精巢长度、宽度和指数最高值分别在29、33、33日龄,与其1日龄相比分别增长了51.2%、91.8%和186.5%。
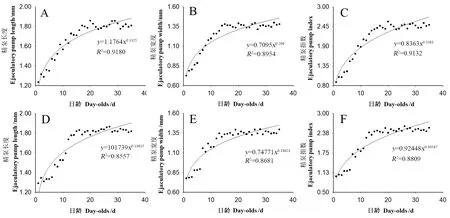
图3 柑橘大实雄虫精泵生长发育趋势Fig.3 The growth and development trend of testis of ejaculatory pump of male adult, B. minaxA、B和C为恒温饲养;D、E和F为室温饲养。A, B and C were kept in constant temperature rearing; D, E and F were kept in room temperature rearing.
室温条件下饲养的成虫的日龄(x)分别与其精泵长度(y=1.1739x0.13615,F=198.3,df=1,33,P<0.01)(图3D)、宽度(y=0.74771x0.18621,F=193.0,df=1,33,P<0.01)(图3E)和精泵指数(y=0.92448x0.30547,F=206.1,df=1,33,P<0.01)(图3F)的相关性均可用幂函数模型(麦夸特法)拟合,他们之间均存在极显著相关。随着雄成虫的日龄的增加,其精泵的长度、宽度和指数均呈现逐渐增加趋势。在1~6日龄,随着日龄的增加,其精泵宽度呈现逐渐增加趋势,在7~35日龄,其精泵宽度呈现平缓增加趋势。在1~35日龄,精泵长度(F=48.5,df=34,315,P<0.01)(图3D)、宽度(F=38.1,df=34,315,P<0.01)(图3E)和指数(F=55.0,df=34,315,P<0.01)(图3F)均存在极显著差异。1日龄时的精泵长度、宽度和指数最低,而精巢长度、宽度和指数最高值分别在27、28和26日龄,与1日龄相比分别增长了43.8%、79.5%和157.4%。
恒温和室温条件下饲养的雄成虫的精泵长度之间[恒温(1.68±0.02) mm,室温(1.69±0.03) mm](t=1.37,df=34,P>0.05)、精泵宽度之间[恒温(1.24±0.03) mm,室温(1.23±0.03) mm](t=0.62,df=34,P>0.05)和精泵指数之间(t=0.55,df=34,P>0.05)均无显著差异。
2.3 柑橘大实蝇雄虫精泵内骨骼在成虫期内的生长发育
恒温条件下饲养的成虫的日龄(x)分别与其精泵内骨骼长度(y=1.1632x0.12227,F=376.3,df=1,33,P<0.01)(图4A)、宽度(y=0.52671x0.26175,F=315.9,df=1,33,P<0.01)(图4B)和精泵内骨骼指数(y=0.65569x0.36180,F=309.7,df=1,33,P<0.01)(图4C)的相关性均可用幂函数模型(麦夸特法)拟合,他们之间均存在极显著相关。随着雄成虫的日龄的增加,其精泵内骨骼的长度、宽度和指数均呈现逐渐增加趋势。在1~5日龄,其精泵内骨骼宽度呈现剧烈逐渐增加趋势,在6~35日龄,其精泵内骨骼宽度呈平缓增加趋势。在1~35日龄,精泵内骨骼长度(F=23.7,df=34,315,P<0.01)(图4A)、宽度(F=27.0,df=34,140,P<0.01)(图4B)和指数(F=30.9,df=34,140,P<0.01)(图4C)均存在极显著差异。在1日龄时的精泵内骨骼长度、宽度和指数最低,而精巢长度、宽度和指数最高分别在28、33和33日龄,与1日龄相比分别增长了52.1%、132.8%和226.5%。
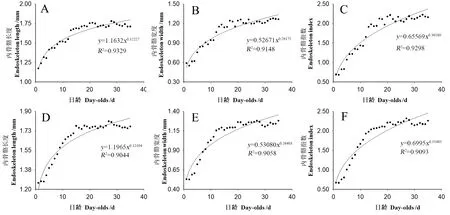
图4 柑橘大实蝇雄虫精泵内骨骼生长发育趋势Fig.4 The growth and development trend of testis of ejaculatory pump endoskeleton of male adult, B. minax A、B和C为恒温饲养;D、E和F为室温饲养。A, B and C were kept in constant temperature rearing; D, E and F were kept in room temperature rearing.
