宁夏闽宁村西夏野利氏家族墓地人骨古DNA研究
2023-10-19常欣张红英张宇轩文少卿
□常欣 张红英 张宇轩 文少卿
一、引言
唐宋时期,党项在西北地区不断发展壮大并建立西夏[1]8。在正史史籍中,西夏史仅作为附传列于辽、金、宋等国史之后,内容简略,给学界研究西夏史带来了较大的困难。20 世纪以来,西夏文书不断出土问世,为西夏研究增添了新的史料,但这些史料涉及有关党项族属的记载很少。党项最早活跃于古析支之地,北周以后逐渐强盛。党项人以姓为部,“有细封氏、费听氏、往利氏、颇超氏、野辞氏、房当氏、米擒氏、拓跋氏,而拓跋氏最为强族”[2]459-461。目前对党项人群来源的争论多集中于党项大族拓跋氏,有“羌人说”[3]166-181[4]38-48、“鲜卑说”[5]1-2[6]409以及二者折中的“羌人—鲜卑混合说”[7]91-95。党项盛行火葬、天葬、水葬等 葬俗[8]17-22,已发掘的西夏墓葬中,出土的人骨遗骸十分稀少。发现的西夏墓葬主要有:宁夏银川西夏陵及陪葬墓、银川西夏区墓、永宁闽宁村墓、贺兰山拜寺口紫疙瘩墓、平罗涝湾村墓,甘肃武威西郊林场墓、武威西郊墓、武威双人骨灰墓、武威西苑小区墓、武威西关墓、武威西郊响水河墓,内蒙古准格尔旗西夏壁画墓、乌海参知政事墓、额济纳旗黑水城塔墓,陕西榆林靖边党项墓等[9]105-115。其中,仅在宁夏银川西夏陵陪葬墓M182 及永宁闽宁村墓地共发现4 具西夏时期人骨遗骸。因此,闽宁村墓地留存的人骨材料非常宝贵。本研究主要是利用新兴的古DNA 技术对党项野利氏家族的族属问题进行探讨。
以古DNA 技术为核心研究方法的分子考古是近年来兴起的、探索历史人群族源的重要手段。其方法是通过对古DNA 提取,获取其遗传信息,进而解读其中蕴含的人群形成史过程。分子考古的研究结果可与历史学、文献学、考古学、体质人类学研究结果相互印证,互为补充。
在分子考古研究中,Y 染色体和线粒体是重要信息载体。Y 染色体为父系遗传模式,线粒体为母系遗传,并在遗传过程中不会发生重组,二者结合可以帮助我们追溯个体父系和母系的历史[10][11]485-497。目前,通过父系Y 染色体和母系线粒体复原西北地区的人群迁徙与融合史的研究案例已经比较丰富。如付巧妹等对新疆41 个考古遗址的237 个个体线粒体基因组进行测序研究,揭示了青铜时代至历史时期,新疆人群的基因结构复杂但具有遗传上的连续性,显示了草原游牧人群、东北亚人群等从青铜时代便进入新疆地区与本地人群混合,并在新疆内部形成地区间的差异:新疆东部与北部的人群与东北亚人群具有更高的遗传相似性,而南部的人群与中亚人群更为接近。复旦大学的研究团队对甘肃地区汉代黑水国遗址及敦煌魏晋时期佛爷庙遗址的人骨Y染色体和线粒体进行研究[12][13],结合历史学、考古学及同位素进行考察显示,佛爷庙遗址和黑水国遗址的人群父系历史是黄河流域人群和本地人群的混合。佛爷庙遗址的人群父系来源还包括一些阿尔泰语人群、北欧亚人群以及东亚南方人的祖先成分。此外,结合线粒体信息表明,黑水国地区存在以男性为主导的从中原地区而来的长距离人口迁徙。宁夏地区的古DNA 研究早在2010 年便已开展,相关机构对彭阳出土的新石器时期6 具人类遗骸的线粒体及Y染色体进行研究,结果表明,彭阳地区史前人类与古代匈奴等北方游牧民族有密切的遗传亲缘关系[14]215-218。但相关研究仅此一例,对于西夏时期人骨的古DNA探索尚属空白。
