基于转录组测序与定量PCR技术挖掘北沙柳株型相关候选基因
2023-11-05贺玉娇阿拉腾苏和王爱君韩若霜孙贵荣张国盛
贺 嵘,赵 恺,贺玉娇,阿拉腾苏和,王爱君,宁 静,韩若霜,孙贵荣,张国盛,*
(1.内蒙古农业大学 林学院,内蒙古 呼和浩特 010019; 2.鄂尔多斯市造林总场,内蒙古 鄂尔多斯 014300; 3.内蒙古林业科学研究院,内蒙古 呼和浩特 010013; 4.内蒙古自治区林业和草原种苗总站,内蒙古 呼和浩特 010010)
生物体的表型性状是基因和外界环境共同作用下的外在体现,是可以直接观测的性状,受到一个或多个基因共同调控。李忠南等[1]报道玉米(Zeamays)的分蘖率由4对主基因与多对微效基因功能决定的。主基因决定性状的稳定性,多对微效基因决定该性状的表现。糯玉米种子的活性,是由1对主基因和几个基因协同调控的,但以主基因遗传为主[2]。WRKY家族参与调控植物的生长发育等生理过程,拟南芥(Arabidopsisthaliana)中WRKY34与WRKY2共同调控植物花粉的发育[3]。WRKY转录因子也可以通过调控HY5的表达来调控植物下胚轴的生长发育[4]。在水稻(Oryzasativa)理想型形成过程中克隆得到IPA1基因,其功能获得性突变体表现为分蘖数少、抗倒伏、茎秆粗壮[5],IPA1不仅可以结合DEP1来调控水稻的株型与产量,还可以增强水稻的抗病性[6]。植物基因的调控功能是多方位的,多层次的综合研究分析对挖掘完善基因功能是十分必要的。植物株型是影响其生物量和利用效率的关键性状之一,开展株型定向培育的重要意义,已经在农作物及经济树种育种中得到了充分的证明。然而植物株型性状多数是数量遗传性状,由多个彼此独立或协同的基因共同决定的,呈现出增效或减效的作用,采用主效基因的评价方法,难以实现育种目标。
北沙柳(Salixpsammophila)别称沙柳或西北沙柳,为杨柳科柳属的速生、多年生灌木,是中国西北地区沙产业的重要资源树种。北沙柳产于毛乌素沙地和库布齐沙漠,西北、华北等地区均有引种栽培。北沙柳根系发达,繁殖和萌蘖力强,抗旱耐贫瘠,抗风沙,耐盐碱,是西北地区特色树种,也是“三北”地区土地荒漠化防治的首选树种[7]。由于北沙柳是木型材、纤维板、刨花板、造纸、柳编、医药、活性炭等产业的重要原料,目前已经形成了以北沙柳为原料的新兴沙产业链。北沙柳的株型呈现直立型、开放型、中间型等特征。不同的利用方式对北沙柳株型的要求不同:作为木材产业化的原料,理想的株型是通直少(或无)侧枝的直立型;而防沙固土的理想株型是多分枝开散生长的开散型。如何能够通过现代生物技术,结合北沙柳生物学特性,开展快速精准定向化育种,以满足沙产业发展的需要是北沙柳资源精细化利用的主要问题。
本研究利用定量PCR技术和转录组测序技术相结合的方法,分析水培预测群体和野外训练群体不同株型无性系基因表达量的变化特征,以及与株型特征值冠高比的相关性。创建基于冠高比相关系数的北沙柳株型后备基因初步筛选方法,为北沙柳株型定向育种亲本筛选提供理论基础,具有重要的现实意义。
1 材料与方法
1.1 实验材料
本研究试验材料全部来自于鄂尔多斯市造林总场沙柳国家林木种质资源库的基本群体(2008年种植)和无性系测定林(2017年营造)。采样部位为茎的尖端(茎尖)和茎尖以下前3个节间的嫩茎(腋芽)。
2021年5月种质资源库的基本群体中筛选9个北沙柳无性系(即直立型、中间型、开散型各3个)分别采集枝条,在内蒙古农业大学智能温室(平均温度27 ℃、平均湿度45%)中进行水培构成预测群体,水培2个月采集茎尖和腋芽。
