桂西北石漠化区不同植被恢复类型对土壤理化性质、酶活与真菌群落多样性的影响
2023-11-05文仕知韦铄星欧汉彪王智慧
高 风,文仕知,韦铄星,欧汉彪,王智慧
(1.中南林业科技大学 林学院,湖南 长沙 410004; 2.广西壮族自治区林业科学研究院 国家林业和草原局中南速生材繁育实验室,广西优良用材林资源培育重点实验室,广西 南宁 530002)
喀斯特地区的石漠化问题是全球范围内的重大生态、经济和环境问题[1]。我国是世界上喀斯特面积最大、分布最广的国家。现阶段,我国喀斯特地区的经济水平仍然相对落后,贫困人口相对集中,人地矛盾非常突出,掠夺性的山地植被破坏活动时有发生,导致土壤侵蚀作用加剧,进而降低水分、养分的调蓄能力[2]。选择合适有效的治理方式实现植被恢复已成为我国南方喀斯特石漠化治理的重点工作之一。
土壤酶是土壤生物化学过程的主要参与者,参与生态系统的物质转化和养分循环过程[3],能灵敏地反映不同土地利用方式下生物化学过程的方向和强度[4],可以体现土壤生物化学过程对土壤理化性质和土壤生物群落的响应[5]。吴丽芳等[6]研究发现,重度石漠化土壤的脲酶、脱氢酶活性最高;中度石漠化土壤的酸性磷酸酶活性最高,淀粉酶、脲酶活性最低;轻度石漠化土壤的酸性磷酸酶活性最低。潘复静等[7]对喀斯特地区不同恢复阶段植物根际土的研究发现,土壤养分与酶活性之间多呈显著正相关。
土壤微生物是土壤生态系统的重要组成部分,参与多种生化反应过程,在物质循环和能量流动过程中具有关键性作用,是化学循环的推动者[8]。真菌作为生态系统中的主要微生物类群之一,其物种组成、功能性状与环境间的相互作用机制不容忽视。真菌喜好酸性环境,而喀斯特森林土壤的pH值通常较其他地区高[9],因此其真菌数量、种类明显少于其他地区[10]。真菌在受到环境影响的同时,也反作用于环境,它广泛地参与纤维素、木质素等的分解,还可以与植物根系共生形成菌根,二者的关系十分密切。
虽然学界已就喀斯特生态系统土壤真菌多样性开展了大量研究,但大多集中于群落结构、植被演替等方面:陈惠君等[11]对喀斯特峰丛洼地不同森林类型土壤真菌的研究发现,土壤真菌与土壤理化因子显著相关;曾庆飞等[12]对石漠化地区不同植被演替阶段的研究发现,石漠化程度越轻,土壤中真菌类群的遗传多样性越优。目前,针对喀斯特地区不同植被恢复模式下土壤真菌群落多样性变化的研究甚少,人们对其土壤养分、酶活性与土壤真菌间的相互关系知之甚少。为此,本文拟就喀斯特地区不同植被恢复模式下土壤真菌群落多样性的差异及其与土壤养分的关系进行探究,研究成果可为该地区的生态恢复和综合治理提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于中国科学院环江喀斯特生态系统观测研究站,地处桂西北河池市环江毛南族自治县大才乡,整体地貌为低山丘陵,是典型的喀斯特峰丛洼地,海拔272.0~647.2 m,60%的坡地坡度大于25°。受砍伐、樵采、垦殖、火烧等人为活动干扰,当地植被曾遭受大面积的破坏,自1984年起,经过20多年的自然恢复,至2006年本试验开始时当地植被处于草丛和草灌阶段。