闽宁村党项野利氏家族墓M4 与M7 出土了人骨遗骸。M4 墓主为一个未成年个体,M7 为夫妇合葬墓。M7 随葬铜甲片并在地面树立碑亭(B1),显示M7 墓主生前在族内应有较高的地位[15]50。本研究团队采集M7 出土的两具人骨材料,进行古DNA 提取。通过分子人类学手段获取父系Y染色体及母系线粒体遗传信息并分析遗传成分类型,进而尝试探讨野利氏家族的族属与族际婚姻问题。
二、材料与研究方法
(一)样本采集
闽宁村西夏墓地[15]1-5共有14 座墓葬,均为阶梯式墓道的土洞单室墓,墓室多呈方形,平顶或穹窿顶。发掘的8 座墓葬中有5 座火葬墓与3 座土葬墓,M4、M7 有人骨与棺木。韩康信对出土人骨进行了体质人类学鉴定[16]297-313,认为M4 墓主为9~11 岁未成年个体,性别不明;M7 的两个墓主为45~55 岁的男性与40~50 岁的女性,应为夫妇合葬墓。从M1、M7、M14 附近碑亭出土的残碑推断,闽宁村墓地的时代应在西夏早期,M1、M14 墓主为党项野利氏,M7男墓主也出自党项。墓地的墓葬方向均朝南,分布极有规律,推测整个闽宁村墓地是一处与野利氏家族有关的党项人墓地。本次研究采集的两个个体出土于M7夫妇合葬墓。
(二)DNA提取
DNA 提取工作在复旦大学科技考古研究院古DNA 实验室进行,严格按照古DNA 研究的操作规范[17]3-6,对样本进行处理:去污染预处理、样品研磨、DNA提取采用柱式提取法。
(三)文库构建与测序
使用NEBNextDNA 超快速文库制备试剂盒对DNA 进行标准的Illumina 文库构建,对获得的文库采用Agilent 2100 Bioanalyzer 平台进行文库质检,然后采用Illumina HiSeq X10 平台进行双端(paired-end sequencing)测序,获取读长为150bp的测序数据(fastq)。
(四)数据分析
基于原始下机数据,采用EAGER 软件的古DNA 分析流程[18]进行分析。首先通过ClipAnd-Merge 软件去除接头、引物序列及序列拼接,并过滤低质量序列,然后用bwa 软件(参数:aln)[19]1754-1760将过滤后的序列与人类参考基因组hg19 版本进行比对(线粒体参考基因组替换为rCRS 序列),生成BAM(Binary Alignment Map)文件。采用bedtools 软件[20]841-842从上一步BAM 文件提取出成功比对到人类参考基因组hg19 版本的所有序列,再将这些序列采用bowtie2软件[21]以加尾的rCRS 作为参考序列重新比对,生成新的BAM。最后,采用samtools 软件[22]2078-2079和bcftools软件[23]2156-2158进行变异位点的识别(VCF),然后将保留纯合突变的突变位点生成变异位点文件(hsd)。对于线粒体基因组,采用Haplogrep2 软件[24]并结合IGV 软件[25]178-192人工校验,推断个体的线粒体单倍型。