2021年7月按照生长旺盛、病虫害少且株型特征有明显变化的标准从种质资源库的无性系测定林中筛选100个左右的无性系,标记后组成北沙柳训练群体,并采集茎尖和腋芽(液氮低温保存待用)。
1.2 研究方法
1.2.1 株型特征的调查
2020年9月和2021年9月分别对无性系测定林中200个无性系的株高、冠幅(东西向和南北向)、地径、萌生枝条数量、叶片特征等指标进行了调查,计算株型特征值冠高比(平均冠幅与高度之比)。两年计算获得的冠高比呈现相同的趋势,既能体现无性系间的差异,又具有稳定性,且具有正态分布趋势,是体现株型性状较理想的特征值(图1)。

A,2020年;B,2021年。A is the training group in 2020; B is the training group in 2021.图1 无性系群体冠高比变化趋势Fig.1 Trends in the ratio of crown and height of training groups
1.2.2 北沙柳株型组的划分
依据无性系间冠高比变化特征,按照“冠高比平均值±0.5倍标准差”将从无性系测定林中筛选的97个北沙柳无性系划分为6个株型组(表1)。
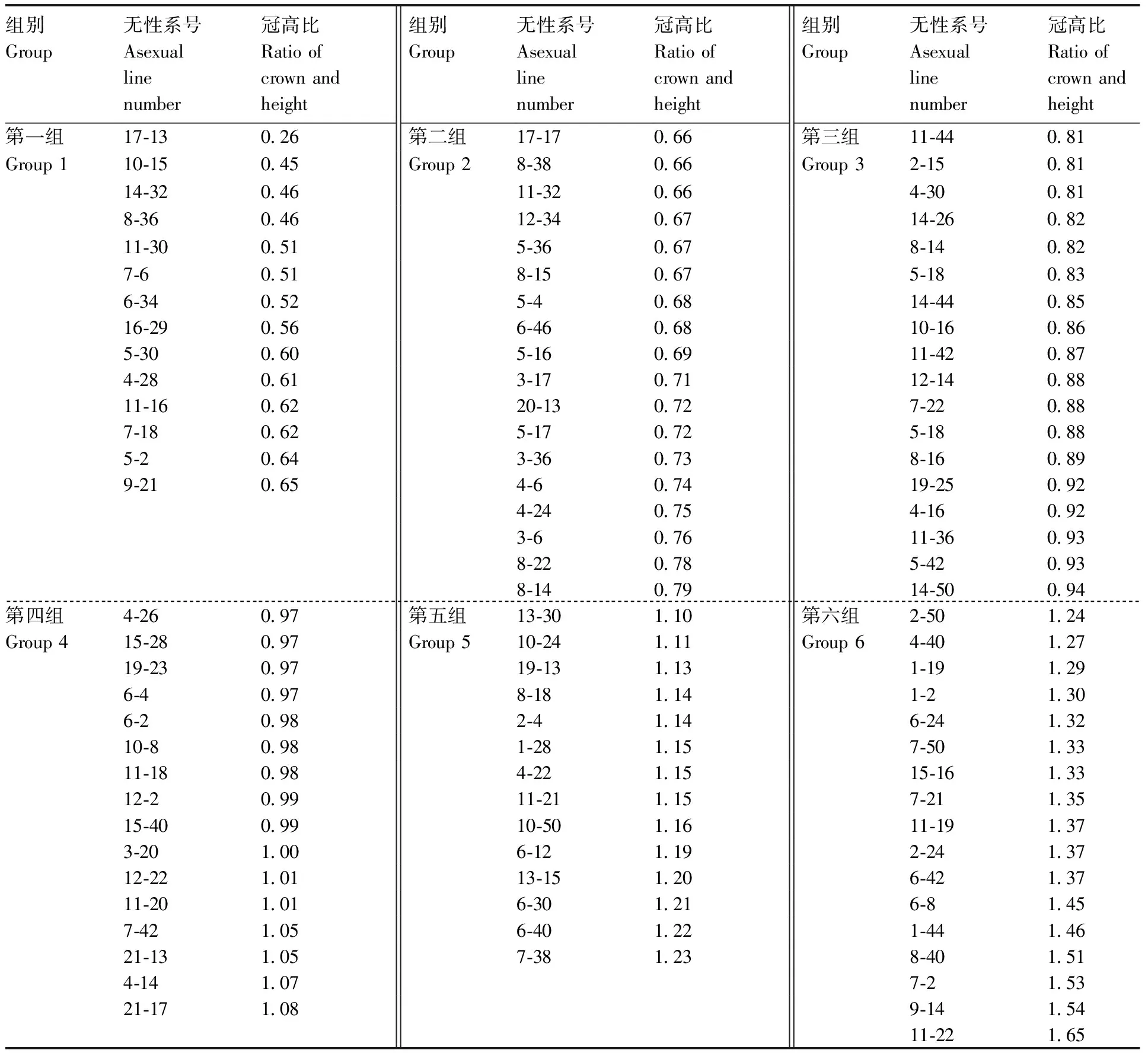
表1 北沙柳无性系株型组
1.2.3 RNA的提取及cDNA的合成
采用TIANGEN RNAprep Pure Plant Plus Kit试剂盒(天根生化科技有限公司,北京)提取北沙柳腋芽与茎尖的RNA,利用TIANGEN FastKing gDNA Dispelling RT SuperMix试剂盒(天根生化科技有限公司,北京)将RNA反转录成cDNA,在-80 ℃冰箱中进行保存。
1.2.4 转录组测序及预测值计算
将水培测试群体(9个北沙柳无性系)腋芽及茎尖RNA送到基迪奥生物科技有限公司进行转录组测序,获得40 049条序列,其中与桤柳(Salixpurpurea)对应的序列有37 865条,未知序列有2 184条。利用Excel中CORREL的平方计算所有基因序列的FPKM值与冠高比的预测值(相关系数),相关系数0.8~1.0为极强相关,0.6~<0.8为强相关,0.4~<0.6为中相关,0.2~<0.4为弱相关,0~<0.2为极弱相关(表2)。

表2 部分基因序列的相关性系数
1.2.5 目标基因的选择
为了验证目标基因在训练群体中的表达特征,探讨预测值与北沙柳的株型是否相关,本研究的目标基因包括已报道的与北沙柳分枝相关的基因SpsLAZY1b[8]、SpsTAC2[9],其他植物中与株型相关的基因ZFP4[10]、TB1[11]、SPA2[12]、ABF2[13]和PYL1[14],以及本研究转录组测序结果中预测值较高的ATX1[15]、FHY1[16]和RFK1[17]共10个基因组成。
1.2.6 引物设计
将从转录组测序结果中筛选的目标基因登录号在数据库Phytozome 13(https://phytozome-next.jgi.doe.gov/)中进行比对,获得对应的桤柳编码区序列,根据编码区序列用Primer3Plus(https://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi)设计引物(表3),引物由擎科公司(北京)合成。

表3 引物序列