在室温条件下饲养,成虫的日龄(x)分别与其精泵内骨骼长度(y=1.1965x0.12104,F=289.0,df=1,33,P<0.01)(图4D)、宽度(y=0.53080x0.26403,F=253.1171,df=1,33,P<0.01)(图4E)和精泵内骨骼指数(y=0.69595x0.35483,F=244.7,df=1,33,P<0.01)(图4F)的相关性均可用幂函数模型(麦夸特法)拟合,他们之间均存在极显著相关。随着雄成虫的日龄的增加,其精泵内骨骼的长度、宽度和指数均呈现逐渐增加趋势。在1~6日龄,其精泵内骨骼宽度呈现剧烈逐渐增加趋势,在7~35日龄,其精泵内骨骼宽度呈现平缓逐渐增加趋势。1~35日龄的精泵内骨骼长度(F=29.3,df=34,140,P<0.01)(图4D)、宽度(F=28.0,df=34,140,P<0.01)(图4E)和指数(F=33.1,df=34,140,P<0.01)(图4F)均存在极显著性差异。1日龄时的精泵内骨骼长度最短,27日龄时精泵内骨骼长度最大,与其1日龄相比增长了43.7%。2日龄时的雄成虫精泵内骨骼宽度,指数最低,而最高值分别在35和27日龄,与其1日龄相比分别增长了141.5%和246.3%。
室温条件下饲养的柑橘大实蝇雄成虫的精泵内骨骼长度[(1.65±0.03) mm]显著高于恒温条件下饲养的雄成虫的精泵内骨骼长度[(1.61±0.03) mm](t=6.29,df=34,P<0.05)。恒温[(1.07±0.04) mm]和室温[(1.09±0.04) mm]条件下饲养的雄成虫的精泵内骨骼宽度之间无显著差异(t=1.82,df=34,P>0.05)。室温条件下饲养的雄成虫的精泵内骨骼指数(1.84±0.08)显著高于恒温条件下饲养的雄成虫的精泵内骨骼指数(1.77±0.08)(t=3.98,df=34,P<0.05)。
3 讨论
高熹等(2013)的研究表明,泽兰实蝇雄成虫的精巢长度和宽度在羽化初期显著增大,分别于第4天和第2天出现最大值,随日龄的增加而增大,且不同日龄间也有显著差异,随后增加平缓。这与本研究结果类似。同样,Drew (1969)研究表明,昆士兰实蝇的雄虫精泵长度随日龄的增加而增大,1~5日龄期间,其呈现剧烈增加趋势,且不同日龄间也有显著差异,随后增加平缓,这与柑橘大实蝇类似。一些学者报道,昆虫精巢在发育后期存在明显缩短现象(Seoetal.,1990),如欧洲玉米螟Ostrinianubilalis(Hübner)(Panagiotis &David,2010)、斜纹夜蛾SpodopteralituraFabricius(徐博等,2012)、卡尼鄂拉蜂ApismelliferacarnicaPollman(薛运波等,2006)、黄钩蛱蝶Polygoniac-aureum(Linnaeus)、(Hiroyoshi &Reddy,2018)。本研究和高熹等(2013)研究结果中,均未出现这种精巢明显缩短现象。这可能与本试验中雌雄成虫未进行交配有关。
本试验中,恒温和变温条件下饲养的雄虫的精巢宽度、精泵的长度和宽度以及指数、精泵内骨骼宽度均无明显差异。变温条件下饲养的雄虫精巢长度及指数、精泵内骨骼长度及指数均分别显著高于其在恒温条件下饲养的雄虫精巢长度、精泵内骨骼长度及指数。这进一步表明,变温(室温)比恒温更有利于柑橘大实蝇雄虫的精巢和精泵内骨骼的发育。诸多研究表明,与恒温相比,变温条件刺激了昆虫的生长发育。Foleyetal.(1981)认为,在相同温度时,与恒温相比,温度变化幅度较大的变温条件能加速未滞育和滞育前的棉铃虫蛹的羽化速率。Radmacher &Strohm (2011)研究了三组恒温和变温条件下OsmiabicornisL.的发育历期,结果表明,变温条件能加速大多数虫态的发育。波动的温度促进了小菜蛾PlutellaxylostellaL.及其天敌——弯尾姬蜂Diadegmainsulare(Cresson)的生长发育(Baharetal.,2012),使麻蝇Sarcophagaargyrostoma(Robineau-Desvoidy)和亮绿蝇LucilliaillustrisMeigen的发育加快(Niedereggeretal.,2010)。但也有研究表明,与恒温(13 ℃)相比,变温(5~29 ℃,平均13 ℃)使红头丽蝇CalliphoravicinaRobineau-Desvoidy和黑颊丽蝇CalliphoravomitoriaL.的发育延长1 d (Niedereggeretal.,2010)。造成研究结果不同的原因可能与昆虫种类以及变温范围有关,变温范围超出了该种昆虫的适宜范围就会对其生长发育造成不利的影响。
虽然雄虫的精巢长度、精巢宽度、精巢指数、精泵长度、精泵宽度、精泵指数、精泵内骨骼长度、精泵内骨骼宽度、精泵内骨骼指数均在一定程度上反映出日龄变化,但从以下因素考虑选择精泵长度推断其雄虫日龄的方法为最佳,在柑橘大实蝇的田间预测预报上有一定的实际意义。(1)解剖测量难度。精泵形态特征明显,容易识别和测量,与精泵内骨骼解剖相比较,精泵解剖不要撕掉精泵表面的肌肉,精泵解剖更省时省力。精巢辨识度也非常高,但其形状扭曲,其长度测量是分段的,相对精泵和精泵内骨骼一次测量,更耗时。(2)在恒温和室温条件下饲养的雄成虫的精泵长度之间、精泵宽度之间差异均不明显。与选择恒温条件下饲养比较,选择室温条件下精泵长度推断雄虫日龄的方法简单、成本低。(3)交配后器官是否萎缩。本研究模型是未交配状态下获得的,其交配后的雄虫精巢是否萎缩并不清楚。一般来说,在达到性成熟后雄性的内生殖系统状态最佳,之后内生殖系统老化,精巢、附腺等产生精子或精液的器官会先开始萎缩(Seoetal.,1990)。而雄虫在野外,有可能交配或未交配,假如雄虫处于交配后的生理状态,其精巢长度有可能出现萎缩,那么此精巢长度预测模型不适合预测其交配后的雄虫的日龄。精泵的内层是精泵内骨骼,外层是肌肉组织,交配前后内骨骼和肌肉变化较小,因此,以精泵长度判定其雄虫日龄的方法较精巢长度更为可靠。但在室内饲养条件下,利用雄虫精泵长度判定其日龄的方法,应用到田间害虫期预测,需要在田间条件下进一步研究和评估。