Y 染色体单倍群通过ISOGG(国际遗传谱系学会,http://isogg.org/)和Y-full(https://www.yfull.com/tree/)数据库中位点对应关系,并在碱基和映射质量超过30 的情况下进行分析。单倍群测定使用Yleaf 软件中的脚本Yleaf.py进行[26]1291-1294,该软件输出了Y 染色体树分支路径上游的祖先型和衍生snp 的等位基因计数。最后采用IGV 软件[25]178-192人工校验。通过鉴定古代线粒体基因组中私有位点的杂合度来判断来自于外源DNA的污染率。综合古DNA片段长度、损伤模式以及污染率来判读DNA的真实性。
三、结果
(一)Y 染色体和线粒体DNA 序列变异情况及其单倍群归属
对样本进行测序后,通过分析比对研究对象的基因组序列占比,两个样本的DNA 内源性比例分别为36.89%与0.25%(表一),绝大部分外源DNA 为细菌和真菌,符合古DNA 高度降解和菌源污染的特点。DNA 片段末端5’端C->T突变特征代表了古DNA 的降解,M7 两个样本的损伤频率分别为5.25%和9.11%,符合古DNA 的损伤特征。同时基于线粒体评估了两个样本现代人类污染,污染水平低(<1%)。

表一 样本文库测序结果
通过分析已覆盖的线粒体及Y 染色体区域,成功读取了样本的突变位点信息,确定M7男性个体甲的线粒体所属单倍群为F1g,Y染色体所属单倍群为R1b1a1b-CTS1415^^;M7 女性个体乙个体的线粒体所属单倍群为F1b1c(表二)。

表二 样本的线粒体及Y染色体突变位点及单倍群归属
(二)M7甲的母系与父系遗传类型及其分布
M7 甲的线粒体单倍群为F1g,Y 染色体SNP位点及所属单倍群为R1b1a1b-CTS1415^^。整合目前已发表中国境内及相关人群线粒体单倍群F1g,使用ArcGIS 软件(ver.10.8.1)绘制了单倍群地理分布图(图一)。所参考的人群为40 个现代族群,共计22322个个体,覆盖了中亚、东亚、东南亚地区,包括中国汉族[27]1643-1652、藏族[28]、柯尔克孜 族[29]124-136,泰国本土人群[30]898-911和傈僳族[31]1563-1579,巴基斯坦哈扎拉人[32]、霍族[33]1008-1018,印度帕西族[34]等。图中还标记了归属于相同线粒体单倍群的6 个古代样本,分布于蒙古国及我国青海、陕西一带。从图一中可以看到,线粒体单倍群F1g在我国青藏地区及西南亚藏缅人群中广泛分布。以西藏定日藏族人群中的分布最为高频(15.79%),并向四周地区逐步递减。
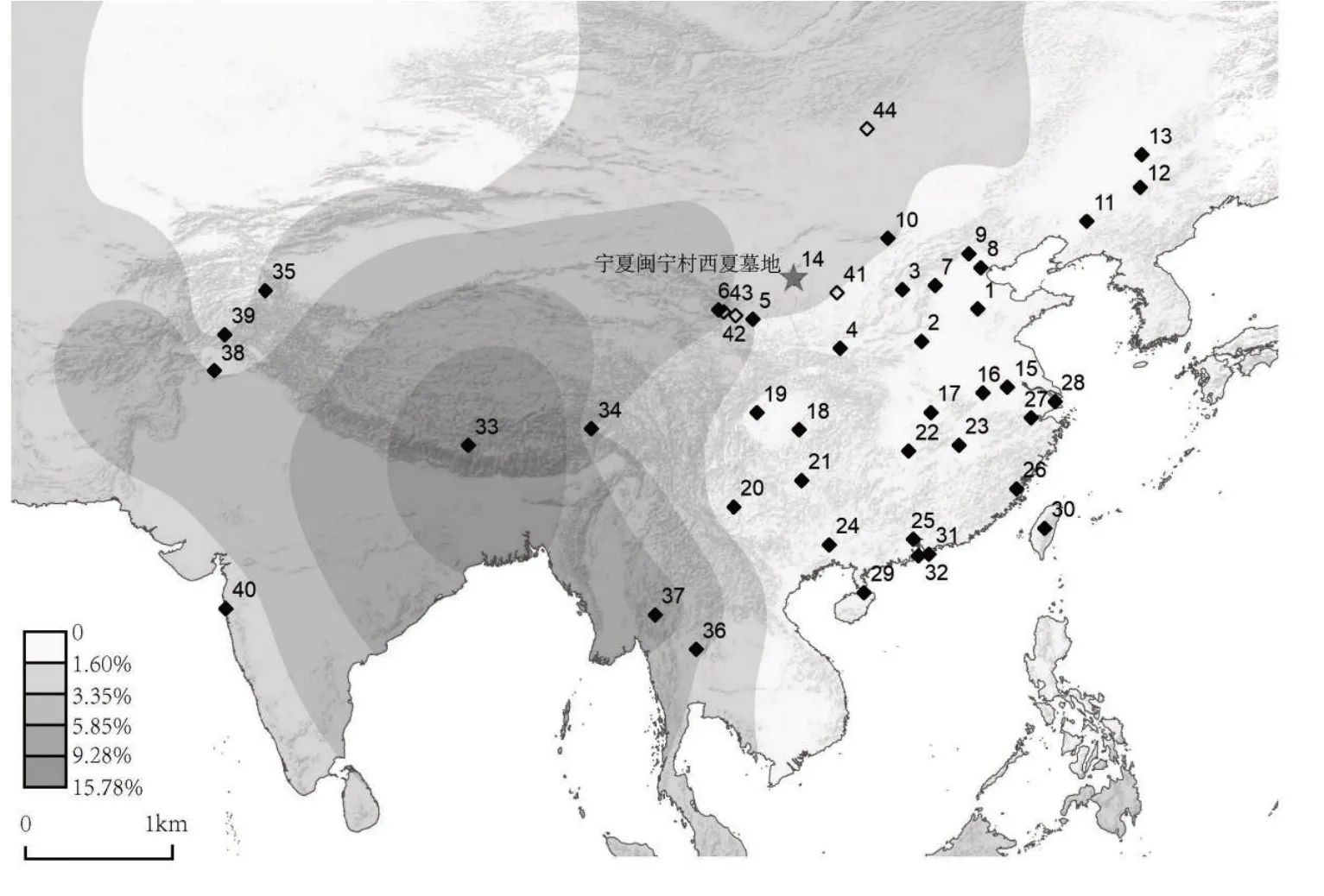
图一 M7甲所属线粒体单倍群F1g分布图
中国境内已发表的、属于线粒体单倍群F1g的古代个体分布于陕西榆林靖边新石器时代五庄果墚遗址(距今5106—4902 年前)[35]413-419、青海民和齐家文化喇家遗址(距今4000—3800 年前)[36]、青海平安东汉—北魏大槽子墓地(距今约2000 年前)[36]。此外,在蒙古国东北部肯特省(Khentii Aimag)发掘出土的中世纪晚期蒙古时代墓地Chandgan Talyn Uurkhai(距今744—582 年前)[37]890-904中也有个体的线粒体类型归属于F1g。