试验区属中亚热带季风气候,年均气温19.9 ℃,极端最低温-5.2 ℃,极端高温38.7 ℃,全年无霜期300~330 d,≥10 ℃积温为5 500~6 530 ℃,年均日照1 600~1 800 h,平均太阳总辐射量413.8 kJ·cm-2,年均降水量1 389.1 mm,降水丰富,但季节分布不均,雨季降水量占全年降水量的70%以上,多年平均相对湿度82%。土壤由白云岩发育而来,为深色或棕色石灰土,洼地基岩裸露面积占15%,土层较薄,一般在10~50 cm。地表多覆盖碎石(粒径>2 mm),表土碎石体积含量可达10%~40%,土壤质地为黏土和黏壤土。
1.2 植被特征
研究区植被是人工干扰封育后恢复的次生林,共有维管束植物91科206属241种,其中:蕨类植物门有15科19种;种子植物门下有裸子植物亚门2科2种,有被子植物亚门双子叶植物纲63科173种、单子叶植物纲11科47种,属种数量占优势的科为大戟科(18种)、菊科(16种)和禾本科(18种),其他科内属种数量不少于5的有防己科(5种)、蔷薇科(8种)、碟形花科(7种)、桑科(6种)、鼠李科(5种)、芸香科(5种)、茜草科(5种)、马鞭草科(6种)、百合科(5种)和莎草科(9种),以上13个科共计有植物113种,占全部种数的46.9%。
依据植物生长型分类系统,目前人为干扰区内共有草本植物107种、灌木73种、藤本29种、小乔木至乔木32种。常见的植物有30多种,其中,圆叶乌桕(SapiumrotundifoliumHemsl.)、华南皂英[Gleditsiafera(Lour.) Merr.]、黄荆(VitexnegundoL.)、构[Broussonetiapapyrifera(Linn.) L’Hér. ex Vent.]、任豆(ZeniainsignisChun)、红背山麻秆[Alchorneatrewioides(Benth.) Muell. Arg.]、盐麸木(RhuschinensisMill.)、银合欢[Leucaenaleucocephala(Lam.) de Wit]、竹叶花椒(ZanthoxylumarmatumDC.)、八角枫[Alangiumchinense(Lour.) Harms]、浆果楝[Cipadessabaccifera(Roth.) Miq.]、火棘[Pyracanthafortuneana(Maxim.) Li]、金樱子(RosalaevigataMichx.)、白茅[Imperatacylindrica(L.) Beauv.]、蔓生莠竹[Microstegiumfasciculatum(Linnaeus) Henrard]、鬼针草(BidenspilosaL.)等是该区域植被群落的主要建群种或共建种,大部分为阳性先锋物种,但由于干扰强度大,退化严重。
1.3 样地设置