1.2.7 定量PCR
将反转录的cDNA稀释3倍作为模板,将UBQ作为内参基因,目的基因和内参基因均设置3个重复,用Roche LightCycler 480 Ⅱ仪器进行定量PCR测定。反应体系为20 μL,其中TB酶10 μL,cDNA模板1 μL,上下游引物各1 μL,RNase-Free ddH2O 7 μL。程序设置为95 ℃ 5 min;95 ℃ 10 s,60 ℃ 15 s,72 ℃ 15 s,45个循环;95 ℃ 10 s,60 ℃ 1 min;40 ℃ 10 s;4 ℃保存。
基因的相对表达量采用2-ΔΔCT法计算。首先将性状明显、冠高比最小的无性系设为实验对照组,ΔΔCT=实验组ΔCT-对照组ΔCT。
转录组测序的FPKM值与2-ΔΔCT计算结果均表示基因的表达量,但二者只是表达量变化的趋势相同,二者之间没有关系[18]。
1.2.8 秩和分析及多元回归分析
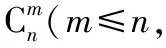
1.2.9 高相关目标基因蛋白互作网络分析
利用STRING(https://cn.string-db.org/)对相关系数高的目标基因分别进行蛋白互作分析。通过分析蛋白质-蛋白质相互作用(protein-protein interaction, PPI)网络图中蛋白结构信息、蛋白与蛋白之间的相互作用以及蛋白与蛋白间的连接程度,来预测蛋白质的功能。
2 结果与分析
2.1 初选目标基因预测值(R2)特征
10个初选目标基因的预测值在测定群体和训练群体的茎尖中虽然存在差异,但基因表达量的变化趋势是相同的。预测值在测定群体中最高为0.924 4,最低为0.001 0。训练群体中最高为0.185 8,最低为0.000 2,训练群体的预测值明显低于测定群体。10个初选目标基因中RFK1、ABF2、TB1的预测值较低,FHY1、TAC2与冠高比呈正相关趋势,其余均为负相关(图2)。

A,水培预测群体;B,野外训练群体。A, The hydroponic prediction population; B, The field training population.图2 茎尖中目标基因表达量趋势Fig.2 Expression trends of target genes in the shoot tips in the hydroponic and training populations
在腋芽中,测定群体的预测值高于训练群体,基因表达量具有相同的变化趋势。预测值在测定群体中变化范围为0.034 8~0.858 9,训练群体中为0.051 7~0.595 1。ATX1的预测值在两个群体中均相对较高,TAC2相对较低。FHY1和TAC2的表达量与冠高比呈正相关,其余都呈现负相关(图3)。
2.2 秩和分析与多元回归分析
按照6个株型组对每个初选目标基因表达量的秩和分析。结果表明,ATX1在茎尖与腋芽,TAC2、ZFP4、SPA2在茎尖,TB1、ABF2、FHY1、RFK1、PYL1在腋芽均有显著差异(P≤0.05)(表4)。

表4 目标基因秩和分析

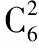
六基因组合中,与冠高比相关系数为0.769,预测模型为y=1.176+0.000×ABF2+0.001×PYL1-0.037×RFK1+0.074×FHY-0.089×TB1-0.460×ATX1。在六基因组合中ABF2、PYL1与冠高比的相关性最小。