现代人参考群体:1.山东汉族,2.河南汉族,3.山西汉族,4.陕西汉族,5.甘肃汉族,6.青海汉族,7.河北汉族,8.天津汉族,9.北京汉族,10.内蒙古汉族,11.辽宁汉族,12.吉林汉族,13.黑龙江汉族,14.宁夏汉族,15.江苏汉族,16.安徽汉族,17.湖北汉族,18.重庆汉族,19.四川汉族,20.云南汉族,21.贵州汉族,22.湖南汉族,23.江西汉族,24.广西汉族,25.广东汉族,26.福建汉族,27.浙江汉族,28.上海汉族,29.海南汉族,30.台湾汉族,31.香港汉族,32.澳门汉族,33.西藏定日藏族,34.西藏林芝珞巴族,35.新疆塔什库尔干柯尔克孜族,36.泰国傣族,37.泰国傈僳族,38.巴基斯坦哈扎拉人,39.巴基斯坦霍族,40.印度帕西族。
古代人参考群体:41.陕西靖边五庄果墚遗址,42.青海民和喇家遗址,43.青海平安大槽子墓地,44.Chandgan Talyn Uurkhai。
以isogg2019—2020[38]725-736为参考,选取与R1b1a1b-CTS1415^^等价的位点R1b1a1b-M269,整合目前已发表的中国境内及附近地区人群Y染色体单倍群数据,绘制单倍群地理分布图(图二)。16 个现代参考人群,共计4817 个个体,覆盖了东欧、中亚、东亚和东南亚地区,包括伊比利亚半岛[39]、土耳其[40]、黎巴嫩[41]873-882、蒙古[42]121-126、乌兹别克斯坦[43]627-640、塔吉克斯坦[43]、巴基斯坦[44]202-221、中国新疆[45]1892-1899和云南[46]341-347地区、印度[44]202-221、老挝[47]1490-1506、泰国[47]1490-1506等。图中还标记出携带相同遗传标记的19 个古代个体,分布于俄罗斯、印度、埃及、哈萨克斯坦的考古遗址。
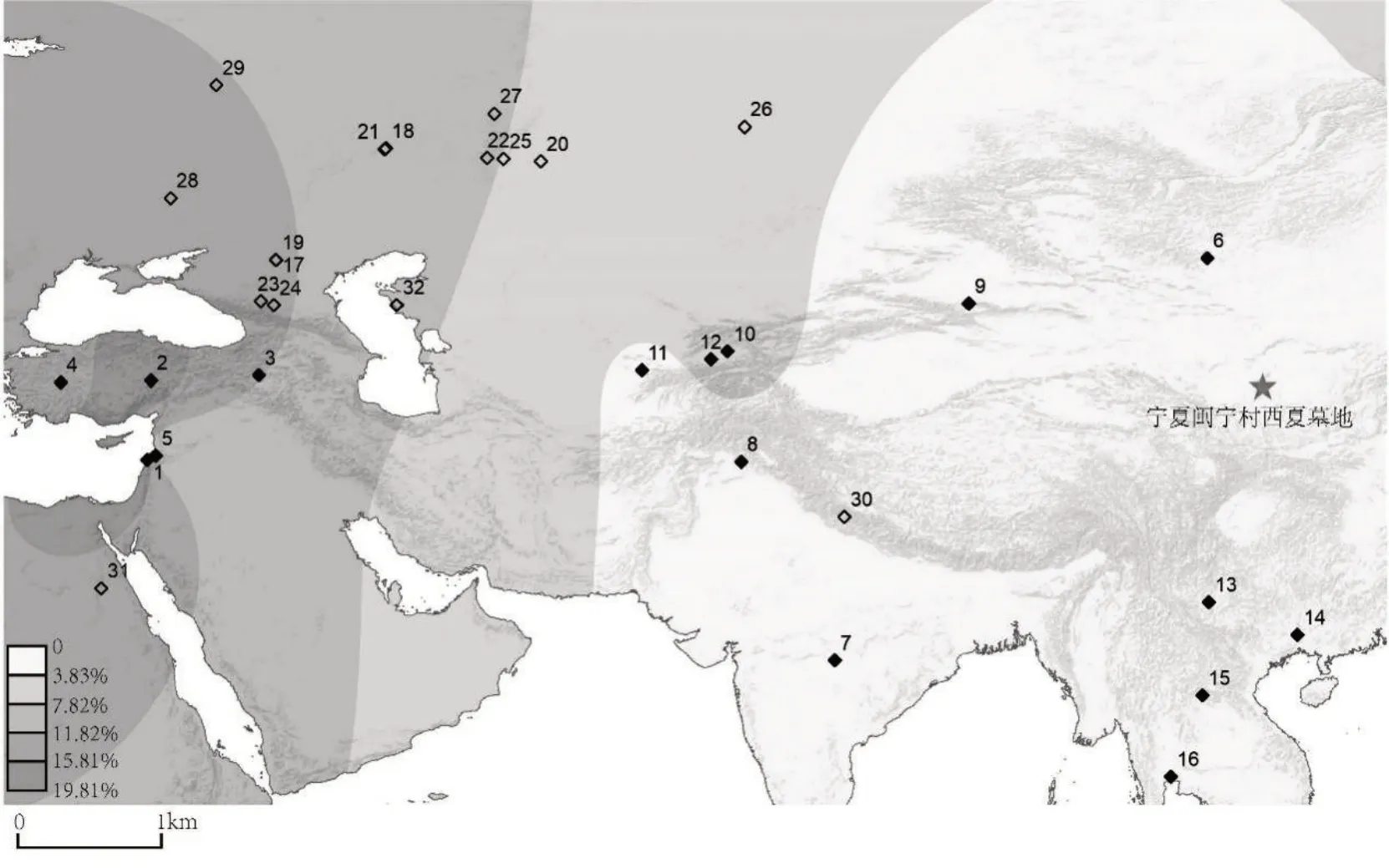
图二 M7甲Y染色体单倍群R1b1a1b的现代及古代人群分布
Y 染色体R1b1a1b 常见于欧洲地区,在安纳托利亚和高加索地区、俄罗斯部分地区以及中亚和南亚也很常见,其分布频率在欧亚大陆上由西向东逐步降低,在我国新疆少数民族柯尔克孜人群中有少量分布(0.96%)。古代人群携带这一父系遗传标记的人群主要分布于欧亚大陆草原地区今俄罗斯境内,如俄罗斯颜那亚文化的遗址:Kalmykia Temrta Ⅳ(距今约4850 年前)[48]167-172、Kalmykia Peshany Ⅴ(距今约4312年前)[48]167-172、萨马拉索克河流域Lopatino I(距今约4420±30 年前)[49]207-211,以及俄罗斯境内乌拉尔山脉的辛塔什塔文化墓地——Kamennyi Ambar 5(距今约3825年前)[50]、萨马拉索克河流域波塔波夫卡文化遗址——Grachevka(距今约3640±20 年前)[50]、梅绍夫斯卡亚文化遗址——Kapova cave(距今约3000年前)[48]、萨尔马提亚文化遗址——Temyasovo-1(距今2176 年前)[51]53-70、萨尔加特文化遗址——Mountain Bitiya#228/27(距今约2150 年前)[52]、纳瓦利诺文化二期遗址——Bartym16/B(距今约1441 年前)[53]、东欧大草原的萨尔托沃—玛雅克文化遗址——Belgorod(距今约1150 年前)[54]369-374、斯雅罗斯拉夫尔East Slav 遗址(距今约700 年前)[55]。