于2006年底,在一面东南向山坡中下部修建了4个大型植被恢复试验小区(24°44′~24°45′N,108°19′~108°20′E),分别为自然封育林(ENF)、落叶阔叶林(DF)、落叶常绿阔叶混交林(EDF)和常绿阔叶林(EF)。其中,ENF的处理方式为保留原始植被,作为对照;DF的处理方式为在原始植被上挖坑,种植落叶乔木;EDF的处理方式为在原始植被上挖坑,种植常绿与落叶乔木;EF的处理方式为在原始植被上挖坑,种植常绿乔木。将各小区的基本情况整理于表1。在每个试验小区内选择代表性地带设置3个20 m×20 m的固定监测样地[13]。
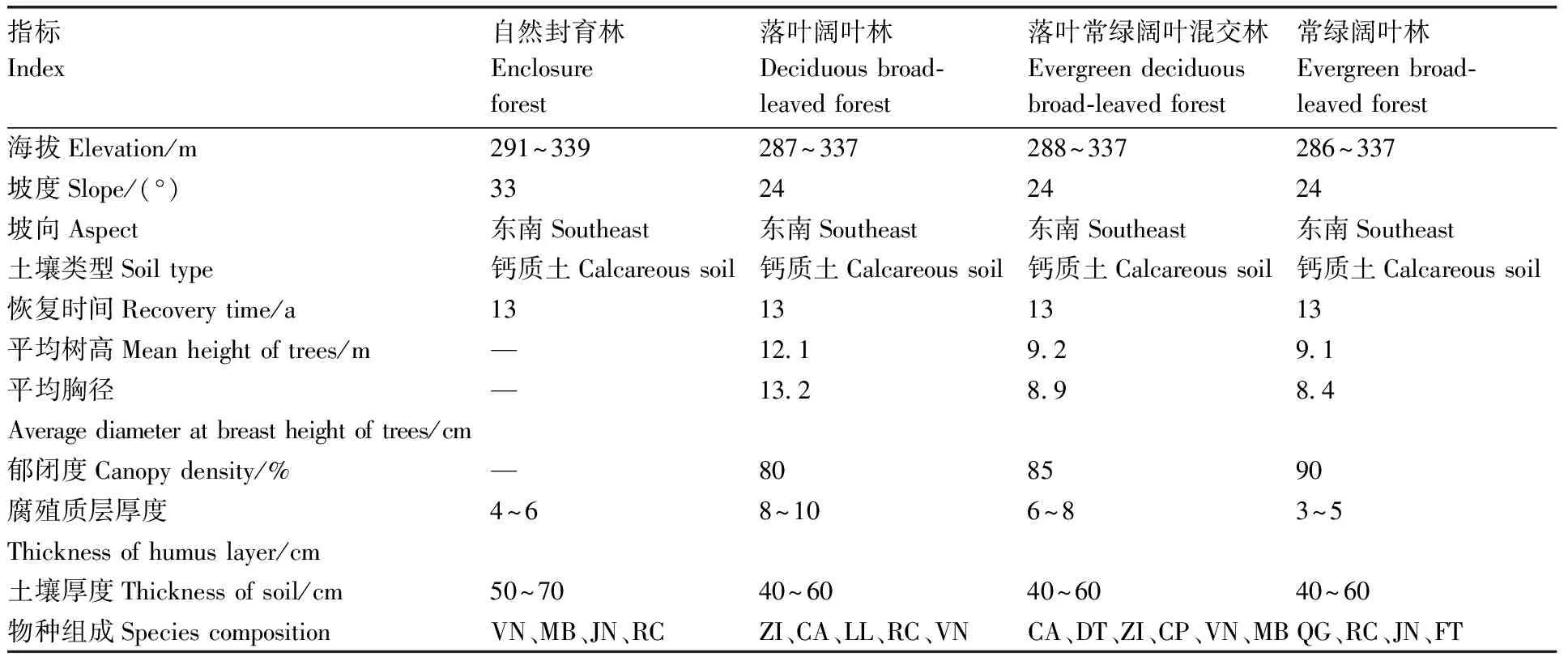
表1 样地基本概况
1.4 样品采集
在各检测样地内使用事先消毒的灭菌直径1 cm的土钻采集0~20 cm的表层土,每个样地按“S”形布设18个取样点,取回的样品混合均匀并去杂,分成3份:一份过2 mm的筛后,于4 ℃冰箱保存,用于测定土壤酶活性;一份自然风干、去杂、过2 mm的筛后,用于土壤理化性质测定;另一份按4分法取1份放入2根已灭菌的离心管中,置于液氮中保存,带回实验室,保存于-80 ℃的冷冻柜中,用于土壤微生物高通量测序。
1.5 指标测定
参照文献[14-15]:采用玻璃电极法测定土壤pH值,采用硫酸-重铬酸钾氧化法测定土壤有机碳含量,采用凯氏定氮法测定土壤全氮含量,采用NaOH水解扩散法测定土壤速效氮含量,采用钼蓝比色法测定土壤速效磷含量,采用火焰光度法测定土壤速效钾含量。
分别采用磷酸苯二钠比色法、高锰酸钾滴定法、苯酚钠次氯酸钠比色法、3,5-二硝基水杨酸比色法测定土壤碱性磷酸酶、过氧化氢酶、脲酶、蔗糖酶活性[16]。
基于Illumina Novaseq 6000测序平台构建土壤真菌的小片段文库并进行测序。简述如下:将处理好的土样用TGuide S96磁珠法土壤/粪便基因组DNA提取试剂盒[天根生化科技(北京)有限公司]提取基因组DNA。根据检测报告中PCR预扩增结果选择扩增条件,并以稀释后的DNA为模板进行扩增,真菌的扩增引物为ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2(5′-GCTGCGTTCTTCATCGATGC-3)。使用MiSeq Reagent Kit v3试剂盒(美国Illumina)构建Illumina文库。用QIAquick Gel Extraction Kit试剂盒(德国QIAGEN,)纯化PCR产物,在Illumina MiSeq PE300平台(美国Illumina)上进行高通量双端测序。
1.6 数据处理
采用Excel 2019软件进行数据录入与整理。采用SPSS version 22.0统计软件进行方差分析,对有显著(P<0.05)差异的,采用Tukey-HSD检验进行多重比较。采用FUNGuild v1.0软件对土壤真菌群落功能进行预测分析,用R 4.0.0软件进行主坐标分析、相关性分析。