所有组合对比发现ATX1对株型形成贡献最大,其次为TB1、RFK1、FHY1、ABF2、PYL1。基因组合中双基因组合(ATX1+FHY1)的相关性较高(0.740)。
2.3 蛋白互作网络分析
将与冠高比相关性较高的ATX1、FHY1、RFK1基因进行蛋白互作分析,预测其功能,置信度为0.7(高等),结果如图4所示。

A, ATX1; B, FHY1; C, RFK1.图4 高相关基因蛋白互作网络Fig.4 Protein interaction network of high correlation genes
在ATX1蛋白互作网络中,与ATX1直接相关的基因有HMA5、RAN1、COPT5和HMA6。COPT1-6为铜转运蛋白,ATX1、CCH、CCS为铜伴侣蛋白。铜蛋白CCS可以提供铜离子,ATX1和CCH可将铜离子传递给HMA5、RAN1,RAN1再将铜离子整合到乙烯受体里,产生乙烯信号,通过调控乙烯的合成来影响植物的生长。
在FHY1蛋白互作网络中,与FHY1直接相关的是PHYA基因。COP1、PHYA、CRY1基因可以通过调控光形态建成来影响植物的生长。AUX1为生长素受体,FHL和FHY1互为同源蛋白。PIFs是一种光敏色素互作蛋白,可以与赤霉素信号通路中的DELLA蛋白相互作用来调控植物的生长发育。
RFK1是与植物育性恢复有关的基因,其与调控植物生长发育方面的基因互作较少,数据支撑较少,但仍具有一定的参考价值,可能在调控北沙柳生长发育方面有着重要作用。
3 讨论
3.1 采样部位的影响
植物的上表型由产生新细胞和器官的分生组织所形成。茎尖分生组织(SAM)促进垂直生长;腋生分生组织(AM)出现在叶腋中,可以产生侧生枝或次生枝。SAM决定了植物叶序,并影响AM的形成[6]。SAM的维持与分化涉及生长素、细胞分裂素和多肽的复杂互作,它们协同调控植物的株型。张磊等[8]通过实验发现,SpsLAZY1a、SpsLAZY1b沉默时,会引起侧枝的水平生长,且基因主要在茎和叶腋处表达。袁梦如等[9]研究发现,TAC2基因与植物的分枝发育有关,且在茎中表达最丰富,其次是叶腋。
茎尖与腋芽都是最活跃的生长点,但茎尖分生组织的分化能力较强,茎尖细胞会通过不断地分裂生长和分化促进茎的伸长来影响植物的高生长;腋芽的分化程度较为稳定,主要影响植物的分枝。本实验采样部位选择了与株型有关的茎尖及腋芽。在大群体中通过qRT-PCR实验证明,北沙柳茎尖和腋芽均与株型均有一定相关性,但腋芽与冠高比的相关系数更接近预测值。这可能是因为茎尖细胞较为活跃,会不断地进行分裂分化和生长,导致营养物质的分配主要促进植物茎的伸长生长而非分枝的生长。
3.2 SpsLAZY1b、SpsTAC2与株型的关系
SpsLAZY1b、SpsTAC2已被证明与北沙柳的分枝有关,但实验结果表明,基因表达量与北沙柳的冠高比相关性较小(腋芽中SpsLAZY1b相关系数为0.1、SpsTAC2为0.051 7)。LAZY1除了可以调控分蘖角度外还可以结合下游基因调控其他性状[19]。在玉米中,ZmLAZY1还可以调节花序的发育[20]。LAZY1可能通过调控一类分子和基因从而调控大量下游基因的表达。在LAZY1下游候选基因中,CKX2、TAW1被报道与水稻的穗粒数有关[21]。研究发现,在杨树中LAZY1a基因过表达会影响其他基因的表达量,如TAC2、PIN1、MAX1等基因的表达量会高于野生型。TAC的表达量也受内源激素影响,在桃树中,TAC的表达量随着生长素含量的升高而减少[22]。在预测值与实验结果中,SpsLAZY1b、SpsTAC2与北沙柳株型相关性较小,为极弱相关。这可能是因为SpsLAZY1b、SpsTAC2在北沙柳株型中为非主效基因,是通过调节其他基因间接来影响株型。