此外活跃于丝绸之路的斯基泰人中也发现了该单倍群的个体,如位于俄罗斯的两个遗址[56]:Klin-Yar-3(距今约2700年前)、Zayukovo-3(距今约2450 年前),以及哈萨克斯坦Aigyrly sanctuary 遗址出土的4 号人骨(距今约1814 年前)[52]。另外,在距今约2450 年前的埃及托勒密王朝时期Thebes 木乃伊遗骸[57]524-531和距今150 年前印度喜马拉雅山区路普康湖(Roopkund Lake)出土遗骸[58]中也有零星发现。
现代人参考群体:1.伊比利亚半岛犹太人,2.土耳其中部,3.土耳其东部,4.土耳其西部,5.黎巴嫩,6.蒙古,7.印度,8.巴基斯坦,9.中国新疆柯尔克孜族,10.乌兹别克斯坦哈萨克族,11.乌兹别克斯坦塔吉克族,12.塔吉克斯坦塔吉克族,13.中国昆明回族,14.中国广西回族,15.老挝,16.泰国。
古代人参考群体:17.Kalmykia Temrta Ⅳ,18.Lopatino I,19.Kalmykia Peshany Ⅴ,20.Kamennyi Ambar 5,21.Grachevka,22.Kapova cave,23.Klin-Yar-3,24.Zayukovo-3,25.Temyasovo-1,26.Mountain Bitiya,27.Bartym,28.Belgorod,29.East Slav,30.Roopkund Lake,31.Thebes,32.Aigyrly sanctuary。
(三)M7乙的母系遗传类型及其分布
M7 乙的线粒体单倍群为F1b1c。整合目前已发表中国境内及相关人群线粒体单倍群F1b1c数据,绘制地理分布图(图三)。37 个现代参考人群,共计22772 个个体,集中分布于东亚地区,主要为中国各省区的汉族群体[29]124-136、内蒙古巴儿忽惕人[59]47-62、越南拉基族[60]、俄罗斯西伯利亚的喀木尼甘人[59]47-62和布里亚特人[61]71-81等。图中还标示了携带该遗传标记的12 个古代个体,覆盖了欧亚大陆北部区域,如俄罗斯、蒙古、中国新疆、吉尔吉斯斯坦、哈萨克斯坦等地区。
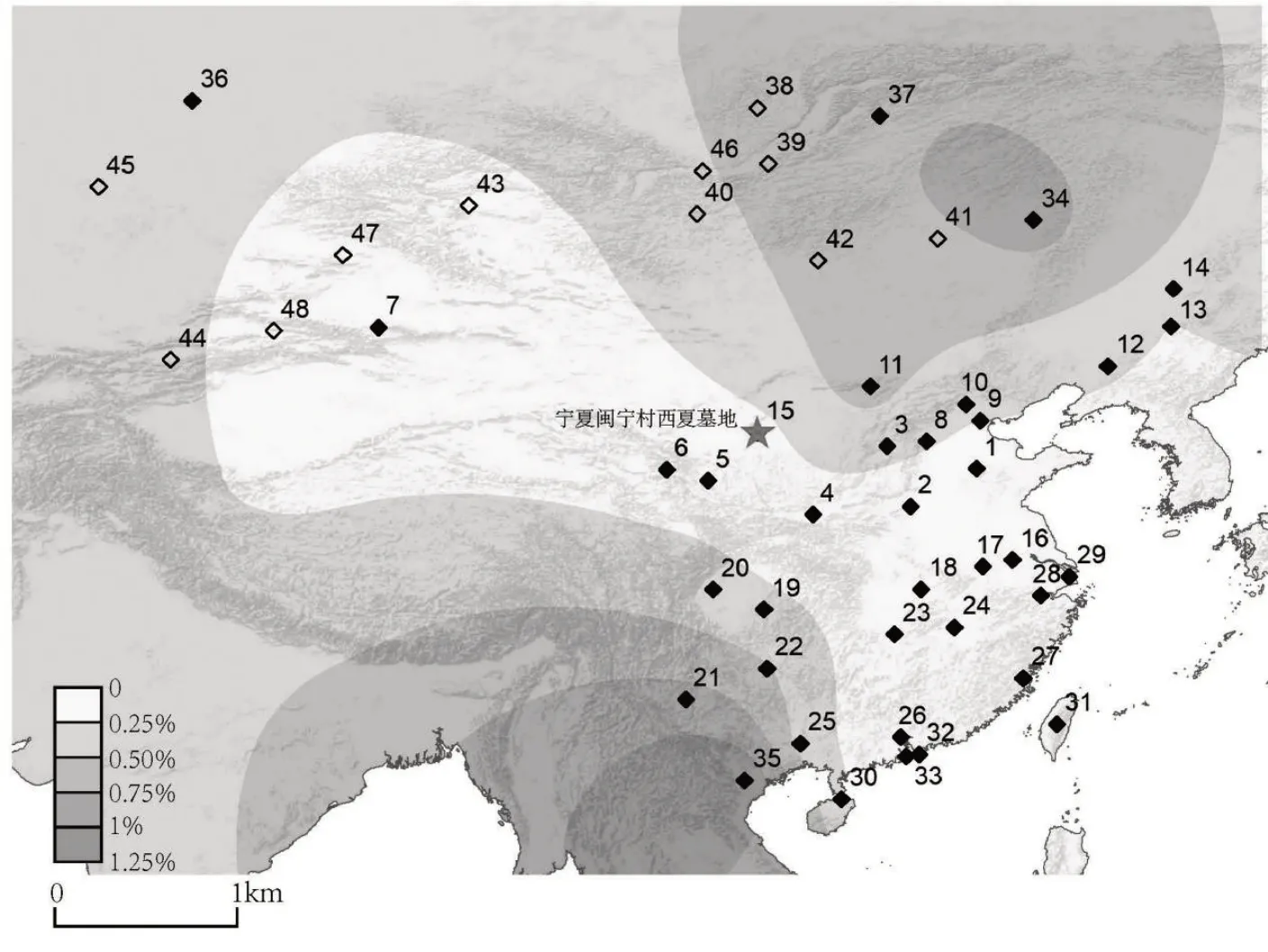
图三 M7乙所属线粒体单倍群F1b1c分布图