2 结果与分析
2.1 不同植被恢复类型的土壤养分
DF和EDF的土壤有机碳含量显著(P<0.05)高于EF和ENF(表2)。DF、EDF和EF的全氮含量无显著差异,均显著高于ENF。土壤速效钾含量由高到低依次为DF>ENF>EDF>EF,4种植被恢复类型间均差异显著。土壤速效氮含量由高到低依次为DF>EDF>ENF>EF,且均差异显著。土壤速效磷含量由高到低依次为EDF>DF>EF>ENF,且均差异显著。不同植被恢复类型的土壤pH值在6.59~7.72,DF和ENF的土壤pH值显著高于EDF和EF。
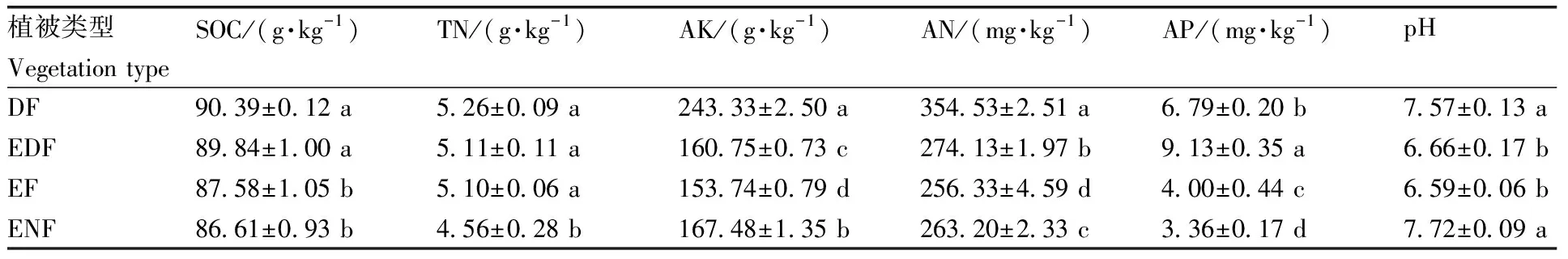
表2 不同植被恢复类型下土壤的基本理化性质
2.2 不同植被恢复类型的土壤酶活性
不同植被恢复模式下,土壤过氧化氢酶活性无显著差异(图1)。与对照(ENF)相比,EDF、DF和EF处理的土壤蔗糖酶活性显著升高;此外,DF和EDF处理显著提高了土壤脲酶和碱性磷酸酶的活性。
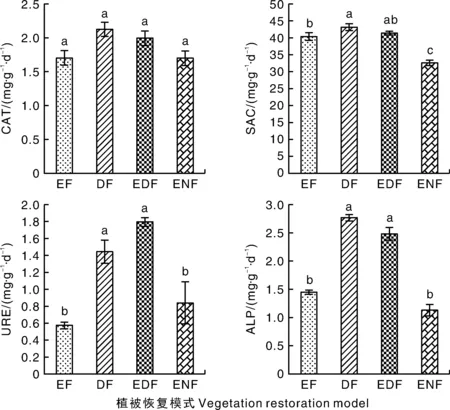
CAT,过氧化氢酶;SAC,蔗糖酶;URE,脲酶;ALP,碱性磷酸酶。ENF,自然封育林;DF,落叶阔叶林;EDF,落叶常绿阔叶混交林;EF,常绿阔叶林。下同。柱上无相同字母的表示差异显著(P<0.05)。CAT, Catalase; SAC, Saccharase; URE, Urease; ALP, Alkaline phosphatase. ENF, Enclosure forest; DF, Deciduous broad-leaved forest; EDF, Evergreen deciduous broad-leaved forest; EF, Evergreen broad-leaved forest. The same as below. Bars marked without the same letters indicate significant difference at P<0.05.图1 不同植被恢复类型下的土壤酶活性Fig.1 Soil enzymes activities under vegetation restoration models
2.3 不同植被恢复类型的土壤真菌多样性
2.3.1 土壤真菌群落物种组成
将各植被恢复类型下门水平上土壤真菌相对丰度排名前10的绘制于图2。将相对丰度大于10%的真菌菌落定义为优势菌落,将相对丰度大于1%、小于10%的真菌菌落定义为亚优势菌落,将优势菌落与亚优势菌落统称为主导菌落。根据上述定义,不同植被恢复类型区的土壤真菌群落优势菌门有子囊菌门(Ascomycota)、担子菌门(Basidiomycota),亚优势菌门为被孢霉门(Mortierellomycota),其余相对丰度排名前10的菌门还有罗兹菌门(Rozellomycota)、球囊菌门(Glomeromycota)、梳霉亚门(Kickxellomycota)、捕虫霉门(Zoopagomycota)、壶菌门(Chytridiomycota)、Calcarisporiellomycota、油壶菌门(Olpidiomycota)。