3.3 ZFP4、TB1、SPA2、PYL1和ABF2与株型的关系
ZFP4、TB1、SPA2在其他物种中已被证明是与植物的生长发育有关的基因。影响TB1基因表达的因素有很多,如光照、植物激素等,红光受体phyB会抑制TB1的表达,在麻风树(Jatrophacurcas)中赤霉素与细胞分裂素共同作用对TB1起着负调控的作用,促进侧芽的萌发[23];在拟南芥中SPA被认定为红光和远红光受体光敏色素信号传导过程中的下游负调控因子,光照强度或红光、远红光降低时,植物的分枝会受到抑制[24];秦琳琳等[25]发现,白桦(Betulaplatyphylla)中ZFP4基因含有多个脱落酸响应顺式作用元件,通过调控脱落酸信号途径来调控植物的生长;在脱落酸信号通路中,PYL1和ABF2基因起负调节作用,具体来说,它们可以抑制脱落酸受体和脱落酸应答转录因子的活性,从而影响脱落酸信号通路的整个过程[26]。这一机制使得PYL1和ABF2基因能够影响植物的开散程度和直立度。在北沙柳中,水培群体预测值与训练群体实验结果的趋势一致,有一定的线性关系,初步预测这些基因在北沙柳中同样可以调控植物的生长发育。
3.4 FHY1、ATX1、RFK1与株型的关系
FHY1、ATX1、RFK1是与北沙柳株型相关性高的基因。在训练群体实验结果中ATX1相关性最高(0.595 1),RFK1相关性最低(0.229 3),相关系数在0.2~0.6为中等程度的相关。FHY转录因子现已证明是远红光信号通路的重要组成部分,在拟南芥中,FHY转录因子在调节开花时间、枝条分生组织的发育等重要生命过程中都具有非常重要的功能[27]。通过蛋白互作网络分析可知,FHY1可能通过与光敏色素的相互作用来影响北沙柳的株型,也可能在生长素或赤霉素的影响下调节北沙柳的株型;铜是植物生长发育所需的微量元素,ATX1为铜伴侣蛋白,其作用就是供应植物生长所需的金属离子,将过量的金属离子排出从而达到解除重金属毒害的作用[15]。通过蛋白互作网络分析可知,ATX1可能通过转运铜离子调节铜的浓度来影响北沙柳的生长发育,也可能通过调节乙烯信号来影响北沙柳的生长发育;RFK1目前在北沙柳上未见报道,在本研究中腋芽部分的RFK1基因与北沙柳的株型相关性较高,显著高于茎尖,这说明RFK1基因作为北沙柳株型的后备基因作用是不可忽视的。
4 结论
按照“基因个数最少,相关系数最高”的原则,选择双基因组合ATX1+FHY1作为鉴别北沙柳株型的基因,该基因组合与冠高比的相关性较高,相关系数达0.740。基因组合法是通过基因与北沙柳株型的相关性构建起来的,存在误差,因此不能完全依靠基因组合的相关系数作为北沙柳株型的预测指标。如果想更准确地预测北沙柳的株型可适当地增加基因数量,提高基因组合与冠高比的相关性。
以不同株型北沙柳作为研究材料,从预测值与训练群体实验结果的相关性来看,利用转录组测序结果可以初步筛选北沙柳株型的后备基因。推测相关性越高,功能基因与目标性状的关系越密切。通过同一性状不同表现形式的个体与功能基因表达量的关系,可利用少数水培个体的转录组测序结果筛选目标性状的后备基因。这种方法不但去掉了复杂的转基因技术,加快了林木育种效率,同时还可以提高后备基因的选择精度。