可以看到不同于M7 甲,M7 乙的线粒体单倍群主要分布于东北亚,南亚地区有着低频分布,而在中国西部宁夏、甘肃、陕西、四川、重庆地区也有零星分布,连接起东北亚与南亚地区。古代人群中携带线粒体单倍群F1b1c 的人群则主要集中在欧亚大陆北部,诸如俄罗斯西伯利亚地区勒拿河流域新石器时期Iushino 墓地(距今6612±23 年前)[62]1232-1245、俄罗斯西伯利亚ShamankaⅡGrave7(距今6329±33年前)[63]、俄罗斯布里亚特省贝加尔湖附近青铜时代早期Fofonovo 遗址(距今5731—5593年前)[37]890-904、蒙古北部布尔干省额金高尔河谷新石器时代墓地——(Marzyn Khutul,tomb 1-055-1,skeleton 2)(距今6615±25年前)[35]413-419、蒙古东方盟巴彦图门省新石器时期塔姆萨格布拉格文化遗存Choibalsan(距今6965±35 年前)[35]413-419、蒙古肯特省德勒格尔汗县铁器时代早期石板墓文化的Khanan 遗址(距今2875±20 年前)[35]413-419、蒙古乌布苏省铁器时代早期萨格勒文化的Ulaangom墓地(距今2210±20年前)[35]413-419、吉尔吉斯斯坦天山地区Kurgan Uch-Kurbu Grave#4 发现的匈奴人(距今1709±34 年前)[54]369-374哈萨克斯坦草原Osakarovskij 发现的斯基泰人(距今2489±39 年前)[54]369-374以及中国新疆吉木乃县青铜时代松树沟墓地(距今3336—3077 年前)[64]、新疆尼尔卡县铁器时代东买里遗址(距今2788—2239 年前)[64]。古DNA 数据主要分布在欧亚大陆北方,结合现代数据分布情况,暗示了携带线粒体单倍群F1b1c的人群向从东北亚沿着长城沿线向南扩散的历史。
现代人参考群体:1.山东汉族,2.河南汉族,3.山西汉族,4.陕西汉族,5.甘肃汉族,6.青海汉族,7.新疆汉族,8.河北汉族,9.天津汉族,10.北京汉族,11.内蒙古汉族,12.辽宁汉族,13.吉林汉族,14.黑龙江汉族,15.宁夏汉族,16.江苏汉族,17.安徽汉族,18.湖北汉族,19.重庆汉族,20.四川汉族,21.云南汉族,22.贵州汉族,23.湖南汉族,24.江西汉族,25.广西汉族,26.广东汉族,27.福建汉族,28.浙江汉族,29.上海汉族,30.海南汉族,31.台湾汉族,32.香港汉族,33.澳门汉族,34.内蒙古巴儿忽惕人,35.越南拉基族,36.俄罗斯西伯利亚喀木尼甘人,37.俄罗斯布里亚特。
古代人参考群体:38.Iushino,39.Fofonovo,40.Marzyn Khutul,41.Choibalsan,42.Khanan,43.Ulaangom,44.Kurgan Uch-Kurbu,45.Osakarovskij,46.ShamankaⅡ,47.新疆松树沟墓地,48.新疆东买里遗址。
四、讨论
(一)M7男性个体的父系和母系来源探讨
羌人是生活在我国西北的古老民族,先秦时期主要居住在黄河、湟水流域[65]835,秦汉以后羌人活动范围渐渐扩大。魏晋时期羌人的活动范围在秦州、永州、凉州、益州,以及今天黄南、甘南藏族自治州区域[66]278。十六国时期,羌人先后建立了后秦、宕昌羌、邓至羌等政权。后秦强大时,版图一度西到河西、东括兖徐[67]699。青海地区青铜时代的卡约文化、辛店文化被认为是羌人创造的文化,与中原文化有着截然不同的面貌[68]117-152[69]51-62。青海地区青铜时代齐家文化磨沟遗址[70]258-268与青海汉代陶家寨遗址[71]239先民都被认为是氐羌人群。