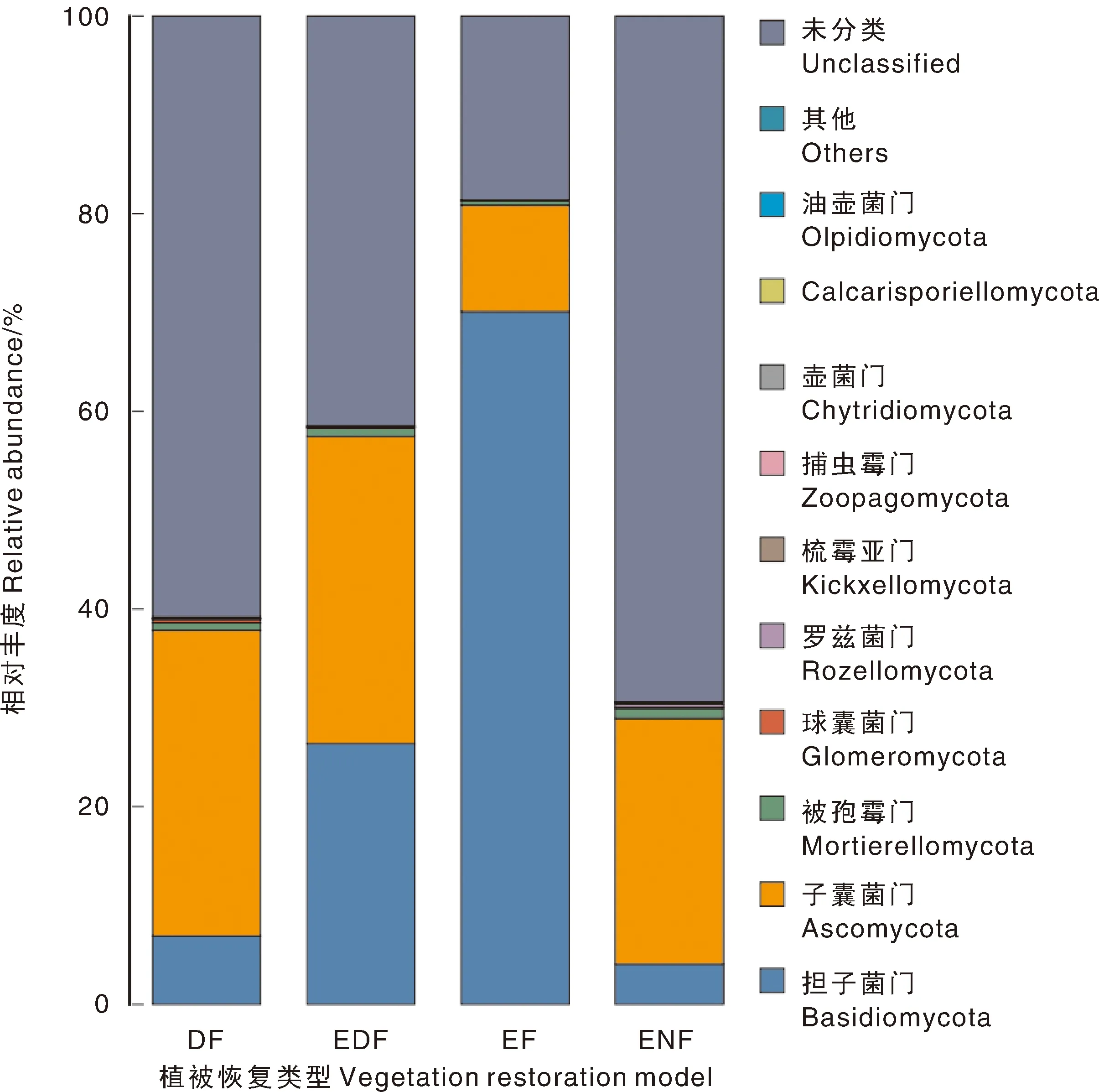
图2 门分类水平上真菌优势种的OTU(运算分类单元)丰度堆积柱状图Fig.2 OTU (operational taxonomic unit) abundance accumulation histogram of soil dominant fungal species at phylum level
不同植被类型下的优势菌门及其占比存在区别:DF主要为子囊菌门(30.94%),EDF主要类为子囊菌门(31.07%)和担子菌门(26.38%),EF主要为担子菌门(70.09%)、子囊菌门(10.76%),ENF主要为子囊菌门(24.88%)。
整体来看,人工林土壤真菌OTU(运算分类单元)的相对丰度均高于ENF,其中,子囊菌门和担子菌门是优势菌门,共占真菌菌门总丰度的50%以上,其次是被孢霉门、罗兹菌门、球囊菌门、梳霉亚门、捕虫霉门、壶菌门、Calcarisporiellomycota和油壶菌门。总体上看,担子菌门是相对丰度最高的菌门,其在EF中的相对丰度显著大于ENF、DF。
2.3.2 真菌Beta多样性分析
采用Bray-Curtis距离算法,结合OTU丰度,绘制Permanova分析箱形图(图3)。对不同植被恢复类型下真菌间的Beta多样性进行差异显著性检验,R2=0.423,不同植被恢复类型的组间差异大于组内差异,P<0.05,说明检验的可信度高。
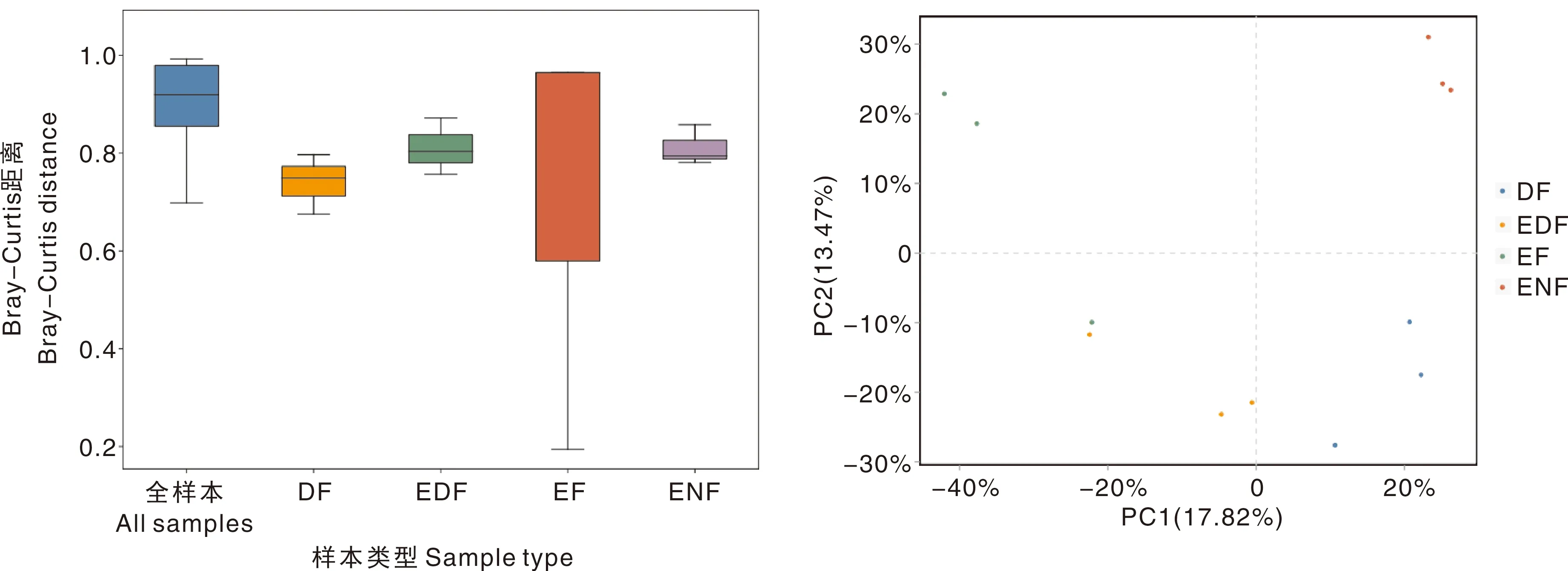
PC1,第一主成分;PC2,第二主成分。PC1, Principle component 1; PC2, Principle component 2.图3 相似性分析箱形图(左)与主坐标分析图(右)Fig.3 Similarity analysis box diagram (left) and principal coordinate analysis diagram (right)
利用各样本注释序列间的进化信息,结合Bray-Curtis距离算法,进行主成分分析。第一主成分(PC1)能够解释样本原始特征的17.82%,第二主成分(PC2)能够解释样本原始特征的13.47%。从组内来看:ENF、DF和EDF的样本聚类较集中,组内真菌群落结构差异较小,且以ENF最小;EF样本的聚类异常分散,其组内差异超过组间差异,说明组内真菌群落结构的差异非常大。从组间来看:PC1上,DF和ENF样本点相距较近,表明这两组的真菌群落结构较为相似,EF与ENF样本点相距较远,表明两者的真菌群落结构差异较大;PC2上,EDF与DF的样本点交错在一起,表明二者组间的真菌群落结构较为相似,但与ENF、EF距离较远。
2.3.3 土壤真菌FUNGuild功能预测
采用FUNGuild软件对不同植被恢复类型的土壤真菌群落进行功能预测分析(图4),发现各植被恢复类型的土壤真菌营养类型主要包括腐生营养型(23.37%~46.50%)、病理营养型(7.64%~47.57%)和共生营养型(7.93%~68.99%)3种,其中,ENF、EDF以腐生营养型为主,DF以病理营养型为主,EF以共生营养型为主。与对照相比,DF提高了病理营养型真菌的相对丰度,EDF、EF提高了共生营养型真菌的相对丰度,3种人为植被恢复模式均降低了腐生营养型真菌的相对丰度。
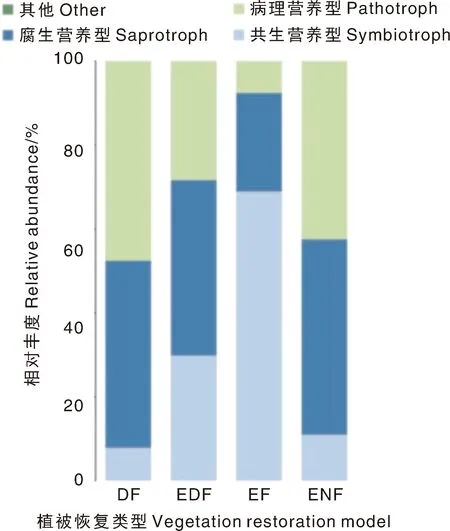
图4 基于FUNGuide预测的不同植被恢复类型下土壤真菌的营养型组成Fig.4 Nutrient patterns of soil fungi under vegetation models based on FUNGuide predictions
2.4 土壤养分、酶活性与真菌的相关性
对土壤化学性质、酶活性、真菌主导菌落进行相关性分析(图5),结果显示,土壤养分、酶活性与真菌主导菌落大多具有显著相关性,其中,土壤pH值与土壤酶活性、真菌丰度多呈负相关,土壤速效氮含量与脲酶活性呈显著正相关,土壤速效钾、有机碳含量与过氧化氢酶活性呈显著正相关,土壤蔗糖酶活性与土壤养分指标、真菌丰度多呈负相关,担子菌门、壶菌门、捕虫霉门、Calcarisporiellomycota与土壤养分、酶活性多呈正相关,子囊菌门、油壶菌门、梳霉亚门、罗兹菌门、被孢霉门与土壤养分、酶活性多呈负相关。
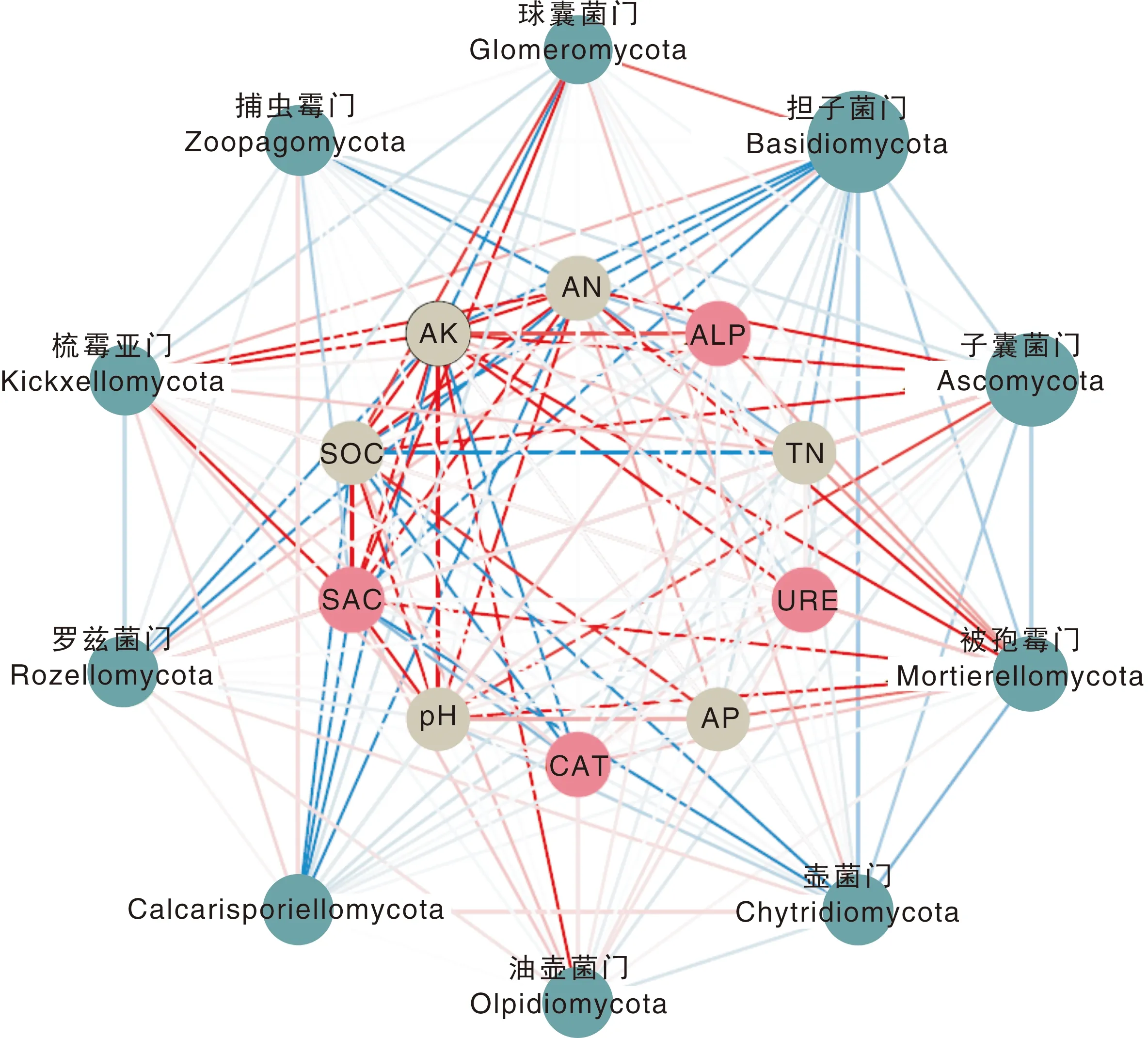
SOC,土壤有机碳;TN,全氮;AK,速效钾;AN,速效氮;AP,速效磷。图中绿色圆点代表真菌主导菌门,圆点大小表示其相对丰度大小,红色圆点代表土壤酶活性指标,灰色圆点代表土壤养分指标,蓝线表示显著(P<0.05)正相关,红线表示显著(P<0.05)负相关,红、蓝颜色的深浅代表相关系数的高低。SOC, Soil organic carbon; TN, Total nitrogen; AK, Available potassium; AN, Available nitrogen; AP, Available phosphorus. The green dots in the figure represent the dominant species of fungi. The size of the dots indicates the relative abundance of the dominant species. The red dots represent the indicators of soil enzymes activities, and the grey dots represent the