韩康信和张君认为,卡约文化居民尤其是李家山组与东部藏族类型存在较为直接的种族起源联系[72]6-15[73]381-415。赵永生认为,青海磨沟遗址古代居民与蒙古人种中的东亚蒙古人种关系最为接近,且磨沟齐家组在“古西北类型”中处于承上启下的位置,其体质特征与甘青地区新石器时代居民基本一致[74]。因此,前期的体质人类学研究揭示甘青地区的古代人类在新石器时代,具有比较一致的体质类型。到了青铜时代,虽然这种体质类型的人群还广有分布,但由于人群的流动,甘青地区古代人群原本单一的体质特征出现分化。但是整体体质特征上较为一致,与氐羌人群存在关联。
线粒体单倍群F1g 在古代遗址中主要分布于陕西与青海地区。五庄果墚遗址、喇家遗址和大槽子墓地位于陕西、青海地区,从遗址出土人群体质特征与DNA 遗传信息上来看,这些人群与黄河上游人群,特别是氐羌人群更为接近。前期古DNA 研究也支持这一结果。对于五庄果墚人群的基因组分析表明该人群和现在的汉族、羌族、藏族个体关系最为密切[35]413-419。现代线粒体单倍群F1g 也高频出现在相关地区人群,如西藏定日藏族(15.78%)、西藏林芝珞巴族(3.26%)、青海汉族(4%)和宁夏汉族(2.56%)等。党项最早出现于古析支之地,即今黄河上游青海境内,党项也被认为来源于氐羌人群[75]1422。线粒体单倍群F在磨沟人群[70]258-268和陶家寨人群[71]239先民中分布频率较高,分别为8.7%和17.2%。线粒体单倍群F 的下游支系F1 在现代羌语支人群中占比最高(5.54%)[76],且从图一中可见F1 下游支系F1g 目前主要分布于藏缅语人群中。因此可以推断拥有线粒体单倍群F1g 的M7 男性个体的母系来源与羌藏人群有着极为密切的关系。
M7 男性个体的父系来源则与欧亚大陆西部草原游牧人群更为接近。欧亚草原地区青铜时代早期颜那亚文化中发现较多例携带Y 染色体R1b1a1b 的个体,颜那亚文化的人群遗传成分在新疆铁器时代早期石人子沟遗址中发现,西欧亚人群的遗传成分随着人群流动而东进[77]。新疆东部哈密天山北路墓地的古DNA 研究发现青铜时代早期(距今3600 年前)该地区由甘青地区氐羌遗传成分的古代人群迁入,并与具有草原西部遗传成分的人群混合[78]71-80。历史时期欧亚大陆上的人群纷繁复杂,丝绸之路与河西走廊是重要的交通往来要道,草原游牧人群与中国西北地区人群往来交流密切。同样的父系成分也发现于斯基泰人中。汉文典籍中,斯基泰人被称为“塞人”。塞人自西向东迁徙,先秦至汉初主要分布在中亚锡尔河以北一带,《汉书•西域传》记载:“(乌孙国)东与匈奴、西北与康居、西与大宛、南与城郭诸国相接。本塞地也,大月氏西破走塞王,塞王南越县度,大月氏居其地。后乌孙昆莫击破大月氏,大月氏徙西臣大夏,而乌孙昆莫居之,故乌孙民有塞种、大月氏种云。”[79]785公元前2 世纪,塞人、月氏都在河西走廊敦煌、祁连山一带进行游牧活动。《后汉书•西羌传》载:“湟中月氏胡,其先大月氏之别也,旧在张掖、酒泉地。月氏王为匈奴冒顿所杀,余种分散,西逾葱岭。其羸弱者南入山阻,依诸羌居止,遂与共婚姻。及骠骑将军霍去病破匈奴,取西河地,开湟中,于是月氏来降,与汉人错居。”[65]847塞人、月氏与氐羌人群在西北地区长时段的活动中,文化交流与民族融合不断上演,党项野利氏的父系R1b1a1b可能与上述欧亚大陆西部草原游牧人群东迁有关。