indicators of soil nutrients. The blue lines indicate significant (P<0.05) positive correlations, and the red lines indicate significant (P<0.05) negative correlations, and the redder or bluer indicates higher correlation coefficients.图5 土壤养分与酶活性、真菌群落的网络分析图Fig.5 Network analysis of soil nutrients, enzymes activities and fungal communities
3 讨论
3.1 不同植被恢复类型的土壤养分和酶活性
C、N、P是植物营养和土壤肥力的核心成分。土壤酶可以催化土壤生物化学反应,参与土壤中的物质循环与转化,反映土壤生物活性的强弱,是土壤肥力的标志之一[17]。在本研究为期13 a的植被恢复试验中,土壤养分含量得到有效改善,DF的各土壤养分指标均较对照(ENF)显著提高,EF和EDF还较对照显著降低了土壤pH值。
丰富的凋落物分解后能够增加土壤有机碳含量,从而缓解土壤酸化[18]。DF处理多种植落叶阔叶树种,可产生大量易腐烂的枯枝落叶,有助于增加穴居蚯蚓群落,抑制土壤酸化。DF和EDF显著提高了土壤有机碳含量,可能是因为落叶树种为有机碳的输入作出了巨大贡献。在各处理中,DF和EDF的4种土壤酶活性均相对较高,且其土壤蔗糖酶、脲酶和碱性磷酸酶活性显著高于对照,但在土壤过氧化氢酶活性上无显著差异。这表明落叶树种的存在显著提高了土壤中水解酶的活性,但对氧化还原酶的活性没有显著影响。
3.2 不同植被恢复类型的真菌多样性
研究发现,不同植被恢复类型的土壤真菌在门水平的组成上具有一定的相似性,在相对丰度上存在一定差异。各植被恢复类型的主导真菌菌门为子囊菌门、担子菌门和被孢霉门,但不同植被恢复类型明显改变了土壤真菌群落的组成比例,这与已有的研究结果类似[19]。总体上,人工林的土壤真菌OTU丰度高于对照,子囊菌门和担子菌门在不同植被恢复模式的土壤真菌中均占有绝对优势。在DF、EDF、ENF中,子囊菌门为绝对优势真菌,这与已有的研究结果类似[11,20]。子囊菌门主要是腐生菌,可分解土壤中的有机质和动植物尸体。EF的子囊菌门相对丰度明显低于其他植被恢复类型,且与其他植被恢复类型的土壤真菌群落结构也相差较大,这可能是由于EF多为常绿阔叶树种,凋落物较少,而其他3种植被恢复类型中均包含落叶树种,均可产生较多的凋落物,更适于子囊菌门等真菌群落的生存。EF的担子菌门相对丰度远高于ENF,这主要是因为常绿阔叶林通气性良好,且土壤pH值较低,有利于提高担子菌门的丰度[21],但这也导致了其余菌门的生态位被压缩,反而使得其整体菌落的多样性降低。不同植被恢复类型的土壤真菌功能类型差异较大,但整体上以腐生营养型和病理营养型为主,这与以往在石漠化区开展的研究结论一致[22]。
3.3 土壤养分、酶活性与真菌的关系
土壤酶和微生物共同参与土壤的生化过程,土壤养分为土壤微生物提供碳源和氮源,土壤微生物在一定程度上决定土壤酶的种类和活性,由此可见,土壤真菌、酶活性与土壤养分间存在密切的联系[23]。本研究发现,土壤pH值与土壤酶活性、真菌群落多呈负相关,这与已有的研究结果类似[24],可能是因为土壤pH值能够影响土壤化合物,从而对真菌的养分吸收产生影响,最终影响真菌分泌土壤酶[25]。本研究中,土壤有机碳、全氮、速效钾、速效氮和速效磷与土壤酶活性、真菌群落结构存在显著相关性,这与以往的研究结果一致[26]。与对照相比,DF和EDF显著增加了土壤速效氮含量,且以DF的增幅更大,土壤脲酶活性与土壤速效氮含量呈正相关,相应地,DF和EDF的土壤脲酶活性也更强,反过来,土壤中脲酶能够分解酰胺氮,水解有机质分子中的肽键,生成铵离子,进而增加土壤有效氮含量[27]。EF的凋落物多为革质,不易分解,不利于有机物向土壤的输入,从而影响土壤脲酶活性和速效氮含量的提升;而落叶树种则可以为表层土壤带来丰富的有机质。磷的主要直接来源是土壤有机质的分解,土壤中的大部分磷以有机磷化合物的形式存在。土壤中的碱性磷酸酶可以催化有机磷化合物的水解,增加土壤有效磷含量[28]。在本研究中,DF和EDF的碱性磷酸酶活性显著高于对照,相应地,其土壤速效磷含量也显著更高。
DF以子囊菌门为优势菌门,子囊菌门与土壤pH值、速效钾含量、有机碳含量、酶活性呈负相关,这与彭辉等[27]发现子囊菌门在速效钾含量较低的土壤中相对丰度较高的研究结果一致,但也导致DF中共生营养型的相对丰度最低,而共生营养型真菌有利于植物的生长,可降低病虫害风险[29]。EF以担子菌门为优势菌门,担子菌门与土壤养分、酶活性多呈正相关,且其病理营养型真菌的相对丰度在各植被恢复类型下最低。病理营养型真菌可从宿主细胞吸收养分,导致植物营养不足,从而引起植物病害,抑制植物生长[30]。这说明,EF处理有利于植物的生长。腐生营养型真菌的相对丰度表现为3种人工林小于ENF,表明自然恢复更容易引起腐生营养型真菌的聚集。多种土壤真菌与土壤过氧化氢酶、蔗糖酶、脲酶、碱性磷酸酶活性呈正相关,这可能是因为微生物群落与土壤酶活性之间具有自相关性[31],人为植被恢复模式下,土壤养分含量的增加,促进了真菌数量的增加,相应带动真菌分泌的酶量增多、活性增强。
4 结论
与对照相比,人工林的土壤全氮、速效磷含量与土壤蔗糖酶活性显著提高,DF和EDF处理的土壤有机碳含量显著升高,EDF和EF处理的土壤pH值显著降低,DF和EDF处理的土壤碱性磷酸酶活性显著升高。在人工林土壤真菌中,子囊菌门和担子菌门是最优势的菌门,占真菌菌门总丰度的50%以上。与对照相比,DF提高了病理营养型真菌的相对丰度,EDF、EF提高了共生营养型真菌的相对丰度。总的来看,相较于自然封育林,人工林有助于增加土壤养分含量,提高土壤酶活性,促进真菌丰度的增加。