(二)M7女性个体的母系来源探讨
现代、古代人群线粒体单倍群F1b1c 的分布主要与东北亚人群有关。韩康信先生体质人类学鉴定结果也表明M7 女性个体与东北亚地区人群关系较为密切[15]157-183。党项不同宗族部落间以及与周边民族间保持着密切而频繁的婚姻关系。M7女性个体可能来自于党项内部其他姓氏部族,也可能来自于与外族人群的联姻。
党项内部姓氏众多,人群来源多样,如党项拓跋氏的族源有鲜卑拓跋说。拓跋氏与野利氏也多有通婚,李继迁之母即为野利氏,李元昊也迎娶野利氏女子为皇后。陕北横山《故野利氏夫人墓志铭》也记载了党项野利氏女性与拓跋氏的婚姻[80]24-31。M7 女性个体可能来自党项拓跋氏或党项内部其他有北方民族来源的姓氏部落。
M7 也有可能来自与党项杂居的吐谷浑人群或其他北方民族。吐谷浑本是人名,是鲜卑部慕容涉归庶子,率部西迁定居。《梁书》卷五十四《诸夷列传•西北戎传》载:“河南王者,其先出自鲜卑慕容氏。初,慕容奕洛干有二子,庶长曰吐谷浑,嫡曰廆。洛干卒,廆嗣位,吐谷浑避之西徙。……遂西上陇,度枹罕,出凉州西南,至赤水而居之。其地则张掖之南,陇西之西,在河之南,故以为号。其界东至垒川,西邻于阗,北接高昌,东北通秦岭,方千余里,盖古之流沙地焉。”[81]810这一区域也是当地土著民族氐羌部落的聚居区,M7女性个体可能由此而来。
此外,西夏前后三次与契丹和亲。《宋史•夏国传》载:继迁请婚契丹,“辽以义成公主嫁继迁,册为夏国主”[2]9617。《辽史•圣宗纪》载:辽统和四年(986),李继迁“引五百骑款寨,愿婚大国,永作藩辅”。辽圣宗当即下诏令:“以王子帐节度使耶律襄之女汀,封义成公主下嫁,赐马三千匹。”辽统和七年(989)三月,成婚[82]359。宋天圣七年、辽太平九年(1029)春二月,德明欲结好契丹,便为元昊请婚,天圣八年十二月,契丹以兴平公主嫁元昊。至夏国主乾顺时代,为结亲于辽,曾屡次向辽主请婚,辽主封宗室女南仙为成安公主,下嫁乾顺。[83]42-48拓跋氏在统治西夏时期,注重利用党项大族的势力,并在政治、军事上遇到困难时以和亲的手段积极向外寻求援助或缓和与敌对势力之间的矛盾。在这样的背景下,M7女性个体也有可能从东北亚地区来到西夏与党项大族野利氏通婚。
五、结论
本文通过对宁夏闽宁村西夏野利氏墓地M7夫妇合葬墓的两个个体进行古DNA 分析,讨论了M7 男性个体父系Y 染色体与母系线粒体来源及女性个体的母系线粒体来源。
M7 男性个体的Y 染色体单倍群为R1b1a1b-CTS1415^^,线粒体单倍群为F1g。通过分析现代及古代人群相关遗传类型的分布,我们认为M7男性个体父系来源则更多地接近欧亚草原游牧人群,而母系来源与氐羌人群有关。从DNA 上看,该个体的族属为氐羌人群与欧亚大陆西部草原人群的混血。M7 女性个体线粒体单倍群为F1b1c,与北方民族人群较为接近,可能来自党项内部与北方民族有关的部落,也可能来自族外人群如与党项混居的吐谷浑,或与西夏政治联姻的其他北方民族。
